
Abstract
The cutaneous wound healing process is tightly regulated by a range of cellular responses, including migration. Sphingosine-1-phosphate (S1P) is a signaling lipid produced in keratinocytes (KC) and it is known to stimulate skin wound repair through increased KC migration. Of the multifunctional triterpene ginsenosides, Rb1 enhances cutaneous wound healing process by increasing KC migration, but cellular mechanisms responsible for the Rb1-mediated increase in KC migration are largely unknown. Therefore, we hypothesized that, and assessed whether, Rb1 could stimulate KC migration through S1P-dependent mechanisms. Rb1 significantly increases S1P production by regulating the activity of metabolic conversion enzymes associated with S1P generation and degradation, sphingosine kinase 1 (SPHK1) and S1P lyase, respectively, in parallel with enhanced KC migration. However, blockade of ceramide to S1P metabolic conversion using a specific inhibitor of SPHK1 attenuated the expected Rb1-mediated increase in KC migration. Furthermore, a pan-S1P receptor inhibitor pertussis toxin significantly attenuated Rb1-induced stimulation of KC migration. Moreover, the Rb1-induced increases in KC migration required S1P receptor(s)-mediated activation of ERK1/2 and NF-κB, leading to production of key cutaneous migrating proteins, matrix metalloproteinase (MMP)-2 and MMP-9. Taken together, the results show that Rb1 stimulates KC migration through an S1P→S1P receptor(s)→ERK1/2→NF-κB→MMP-2/-9 pathway. This research revealed a previously unidentified cellular mechanism for Rb1 in enhancing KC migration and pointing to a new therapeutic approach to stimulate the cutaneous wound healing process.
Introduction
C
The skin produces a number of lipid modulators, including prostanoids and sphingolipids, which contribute to maintain overall skin integrity by their structural properties and signaling functions. 11 Recent studies demonstrated that sphingolipids also regulate would healing processes as well as restoration of epidermal barrier function during wound repair. 12 –14 In particular, a signaling sphingolipid metabolite sphingosine-1-phosphate (S1P) enhances the cutaneous wound healing process through increased expression of MMPs, which are regulated by intracellular kinases, for example, Akt, ERK1/2, PI3K, and certain transcriptional factors, including NF-κB, Sp1, And AP-1. 3,15 In contrast to migration, prior studies revealed that S1P significantly suppresses KC proliferation. 16 These results suggest that modulation of S1P by chemicals could enhance wound healing.
Ginsenosides, triterpenoid saponins found in Panax ginseng, have been shown to modulate diverse cellular/tissue metabolisms to improve disease conditions, such as diabetes, atherosclerosis, hyperlipidemia, hypertension, and multiple malignancy, due to their roles in immune modulation. 17 –22 While over 30 ginsenoside molecular species have been identified, certain species of ginsenoside alter skin function, including stimulation of collagen production leading to potential antiaging agents and wound healing effects. However, mechanisms responsible for these putative biological effects of ginsenosides are largely unknown.
Because prior studies have shown that ginsenosides regulate sphingolipid metabolism, 23 we here investigated an involvement of ginsenoside-mediated alteration of sphingolipid in cutaneous wound healing. We found that Rb1, a ginsenoside species, significantly increases cellular levels of S1P signals to activate ERK1/2 and NF-κB, leading to enhanced KC migration through increased expression of MMP-2 and MMP-9. Moreover, these studies indicate that Rb1-mediated increases in S1P enhance KC migration through an S1P receptor-dependent mechanism. The insights gained from these studies illuminate a previously unidentified cellular mechanism for this ginsenoside in regulating KC migration.
Materials and Methods
Cell culture
Immortalized, nontransformed (HaCaT) human KC, derived from normal human skin (a gift from N. Fusenig, Heidelberg, Germany), were grown as described previously. 24 Culture medium was switched to serum-free KC growth medium containing 0.07 mM calcium chloride and growth supplements (Thermo Fisher Scientific, Carlsbad, CA, USA) 1 day before treatment with ginsenosides.
Cell migration assay
In vitro wound scratch assay was performed as described previously.
25
Briefly, cells were grown to 80% confluence in 12-well plates. To eliminate cell proliferation, cells were pretreated with 10 μg/mL mitomycin C (Sigma-Aldrich, St. Louis, MO, USA) for 1 h and washed with phosphate-buffered saline before scratching. The scratching was performed by scraping with a sterile rubber policeman, followed by incubation with media only (Vehicle) or with 20 or 50 μg/mL of ginsenosides dissolved in culture media for 48 h. The wound closures were subsequently photographed under a microscope (Eclipse Ti-U; Nikon Corporation, Tokyo, Japan) using a QImaging Retiga 2000R CCD digital camera (QImaging, Surrey, Canada). Cell migration was analyzed by image J (image processing and analysis in Java by NIH image:
Western immunoblot analysis
Western blot analysis was performed as described previously. 24 Cell lysates, prepared in radioimmunoprecipitation assay buffer, were resolved by electrophoresis on 4–12% Bis-Tris Gel (Thermo Fisher Scientific). Nuclear and cytoplasmic fractions were prepared using extraction reagents (Thermo Fisher Scientific). Resultant bands were blotted onto nitrocellulose membranes, probed with anti-MMP-2, anti-MMP-9, anti-ERK, anti-phosph ERK (Thr202/Tyr204), anti-NF-κB p65, or anti-human β-actin (Abcam, Cambridge, MA, USA), and detected using enhanced chemiluminescence (Thermo Fisher Scientific).
Measurement of contents of sphingolipids
The levels of ceramide, sphingosine, and S1P were quantified using a high-performance liquid chromatography (HPLC) system equipped with a fluorometrical detector system (JASCO, Tokyo, Japan), as described previously. 24,26 Sphingolipids are expressed as pmol/mg protein.
Enzyme activity assays for sphingomyelinases
The activity of acidic and neutral sphingomyelinase (SMase) was measured by BODIPY-SM substrate using HPLC-fluorescence detector (FLD) assay system, as described previously. 27 The activity of SMases is expressed as sphingosine pmol/mg protein/min.
Enzyme activity assays for sphingosine kinase 1 and SPHK2
Sphingosine kinase 1 (SPHK1) and SPHK2 activities were determined as described previously. 23,28 Briefly, cell lysates in 20 mM Tris (pH7.4) containing 5 mM EDTA, 5 mM EGTA, 3 mM β-mercaptoethanol, 5% glycerol, protease inhibitors (Sigma-Aldrich), and phosphatase inhibitors were incubated with 10 μL of 200 μM C17-Sphingosine as an SPHK substrate. To assay each isoform of SPHK activity, 0.5% Triton X-100 or 1 M KCl (for SPHK1 and SPHK2, respectively) was added into assay buffer and then incubated at 37°C for 30 min. Enzyme reactions were terminated by the addition of CHCl3:MeOH:HCl (8:4:3, v/v/v). C17-sphinganine-1-phosphate (100 pmol) was added as an internal standard. The organic phage separated by addition of CHCl3 was dried using a vacuum system (Vision, Freiburgstrasse, Switzerland). The dried residue was redissolved in MeOH and then analyzed by liquid chromatography–electrospray ionization tandem mass spectrometry (LC-ESI-MS/MS; API 3200 Triple quadruple mass; AB/SCIEX) as above, and the activities of SPHK1 and SPHK2 are expressed as C17-S1P pmol/mg protein/min.
Enzyme activity assay for S1P lyase
S1P lyase activity was assessed as described previously. 29 Briefly, cell lysates (50 μg) prepared in lysis buffer, as reported previously, 30 were incubated with 10 nmol S1P for 20 min. The reaction was stopped by lipid extraction with the addition of 100 pmol of (2E)-d5-hexadecenal as the internal standard. Total lipid extracts were derivatized with 5 mM semicarbazide hydrochloride in methanol containing 5% formic acid at 40°C for 2 h and analyzed by ESI-LC/MS/MS (API 3200 triple quadrupole mass; AB/SCIEX), as described previously. 30 The activity of S1P lyase is expressed as pentadecanal pmol/mg protein/min.
Statistical analyses
All experiments were repeated at least thrice. For each experiment, results from triplicate samples were expressed as the mean ± standard deviation. Significance between groups was determined using unpaired Student's t-tests. The P values were set at <.01.
Results
Rb1 increases cellular levels of S1P in parallel with enhanced migration of human KC
Although a number of different ginsenosides have been shown to promote cutaneous wound healing process through induction of cell migration, 31,32 the details of the regulatory mechanism remain unknown. Because prior studies have shown that S1P is a key signaling mediator to regulate KC migration, we hypothesized that a ginsenoside(s)-mediated increase in S1P enhances KC migration. To address this hypothesis, we first determined which structurally and functionally different ginsenosides could increase cellular S1P levels by measuring the activity of both S1P-producing and S1P-degrading enzymes, SPHK1 and S1P lyase, respectively. Of the 11 ginsenosides tested, Rb1 significantly increases SPHK1 activity, whereas the other ginsenosides did not alter or even decreased the activity (Table 1). In addition, S1P lyase activity was significantly suppressed in cells treated with certain ginsenosides, such as Rb1, Rc, Re, Rg1, and Ro (Table 1). Together these results suggest that Rb1 is a potential candidate that can increase cellular levels of S1P due to its ability to increase S1P-producing enzyme, SPHK1, and to inhibit S1P lyase, an enzyme that degrades S1P. Thus, we employed Rb1 in all of subsequent studies described below. Accordingly, as shown by lipid quantification analysis, there is an increase in not only intracellular S1P but also extracellular S1P levels in conditioned media from human KC treated with Rb1 (Table 2). Our lipid assays further identified that ceramide and sphingosine, precursor lipids of S1P, were elevated in cells following Rb1 treatment (Table 2). We next determined whether Rb1 stimulates KC migration. Wound scratch assay showed that wound coverage was significantly increased in cells treated with Rb1 as well as exogenous S1P, a known inducer of KC migration (Fig. 1A, B). Consistent with increased wound coverage, protein expressions of MMP-2 and MMP-9, critical biomarkers required for initiation/maintenance of KC migration and wound reepithelialization, were upregulated in response to Rb1 treatment (Fig. 1C). Together, these results indicate that Rb1 enhances KC migration, in parallel with an increase in cellular S1P levels and MMP-2/-9 expressions.
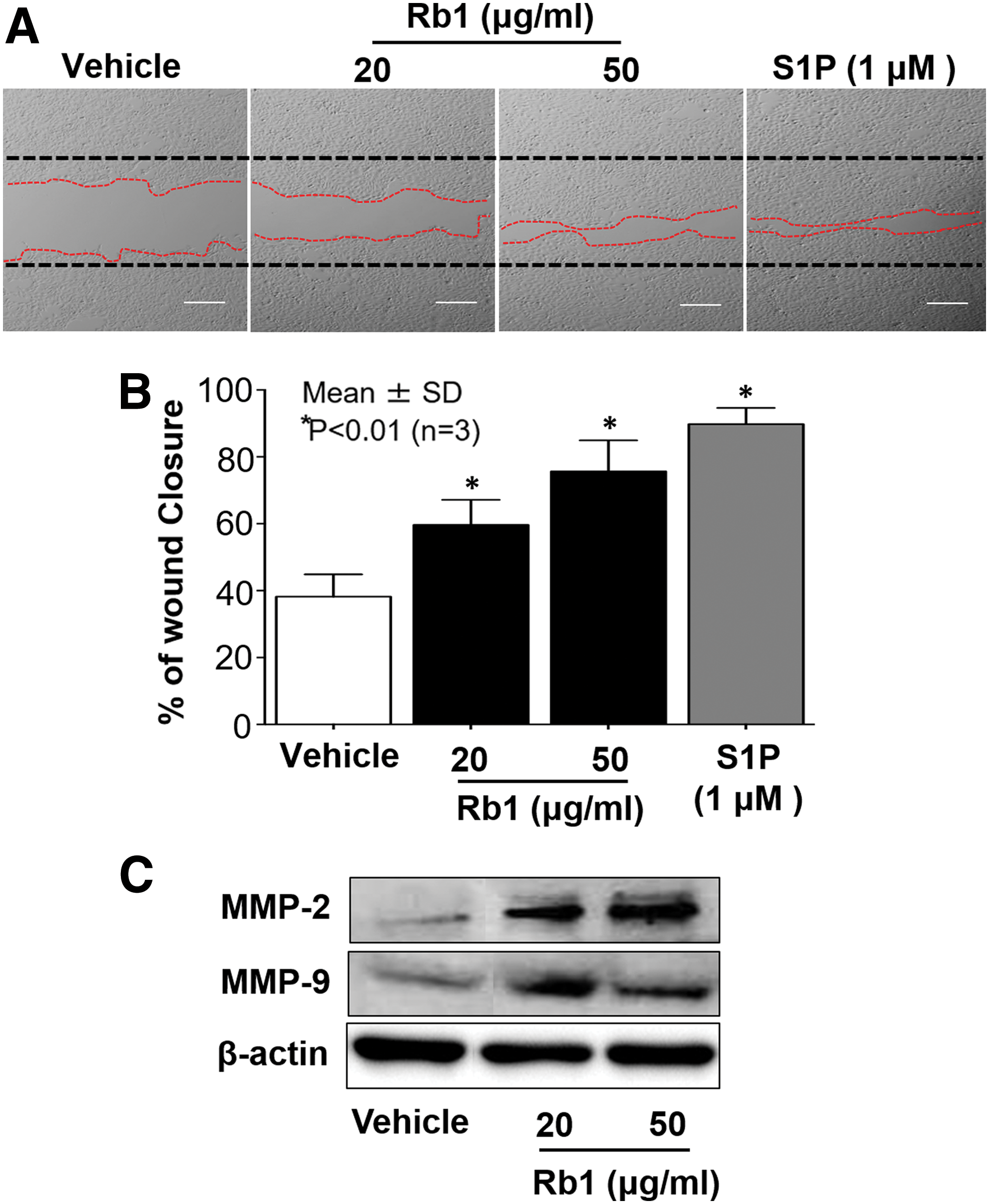
Rb1 stimulates KC migration in parallel with increased expression of MMP-2 and MMP-9. Human KC were grown to confluence and in vitro wound was produced using a sterile scraper, followed by incubation with Rb1 (20–50 μg/mL) or exogenous S1P (1 μM) for 48 h. Black dashed lines represented initial scratch and red dashed lines the migration front
Altered Activity of Sphingosine Kinase 1 and Sphingosine-1-Phosphate Lyase in Human Keratinocytes Following Ginsenosides
Human KC were treated with different ginsenosides (20 μM).
Values are mean ± SD (n = 3).
P < .01 versus vehicle control.
KC, keratinocytes; S1P, sphingosine-1-phosphate; SD, standard deviation; SPHK1, sphingosine kinase 1.
Increased Levels of Sphingolipids in Keratinocytes in Response to Rb1 Treatment
Human KC were treated with ginsenoside Rb1 (20–50 μM).
Values are mean ± SD (n = 3).
P < .01 versus vehicle control.
Rb1-mediated increase in S1P stimulates KC migration through S1P receptor-dependent mechanism
Because our results revealed that Rb1 increases cellular levels of ceramide, we next determined the mechanism by which cellular ceramide was elevated in cells treated with Rb1 by measuring the activity of SMase, an enzyme responsible for production of ceramide from sphingomyelin (SM), which is the most abundant lipid species localized at the plasma membrane. The activity assay showed a significant increase in both acidic and neutral SMase following Rb1 treatment (Table 3). We then assessed the involvement of enzymes involved in the conversion of ceramide to S1P. Rb1 significantly increases the activity of SPHK1, but not SPHK2, key enzymes in the production of S1P from ceramide (Table 4). Moreover, as described above, S1P lyase, the intracellular enzyme that catalyzes the irreversible degradation of S1P into phosphoethanolamine and (2E)-hexadecenal, was suppressed in KC following Rb1 treatment (Table 4). These results indicate that Rb1 increases S1P production through increased metabolic conversion of ceramide into S1P, as well as declined activity of S1P lyase, an enzyme required for S1P degradation. Next, we further determined whether S1P is responsible for Rb1-mediated increases in KC migration. Blockade of S1P production using SKI, a specific inhibitor of SPHK1, significantly attenuated the expected Rb1-induced increase in KC migration (Fig. 2). These results show that the Rb1-induced enhancement of KC migration can be attributed to increased S1P generation. Moreover, since S1P mainly regulates diverse biological functions, including migration, through the binding of ligands to their receptors, we next explored whether Rb1-mediated increases in S1P stimulates KC migration through S1P receptor-dependent mechanisms. Our wound scratch assay showed that a pan-S1P receptor inhibitor, pertussis toxin (Ptx), significantly suppressed Rb1-induced stimulation of KC migration, suggesting that Rb1-induced production of S1P enhances cell migration through a receptor-dependent mechanism (Fig. 3).
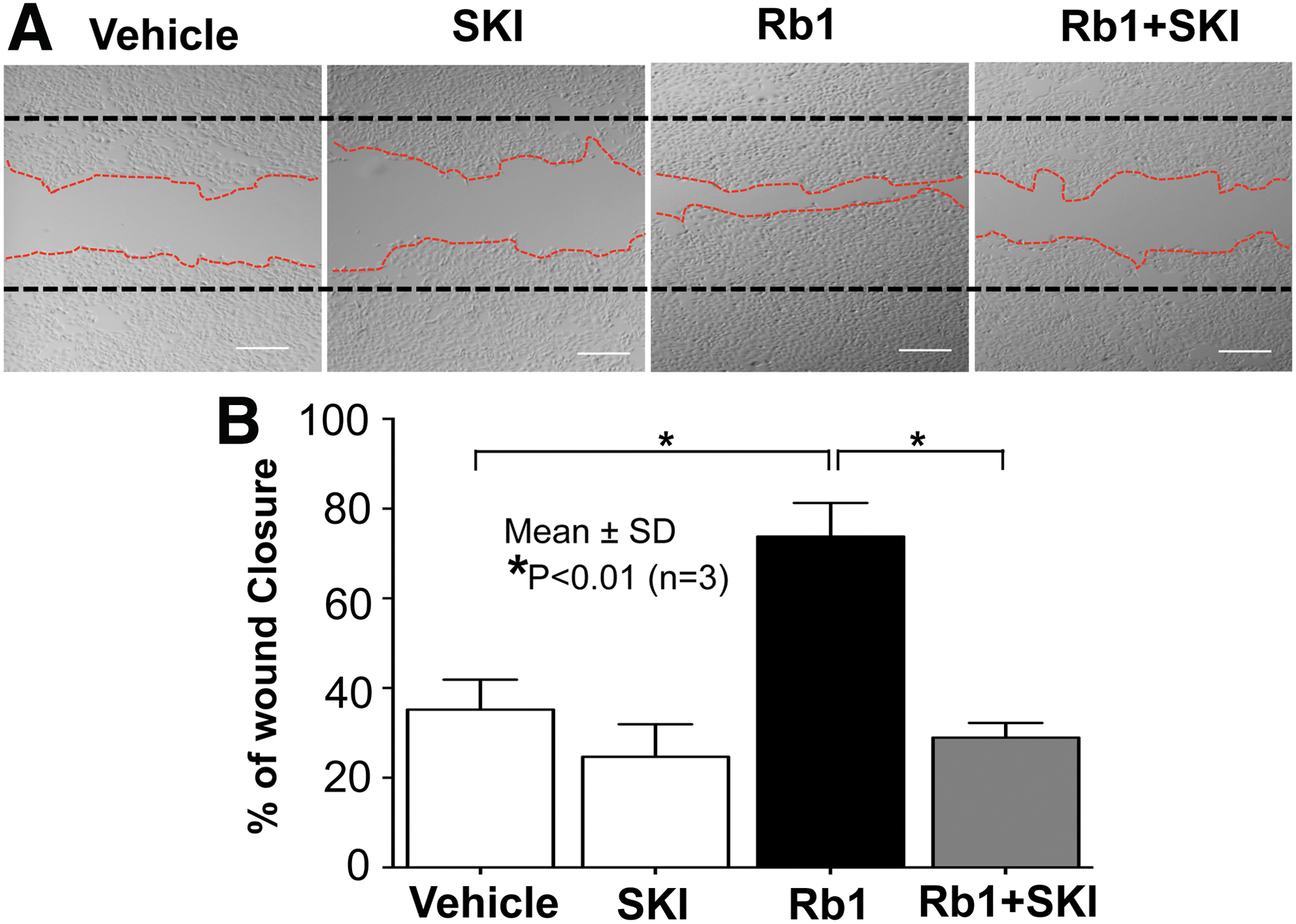
S1P is responsible for Rb1-mediated increase in KC migration. KC were grown to confluence and in vitro wound was produced using a sterile scraper. After wounding, KC were pretreated with a specific inhibitor of SPHK1, SKI (1 μM), followed by incubation with Rb1 (50 μg/mL). Black dashed lines represented initial scratch and red dashed lines the migration front

Rb1-induced increase in S1P enhances cell migration through receptor-dependent mechanism. KC were grown to confluence and in vitro wound was produced using a sterile scraper. After wounding, KC were pretreated with a pan inhibitor of S1P receptors, Ptx (12.5 ng/mL), followed by incubation with Rb1 (50 μg/mL). Black dashed lines represented initial scratch and red dashed lines the migration front
Increased Activity of Sphingomyelinases in Keratinocytes Exposed to Rb1 Treatment
Human KC were treated with ginsenoside Rb1 (20–50 μM).
Values are mean ± SD (n = 3).
P < .01 versus vehicle control.
SMases, sphingomyelinases.
Altered Activity of Sphingosine-1-Phosphate Generation and Degradation in Keratinocytes Following Rb1 Treatment
Human KC were treated with ginsenoside Rb1 (20–50 μM).
Values are mean ± SD (n = 3).
P < .01 versus vehicle control.
Rb1-induced stimulation of KC migration requires ERK1/2 and NF-κB activation
Prior studies demonstrated that MMP-2 and MMP-9 are a critical factor for cell migration and their expressions are signaled by activation of the ERK1/2-NF-κB axis. 33 Therefore, we next measured changes in ERK1/2 phosphorylation and NF-κB activation after Rb1 exposure. After incubation with Rb1, additional phosphorylated ERK1/2 levels were increased (Fig. 4A). In addition, NF-κB activation assessed by nuclear translocation was significantly was elevated following Rb1 treatment (Fig. 4B). Moreover, our inhibitor studies revealed that inhibition of NF-κB activation using a specific inhibitor of NF-κB, BAY-7082, attenuated the Rb1-mediated increase in KC migration (Fig. 4C, D). Together, these results indicate that the Rb1-induced increases in S1P activates ERK1/2 and NF-κB, leading to enhanced KC migration.
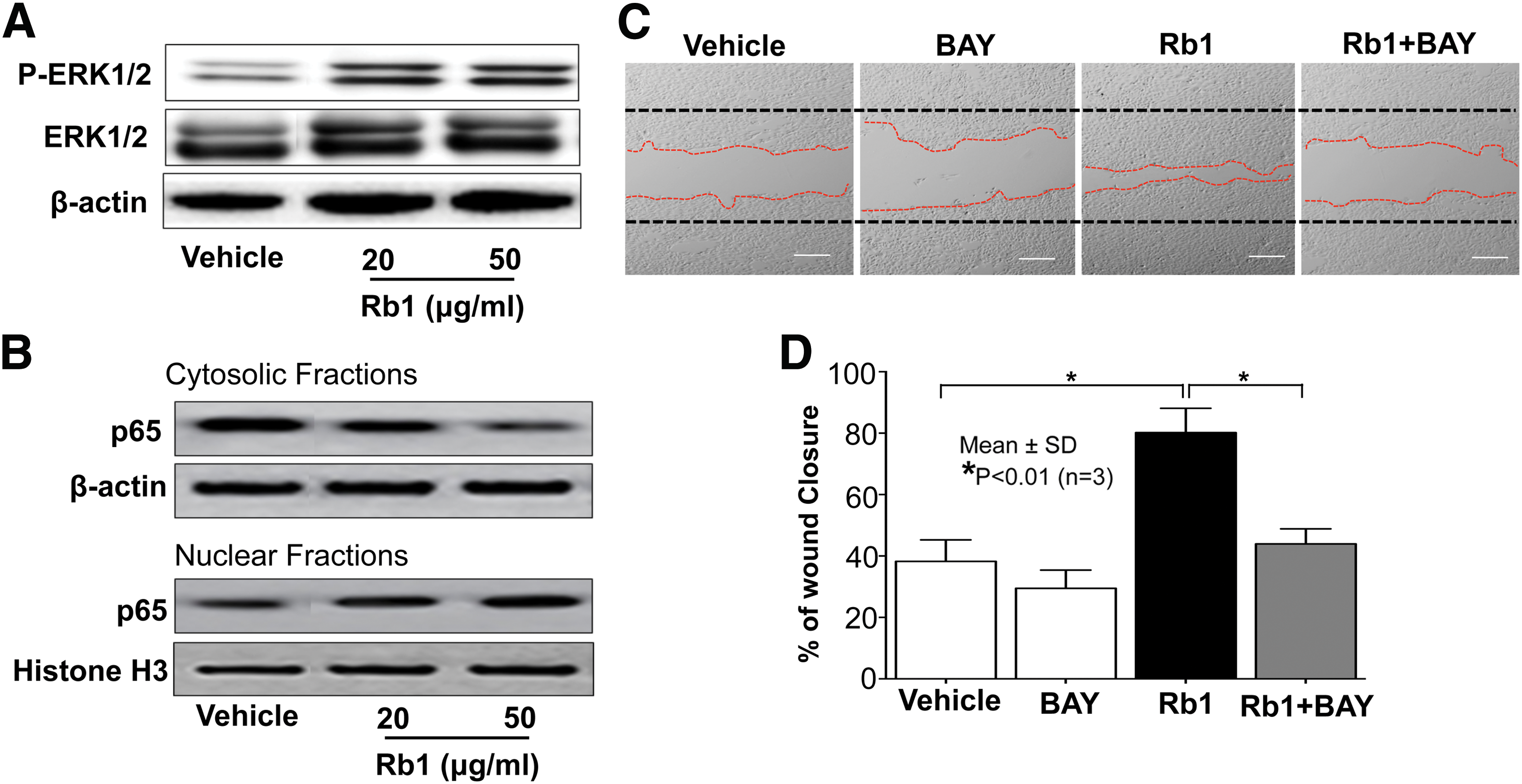
ERK1/2-NF-κB activation is responsible for Rb1-mediated stimulation of KC migration. KC were grown to confluence and in vitro wound was produced using a sterile scraper. After wounding, KC were pretreated with a specific inhibitor of NF-κB, BAY11-7082 (BAY, 1 μM), followed by incubation with Rb1 (50 μg/mL). Alterations of phosphorylated ERK1/2 (Thr202/Tyr204), ERK1/2, and NF-κB p65 in cytosolic or nuclear fractions were assessed by Western immunoblot analysis
Discussion
Ginsenosides, including Rb1, are triterpene saponins, and have multiple biological activities as follows: (1) anti-inflammatory effects by reducing the production of inducible nitric oxide synthase, cyclooxygenase-2, and proinflammatory cytokines 34 –37 ; (2) regulation of proliferation/differentiation/apoptosis in various cell types through the modulation of intracellular signaling molecules 38 –40 ; (3) modulation of calcium influx, leading to alteration in calcium-dependent cellular signaling, such as MAPK, PI3K, and PKC 41 ; (4) immunomodulating effects by regulating complex immune responses of different innate and adaptive immune cells 42 ; and (5) regulation of cell migration, invasion, and angiogenesis in different cells. In addition, because ginsenosides can modulate such cellular/biological functions in multiple tissues, their activities are being studied in diverse clinical settings for the treatment of different diseases, including cardiovascular disease, diabetes mellitus, cancers, obesity, and neurodegenerative disorders. 42 Of over 30 ginsenosides, in particular, Rb1 has a paradoxical effect on proliferation and migration in different type of cells, that is, normal cells versus cancer cells. Rb1 stimulates the reepithelialization and wound healing processes by increasing proliferation and migration of normal cells/tissues, including skin, 43,44 whereas, growth and invasion of tumor cells are inhibited by Rb1 treatment through their abilities to attenuate cellular signaling pathways associated with proliferation and migration. 43,45 Although a detailed mechanism is still not fully elucidated, S1P is one of the possible signaling mediators that could explain these cellular responses regulated by Rb1. Similar to Rb1, different roles of S1P are also highlighted in between non-cancer cells and cancer cells. 46 Prior studies demonstrated that S1P significantly stimulates migration of non-cancer cells, including KC, but S1P declines migration and invasion of certain cancer cells, for example, melanoma, osteosarcoma, and breast cancer cells. 47 While these previous findings likely suggest that Rb1-induced biological functions (at least migration) would be regulated by an S1P-mediated cellular mechanism, to our knowledge, this work is the first study to demonstrate that an S1P-dependent mechanism accounts for the Rb1-induced enhancement of KC migration.
Cells employ two pathways to generate ceramide: (1) de novo synthesis through the enzymatic condensation of serine and palmitoyl-CoA by serine palmitoyltransferase; and (2) the SMase pathway for the conversion of SM into ceramide. The latter is the most rapid, abundant mechanism to generate cellular ceramide, followed by the metabolic conversion of ceramide to S1P by SPHK1. S1P is then degraded into phosphoethanolamine and (2E)-hexadecenal by S1P lyase. As such cellular S1P levels are tightly regulated by metabolic conversion of SMase-mediated produced ceramide into S1P by SPHK1 and the levels of S1P degradation by S1P lyase. In these studies, we show that Rb1 exploits this pathway, that is, increased SPHK1 activity and suppressed S1P lyase activity, to increase cellular S1P. While S1P-mediated signaling of cellular functions is often dependent upon prior binding to one or more of five known S1P receptors (S1P1 to S1P5), 48 our results demonstrate that Rb1-induced stimulation of KC migration likely occurs by an S1P receptor-dependent mechanism. A number of prior studies revealed that S1P receptors differently regulate cell migration. S1P2 significantly decreased cell migration through an evident Rho activation and induction of stress fibers, 49 whereas S1P1 and S1P3 enhance cell migration by increased expression/activation of signaling protein-associated migration. 50 –52 This difference is caused by expression levels of each receptor. Collectively, S1P blocks migration in cells dominantly expressing S1P2, whereas S1P obviously stimulates cell migration in cells, including KC, through S1P1- and/or S1P3-dependent mechanism. Although further studies are needed to clarify which subtype(s) of S1P receptor is responsible for Rb1-induced increases in migration, we here demonstrated that Rb1-mediated increase in S1P enhances KC migration through likely either S1P1- and/or S1P3-dependent signaling pathway(s). Finally, we further elucidated downstream mediators of the Rb1-mediated activation of S1P receptor(s), which signal to increase phosphorylation of ERK1/2, leading to activation of NF-κB, which is a critical transcription factor found in the promoter regions of MMP-2 and MMP-9. Since prior studies revealed that both MMP-2 and MMP-9 stimulate the movement of KC by modifying adhesion to type IV collagen or by cleaving laminin 332, leading to enhanced wound closure/repair, 53,54 MMP-2-/-9-mediated cellular responses are considered essential for the Rb1-mediated stimulation of KC migration.
In summary, we demonstrated a previously unidentified signaling mechanism of Rb1 that facilitates cell migration, that is, Rb1 enhances KC migration through production of important migration factors, MMP-2 and MMP-9 signaling by an S1P/S1P receptor-mediated ERK1/2→NF-κB mechanism.
Footnotes
Acknowledgments
This research was supported by Hallym University Research Fund, 2017 (HRF-201703-004), by the Korea Institute for Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through High Value-added Food Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA; 117063031SB101; to K.P.). This research was also supported by a grant (714001-07) from the Center for Industrialization of Natural Nutraceuticals through the Agriculture, Food and Rural Affairs Research Center Support Program, MAFRA, Republic of Korea (to Y.J.).
Author Disclosure Statement
No competing financial interests exist.