
Abstract
Methionine and choline, which are essential nutrients for mammalian animals, are important for cell composition, as metabolic factors, and for the synthesis of other biochemical compounds for cell metabolism. Methionine and choline, which are methyl group donors, play key roles in the homocysteine cycle and neuronal development and maintenance. In this study, we investigated the effects of methionine and choline deficiency on adult hippocampal neurogenesis and neural stem cell (NSC) lineage in the adult stage. For this study, we divided C57BL/6 mice into three groups as follows: normal chow (NC)-fed, methionine choline sufficient (MCS) diet-fed, and methionine choline deficient (MCD) diet-fed mice. The mice were fed the NC, MCS, and MCD diets for 4 weeks from the age of 8 weeks. MCD diet-fed mice showed significantly decreased proliferation and differentiation of NSCs when compared with the NC diet-fed or MCS diet-fed mice. In addition, the survival of newly generated neurons was critically impaired in the MCD diet-fed mice. We confirmed a decrease in the proliferation and differentiation of NSCs after 4 weeks of MCD diet administration, compared with that in NC- and MCS diet-fed mice. MCD diet critically impaired NSCs survival and survival of neurons during the 4 weeks. The number of phosphorylated cyclic AMP response element binding (pCREB) protein immunoreactive nuclei was decreased in the MCD diet-fed mice compared with that in the NC- or MCS diet-fed group. These results suggest that suitable levels of methionine and choline are essential for the maintenance of hippocampal neurogenesis in mice and affect NSC proliferation and differentiation through phosphorylation of CREB.
Introduction
Methionine is an essential amino acid for mammals and is used in cell growth and development. As a metabolite in amino acid biosynthesis and a cofactor of antioxidant enzymes, methionine is a key amino acid for protein and enzyme synthesis. 1 Circulating methionine can cross the blood–brain barrier and is often incorporated as the first amino acid during protein synthesis in the brain. 2 Methionine is metabolized by methionine adenosyltransferase to S-adenosylmethionine (SAM), which is the most critical methyl donor and sequentially converted to S-adenosylhomocysteine and homocysteine in the body. 3
Choline was also defined as an essential nutrient by the Institute of Medicine (U.S.) Standing Committee on the Scientific Evaluation of Dietary Reference. 4 Dietary choline is a source of phosphocholine, phosphatidylcholine, and sphingomyelin as a compartment of cell membrane lipids, and it is also a source of acetylcholine (ACh), which exists in neuronal synapses. 5,6 As a trimethyl amine, choline is converted to betaine in the mitochondria, which is then used in the formation of methionine from homocysteine. 7 Choline and methionine contribute to the formation of SAM and glutathione. 1,7 Methionine and choline are critical and basic nutrients that are required in cell metabolism and neuronal network. 1,8
Accumulating evidence indicates that diet can modulate adult hippocampal neurogenesis (AHN) depending on conditional diets, different compositions of nutrients, deficiency or sufficiency of specific amino acids, or vital elements. Studies on diet-induced impairment of AHN report that diets deficient in thiamine, 9 zinc, 10 and folate 11 decrease or inhibit proliferation of neural stem cells (NSCs). In addition, there is evidence that high-fat diets also impair AHN in C57BL/6N and C3H/HeN mice. 12 In contrast, omega-3 fatty acids, 13 flavonoids, 14 and curcumin 15 increase the proliferation of NSCs.
Human data show that dietary methionine during gestation is associated with better cognitive function of offspring. 16 Higher content of methyl donors in maternal diet increases the in vitro proliferation of neural stem/progenitor cells isolated from rat embryonic day 19 fetuses. 17 In a genetic mouse model for Down syndrome, the Ts65Dn mouse, maternal choline supplementation increases hippocampal neurogenesis. 18 However, these studies have focused on the effects of methionine and choline in pathological, not physiological conditions or in the postnatal response after maternal choline supplementation.
In addition, there are few reports on the methionine choline deficient (MCD) effects on hippocampal neurogenesis. In this study, therefore, we investigated the effects of abnormal levels of methionine and choline on AHN in the non-transgenic and healthy C57BL/6 mice through in vivo studies. We assessed AHN in C57BL/6 mice fed an
Materials and Methods
Experimental animals
Seven-week-old C57BL/6J male mice were purchased from Japan SLC, Inc. (Shizuoka, Japan). The animals were housed in a specific pathogen-free animal facility maintained at 23°C with 60% humidity, on a 12 h/12 h light/dark cycle, with ad libitum access to food and tap water. The handling and care of the animals conformed to the established guidelines, which were compliant with current international laws and policies (NIH Guide for the Care and Use of Laboratory Animals, NIH Publication No. 85-23, 1985, revised 2011) and were approved by the Institutional Animal Care and Use Committee (IACUC) of Seoul National University (SNU-130830-1). All experiments were conducted in a way that the number of animals used and the suffering caused by the study methodology were minimized as much as possible.
Diet administration
After 1 week of acclimation to laboratory conditions, the mice were divided into three groups (n = 5 in each group): NC (Purina 5008 diet; Purina Korea, South Korea), MCS (no. 518754; Dyets, Bethlehem, PA, USA), and MCD (no. 518810; Dyets) feeding groups. Apart from their methionine and choline content, the MCD and MCS diets contained the same nutrients. The NC group was used as a general control to compare the levels of proliferation and differentiation of NSCs in different diets. All the mice fed the NC, MCD, and MCS diets had access to the diets ad libitum for 4 weeks, until they were 12 weeks of age.
Measurement of body weight and calorie intake
Mouse body weights were measured at 10:00 a.m. every week on a Monday and at the end of the experiment. Food intake was measured, and it was corrected for spillage by weighing the jars containing food, every week between 9:00 and 10:00 a.m. Food intake data were calculated from the data on intake of each week, over the 4-week experimental period, and expressed as kcal/mouse/week. NC and MCS contained different levels of methionine and choline: in NC (methionine 4.3 g/kg and choline chloride 2 g/kg) and MCS (methionine 1.7 g/kg and choline bitartrate 14.48 g/kg); MCD diet had neither methionine nor choline. NC and MCS have different sources of choline (NC, choline chloride and MCS, choline bitartrate), and choline chloride and choline bitartrate have different dissociation rates in water (74% and 41%, respectively). 19 Total methionine and choline intake was calibrated by dissociation rates and calculated in NC and MCS groups.
Labeling of newly generated cells
To label newly generated cells in the hippocampus, intraperitoneal injections of 5-bromo-2′-deoxyuridine (BrdU, 50 mg/kg; Sigma-Aldrich, St. Louis, MO, USA) were administered to all the mice twice daily (8:00, 20:00) for 3 consecutive days after the mice started on the NC, MCD, and MCS diets, at the age of 8 weeks.
Tissue preparation
Animals (n = 5 in each group) were anesthetized with an intraperitoneal injection of 1.5 g/kg urethane (Sigma-Aldrich), and they were perfused transcardially with 0.1 M phosphate-buffered saline (PBS, pH 7.4) followed by 4% paraformaldehyde in 0.1 M PBS. The brains were dissected and post-fixed in the same fixative for 12 h. The brain tissues were cryoprotected with an overnight immersion in 30% sucrose. Brain sections, which were 30-μm thick, were serially sectioned in the coronal plane, by using a cryostat (Leica, Wetzlar, Germany). The sections were collected in six-well plates containing PBS and stored at −20°C for further processing.
Immunohistochemistry
To obtain accurate data for immunohistochemistry, the free-floating sections from all animals were processed carefully under the same conditions. For each animal, tissue sections between −1.46 and −2.46 mm to bregma were selected in reference to the mouse atlas by Franklin and Paxinos. 20 Ten sections, 90 μm apart from each other, were sequentially treated with 0.3% hydrogen peroxide (H2O2) in PBS for 30 min, and 10% normal goat or rabbit serum in 0.05 M PBS for 30 min. The sections were then incubated with a rabbit anti-Ki67 antibody (1:1000; Abcam, Cambridge, United Kingdom), goat anti-doublecortin (DCX) antibody (1:50; Santa Cruz Biotechnology, Santa Cruz, CA, USA), or rabbit anti-phosphorylated cyclic AMP response element binding (pCREB) protein antibody (1:400; Cell Signaling Technology, Beverly, MA, USA) overnight at 25°C, and they were subsequently treated with a biotinylated goat anti-rabbit IgG, rabbit anti-goat IgG, or fluorescein isothiocyanate (FITC)-conjugated anti-rabbit IgG, and a streptavidin-peroxidase complex (1:200; Vector Labs, Burlingame, CA, USA). Sections were visualized with a reaction with 3,3′-diaminobenzidine tetrachloride (Sigma-Aldrich) in 0.1 M Tris-HCl buffer (pH 7.2), dehydrated, and mounted in Canada balsam (Kanto Chemical, Tokyo, Japan) onto gelatin-coated slides.
Double immunofluorescence
For BrdU and NeuN double immunofluorescence, the sections were treated with 2 N HCl for 30 min at 37°C, and they were incubated with a mixture of mouse anti-NeuN (1:1000; Millipore, Temecula, CA, USA) and rat anti-BrdU (1:200; Abd Serotec, Bio-Rad Laboratories, Inc., Grand Island, NY, USA) for 2 h at 25°C, followed by incubation overnight at 4°C. After washing with PBS, the sections were subsequently incubated with secondary antibodies, FITC-conjugated goat anti-mouse IgG (1:100; Jackson ImmunoResearch, PA, USA) and Cy3-conjugated goat anti-rat IgG (1:100; Jackson ImmunoResearch) for 2 h. The sections were mounted on silane-coated slides with DAPI-containing mounting medium (Vector Labs) for staining of the nuclei.
Microscopic analysis
Two independent, masked investigators counted the Ki67, DCX, BrdU, or BrdU/NeuN-labeled cells in the dentate gyrus at 400 × magnifications under a light microscope (BX51; Olympus, Tokyo, Japan). All cells labeled with Ki67, DCX, BrdU, or BrdU/NeuN were counted bilaterally in 10 sections (90 μm apart from each other) across the entire dentate gyrus between −1.46 mm and −2.46 mm to bregma in reference to the mouse atlas by Franklin and Paxinos. 20 The granule cell layer and molecular layer of the dentate gyrus (100 × primary magnification) were selected and images were calibrated into an array for relative optical density (ROD) analysis, by using ImageJ v. 1.60 software (National Institutes of Health, Bethesda, MD, USA). Each pixel resolution was 256 gray levels, and the intensity of DCX immunoreactivity was evaluated by ROD, which was obtained after transformation of the mean gray level by using the formula: ROD = log (256/mean gray level). The ROD of background staining was determined in unlabeled portions of the sections by using Photoshop CC software (Adobe Systems, Inc., San Jose, CA, USA), and this value was subtracted to correct for nonspecific staining, by using ImageJ v. 1.60 software (National Institutes of Health). The ROD data are expressed as a percentage of the NC group values (set to 100%).
Statistical analyses
Statistical analysis was performed by using SPSS V.20.1 (IBM Corporation, Armonk, NY, USA). Experimental groups were compared by using one-way analysis of variance, followed by post hoc analysis with Tukey's honest significant difference test.
Results
Effects of MCD and MCS diets on body weight and calorie intake
At the age of 8 weeks, mice fed with NC or MCS diets showed similar body weights; however, the MCD diet-fed group had significantly lower body weights compared with the NC diet-fed group, from week 9, and continuously decreased up to week 12 (Fig. 1A). However, the MCD diet-fed mice consumed relatively higher calories than the MCS and NC diet-fed groups (Fig. 1B). At the end of the experiment, the mice of the NC and MCS diet-fed groups showed a tendency to gain more body weight with age (Fig. 1A). Body weight gain was higher in the MCS group at weeks 9, 10, and 11 compared with the NC diet-fed group (Fig. 1A). Similarly, the caloric intake was also relatively higher in the MCS group during weeks 10, 11, and 12 compared with that of the NC diet-fed group (Fig. 1B). Actual methionine intake was lower in the MCS group during all periods compared with the NC diet-fed group (Fig. 1C). Choline intake was higher in the MCS group during all periods compared with the NC diet-fed group (Fig. 1D).
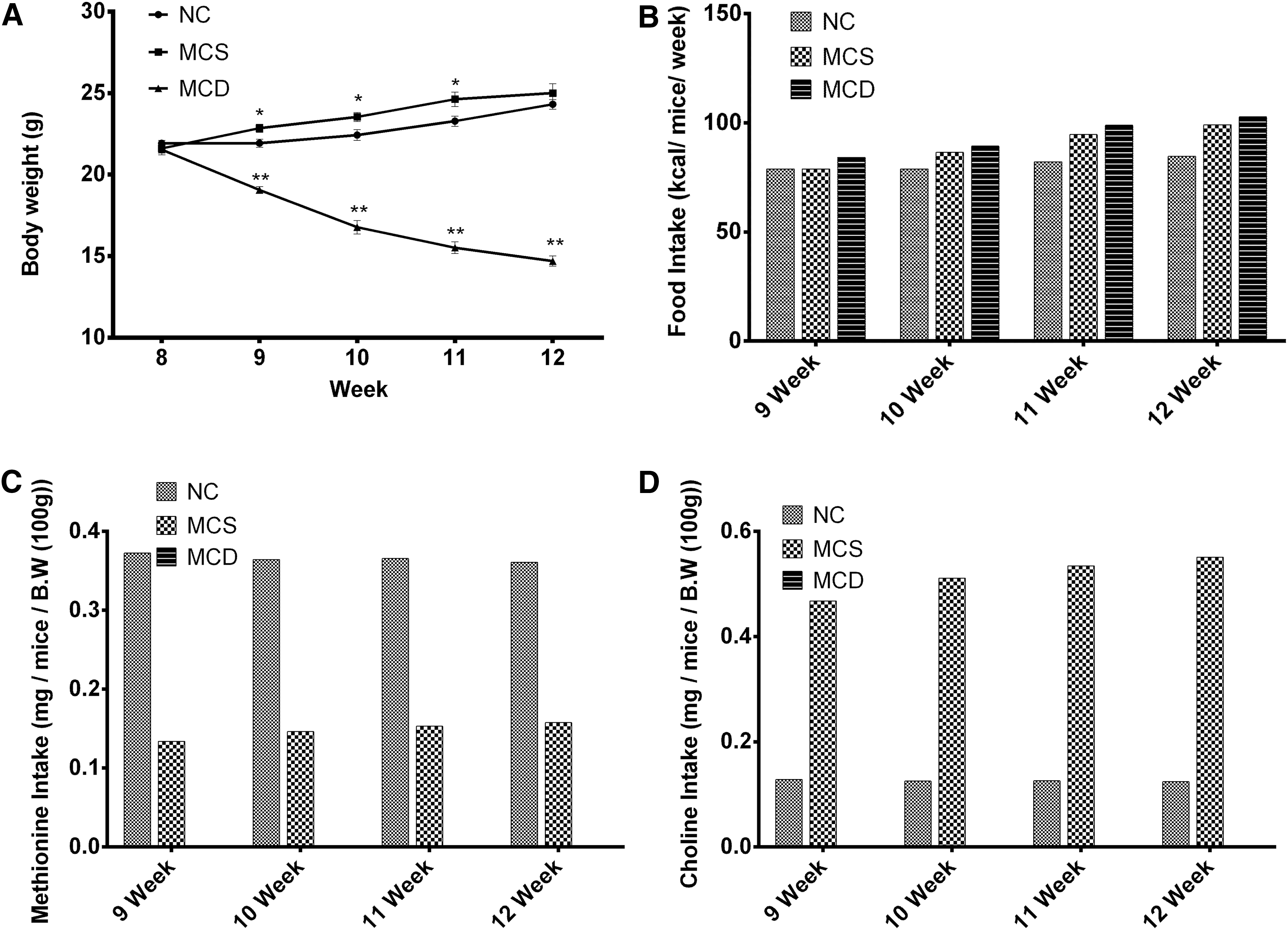
Body weight changes during experimental periods
Effects of MCD and MCS diet on cell proliferation in the dentate gyrus
To determine the effects of MCD on the proliferating NSCs, we counted the number of cells that were positive for Ki67, a marker of cell proliferation used for NSCs, in the subgranular zone of the dentate gyrus. The mean number of Ki67-positive cells in the NC diet-fed mice was 22.40 ± 1.29 (Fig. 2A, B). Compared with the NC diet-fed group, the numbers of Ki67-positive cells were lower in the MCD and MCS diet-fed groups (Fig. 2C–F). There were significant differences in the number of Ki67-positive cells between NC and MCS diet-fed groups, and the number of Ki67-positive cells in the MCS diet-fed group was 15.60 ± 2.29, that is, 74.11% compared with the NC diet-fed group (Fig. 2G). In the MCD diet-fed group, the number of Ki67-positive cells was also significantly decreased and was 6.40 ± 0.93, which was 41.02% of that in the MCS diet-fed group (Fig. 2G).
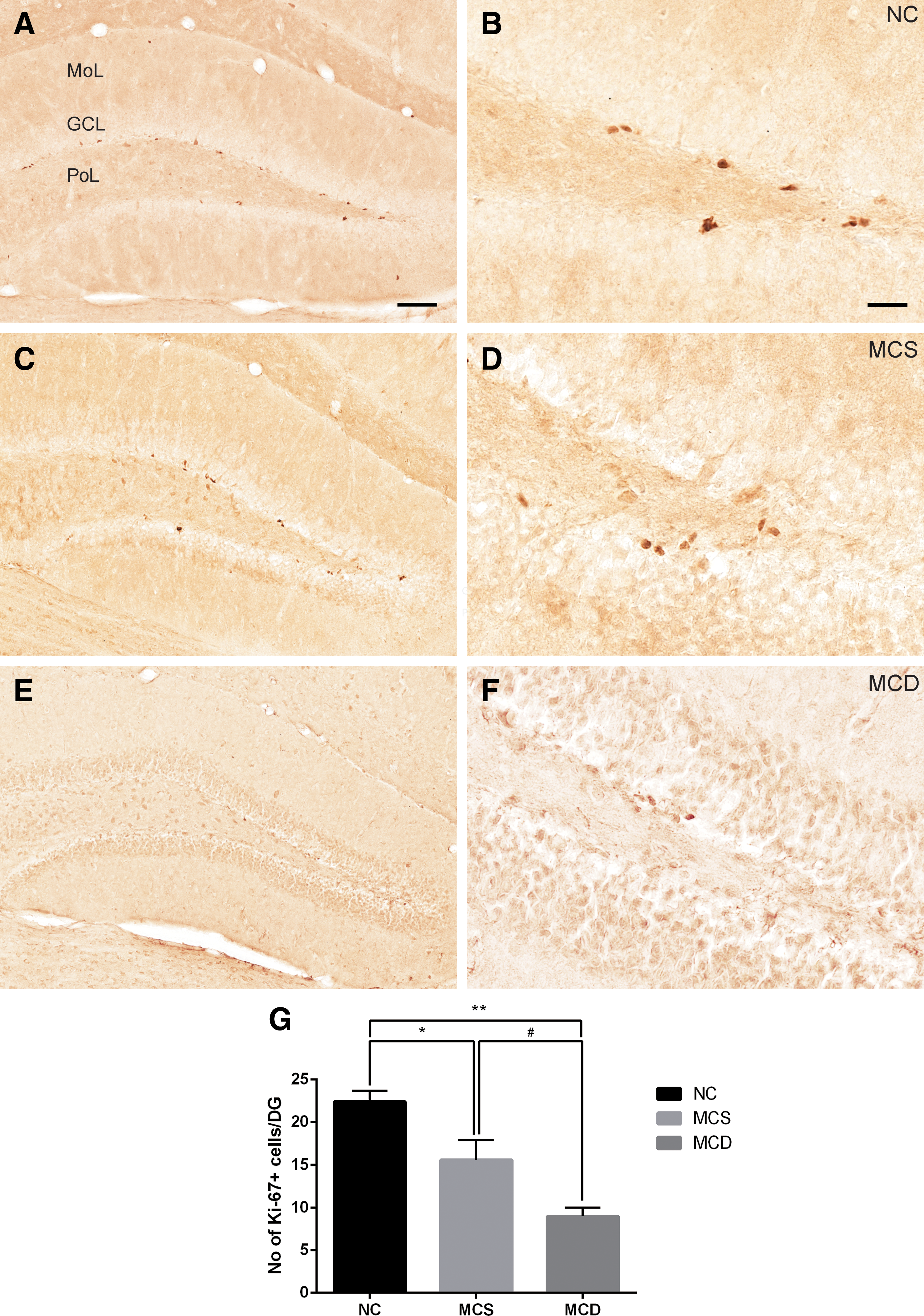
Immunohistochemistry for Ki67 in the dentate gyrus of NC
Effects of MCD and MCS diet on cell proliferation in the dentate gyrus
To investigate the effects of MCD on the differentiation of NSCs into neuroblasts, the number of neuroblasts immunoreactive for DCX, a marker for differentiated and migrating neuroblasts, was measured in the dentate gyrus. In the NC diet-fed group, DCX-immunoreactive neuroblasts were abundant in the dentate gyrus (Fig. 3A, B); the mean number of DCX-immunoreactive neuroblasts was 150.40 ± 5.09 (Fig. 3G). Compared with the NC diet-fed group, however, DCX-immunoreactive neuroblasts were significantly lower in the dentate gyrus of MCS and MCD diet-fed groups (Fig. 3C–F). The decrease in the number of DCX-immunoreactive neuroblasts was particularly more prominent in the MCD diet-fed group compared with those in the MCS diet-fed group (Fig. 3G). The mean number of DCX-immunoreactive neuroblasts was 118.80 ± 6.32 and 70.80 ± 5.11 in the MCS diet-fed and MCD diet-fed groups, respectively. In the MCD diet-fed group, the number of DCX-immunoreactive neuroblasts was 60.82% of that in the MCS diet-fed group (Fig. 3G). In addition, we also measured ROD of DCX immunoreactive neuroblasts in the molecular layer and granule cell layer of the dentate gyrus. In the MCS and MCD groups, DCX immunoreactivity in the granule cell layer and molecular layer of dentate gyrus showed a decrease in ROD compared with the NC group (Fig. 3H). The ROD of DCX-immunoreactive neuroblasts was 76.27% in the MCD compared with the MCS diet-fed group.
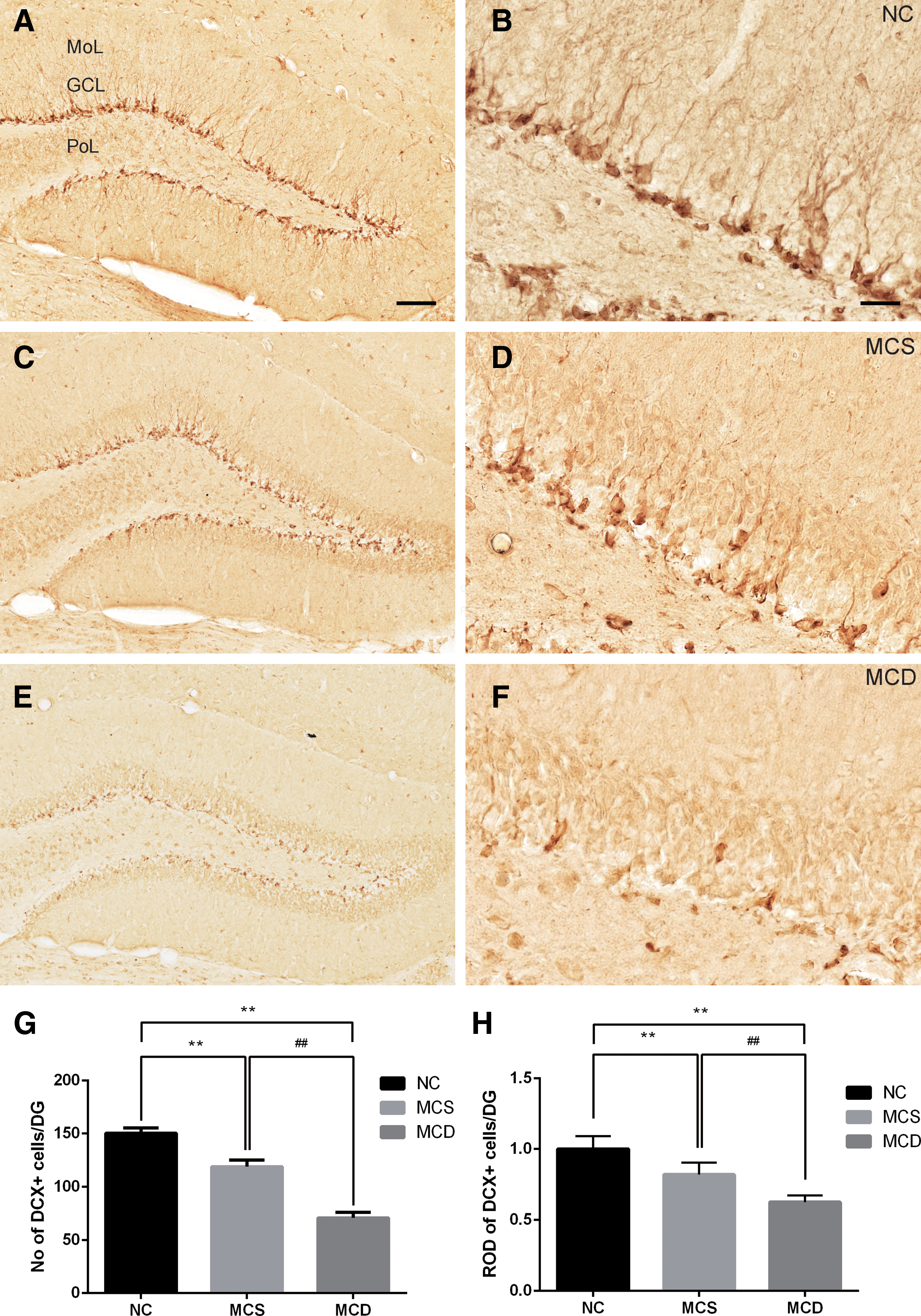
Immunohistochemistry for DCX in the dentate gyrus of NC
Effects of MCD and MCS diet on the survival of newly generated cells
We evaluated the effects of MCS and MCD on survival into mature neurons of NSCs by double immunofluorescence for BrdU and NeuN. In the NC diet-fed group, NeuN and BrdU double-positive cells were found mainly in the subgranular zone and granule cell layer of the dentate gyrus and the mean number of BrdU and NeuN double-positive cells was 23.00 ± 2.24 (Fig. 4A, B, G). In the MCS and MCD diet-fed groups, fewer BrdU and NeuN double-positive cells were found in the dentate gyrus compared with that in the NC diet-fed group (Fig. 4C–F). The number of BrdU and NeuN double-positive cells was substantially decreased in the MCD diet-fed group compared with that in the MCS diet-fed group. In the MCD group, the mean number of BrdU and NeuN double-positive cells was decreased by 22.24% compared with that of the MCS group (Fig. 4G). The ratio percentage of BrdU and NeuN double-positive cells per BrdU-positive cells was calculated in all groups. The proportion of BrdU-/NeuN-positive cells was relatively sequentially lower in all groups, with no significance (Fig. 4H).
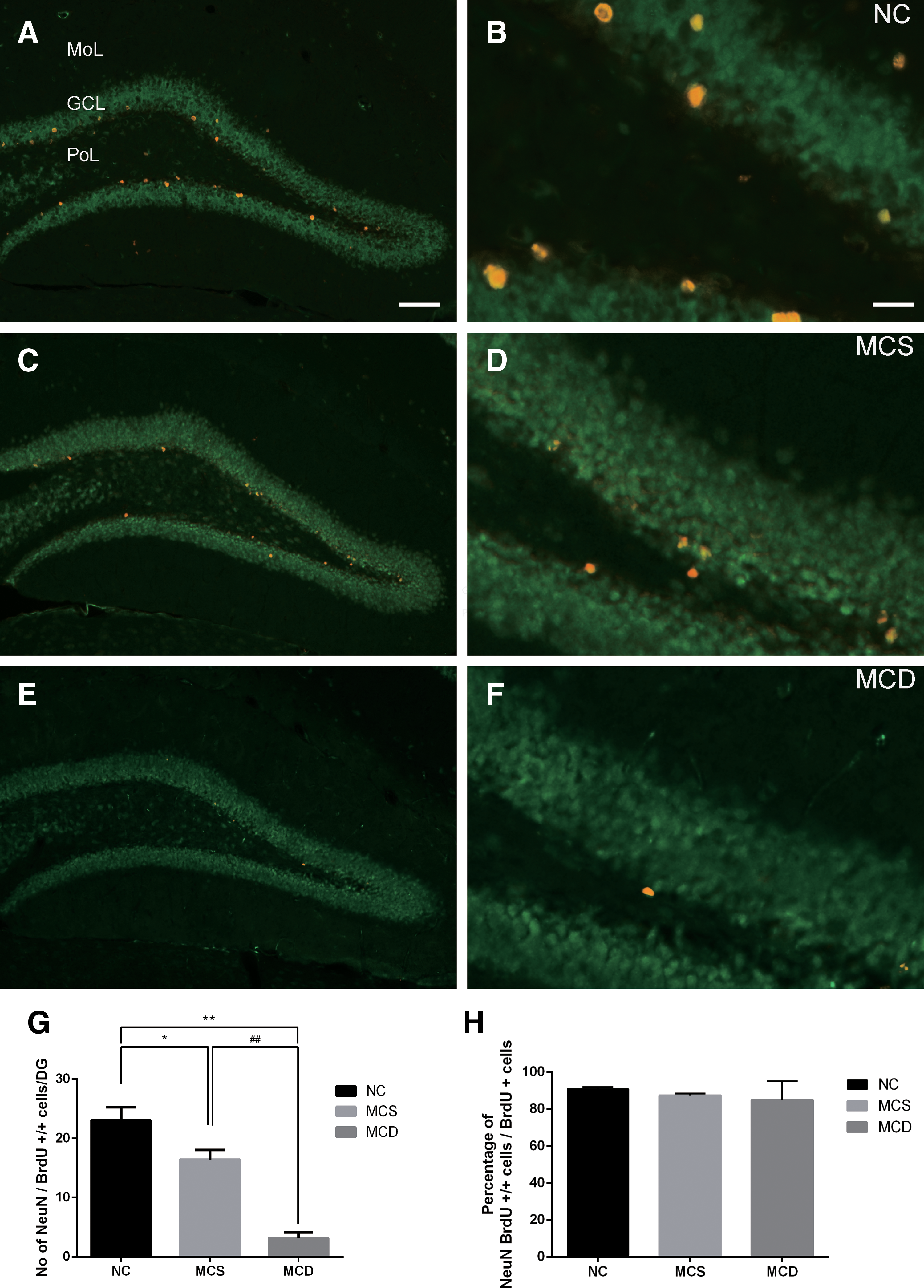
Double immunofluorescent staining for BrdU (red) and NeuN (green) in the dentate gyrus of NC
Effects of MCD and MCS diet on the phosphorylation of CREB
To investigate the mechanisms of MCD on hippocampal neurogenesis, pCREB immunoreactivity was observed in the dentate gyrus. In the NC diet-fed group, pCREB-immunoreactive nuclei were found in the dentate gyrus (Fig. 5A, 5B) and the mean number of pCREB-immunoreactive nuclei was 118.60 ± 8.93 (Fig. 5G). In the MCS and MCD diet-fed groups, the number of pCREB-immunoreactive nuclei significantly decreased in the dentate gyrus (Fig. 5C–F). The mean number of pCREB-immunoreactive nuclei was 85.20 ± 7.92 and 20.20 ± 2.63 in the MCS diet-fed and MCD diet-fed groups, respectively (Fig. 5G).

Immunohistochemistry for pCREB in the dentate gyrus of NC
Discussion
Although the caloric intake was relatively higher in the MCD diet-fed group, the administration of an MCD diet significantly reduced the body weights of mice. In addition, the MCS diet-fed animals showed reduced weight gain compared with the NC diet-fed animals. This result is supported by previous studies, which showed significant weight loss although no evidence of insulin resistance in their MCD diet models. 21,22 In this study, we investigated the effects of deprivation and supplementation of methionine and choline on NSC proliferation, differentiation, and survival into mature neurons. MCD diets, when fed for 4 weeks, caused decreases in the proliferation, differentiation, and survival of NSCs into mature neurons compared with MCS diet-fed groups.
Numerous studies have elucidated the relative roles of methionine and choline in early stage hippocampal development. 23,24 Choline plays an important role in the rat hippocampus and septum development at embryonic stages 23 and it is an important factor for the regulation of mitosis, apoptosis, and in determining the cell fate of neural and endothelial progenitor cells. 25 In addition, choline supplementation at prenatal stages upregulates mitogen-activated protein kinase and cAMP-response element binding protein signaling cascades, and it enhances hippocampal maturation. 26 Prenatal and postnatal choline supplementation enhanced spatial memory function and long-term facilitation. 27 In contrast, deficiency of choline during the embryonic stage (especially p12–p17) increases apoptotic cells in the hippocampus. 24
In a mouse model of deficiency of methionine synthase reductase, inhibition of methionine synthesis exhibits impairment of memory functions and choline metabolism. 28 Methionine deprivation causes cell cycle arrest, impairment of survival, and it poises differentiation of human pluripotent stem cells with a decrease in SAM and p58–p38 signaling activation. 29 Dietary choline deprivation alters gene-specific methylation in fetal mouse brain, 30 and dietary choline deprivation also decreases proliferation of embryonic neural progenitor cells via inhibiting the epidermal growth factor signaling pathway. 31
Studies of methionine and choline deprivation investigated during prenatal stages showed limitation in adult stages and the AHN process. Our data show a decrease of proliferation, differentiation, and survival of NSCs in MCD diet-fed mice, revealing that methionine and choline are essential nutrients for the AHN process.
In biochemical metabolism, methionine, choline, betaine, and vitamin B12 are mediators of homocysteine metabolism, whereas methionine, vitamins B12, and folate are indispensable methyl donors. 32,33 Nutrients involved in the homocysteine-methionine cycle, including folate and vitamin B12, play a key role in neuronal development and maintenance throughout the lifespan. 34 The methionine and homocysteine cycle maintain the balance of each other in normal status. 33 A deficiency of methionine, however, can influence the methionine and homocysteine cycle, increasing homocysteine levels in the blood (hyperhomocysteinemia). 33 In hippocampal function, homocysteine imbalance causes cognitive decline in aging and neurodegenerative disease. 35 –37
In NSCs cultures, addition of homocysteine inhibits the proliferation of NSCs via the reduction in DNA methylation, and it increases S-adenosylhomocysteine and decreases SAM. 38 Further, elevated homocysteine inhibits proliferation of neural precursor cells in the DG in the adult brain, by inhibiting basic fibroblast growth factor (bFGF) and bFGF-dependent extracellular regulated kinase 1/2 cascades both in vivo and in vitro in mouse models. 39 Although the direct correlation between methionine and neurogenesis has not been investigated in developmental or adult stages, the methionine-homocysteine metabolic cycle can regulate NSCs proliferation and cell fate. In our study, we confirmed that methionine and choline deficiency impaired the proliferation, differentiation, and survival of NSCs in the hippocampus.
This finding could be attributed to the association between methionine and choline and DNA methylation, which is the key factor regulating neural activities and memory formation. 40 Conversion from methionine to SAM is a critical step for the methyl donor; this reaction contributes to the methylation reaction, and, thus, gene regulation. 32 In addition, dietary choline is another major methyl group donor source; choline is needed for the synthesis of methionine and SAM, and these factors also are used in DNA methylation. 41 DNA methylation is catalyzed by DNA methyltransferases (i.e., DNMT1, DNMT2, and DNMT3). 42 In hippocampal NSCs, DNA methylation regulates cell cycle exit and differentiation, via the induction of regulatory genes that are specific for differentiation. 43
In addition, methionine treatment increases brain-derived neurotrophic factor (BDNF) DNA methylation in the hippocampus and rescues hippocampal-dependent memory function. 44 Similarly, low choline status decreases in the contents of methionine and SAM, which induces DNA and histone hypomethylation, with changes in gene expression in brain progenitor cells. 45 Gene expression changes cause a decrease in mitosis, differentiation, and migration of fetal brain progenitor cells. 45 Choline and methionine play a role in SAM synthesis and regulation of methylation and, thus, affect the adult brain. This evidence provides clues to explain our data about the impairment of AHN. Critical impairment in the survival of NSCs in the MCD diet-fed group indicates that dietary methionine and choline can be key regulators for the survival and maturation of NSCs.
Methionine and choline are related with neuropathology, since many studies show evidence of the dysfunction in their metabolism in neurodegenerative disease. For instance, the symptoms of cognitive impairment and dementia in Alzheimer's disease (AD) and Parkinson's disease (PD) are accompanied by dysfunction of the cholinergic and dopaminergic pathways. 46,47 Further, in AD, cortical acetyltransferase activity showed a lower correlation with dementia and histological changes. 48 Further, impairment of ACh synthesis in AD patients was correlated with cognitive impairment. 48,49 In a mouse model for PD, impairment of neurogenesis in the hippocampus was accompanied by elevated homocysteine level in plasma and cerebrospinal fluid, and elevated homocysteine caused dysfunction of mitochondria, protein, oxidative stress responding, and apoptosis induction. 50,51
Further, a patient with AD showed impairment in the re-methylation of homocysteine to methionine. 52 Diet-induced high-level homocysteine in 3xTg mice also exacerbated the cognitive deficit, the β-amyloid, and tau pathology. 53 These results indicated that the methionine-related homocysteine cycle was disrupted in neurodegenerative diseases. Conversely, methionine and choline supplementation can restore hippocampal-related function. In a kainic acid-induced epilepsy model, supplementation of methionine increased BDNF DNA methylation, and it restored many behavioral test scores that were impaired in epilepsy. 54 In the transgenic Down syndrome mouse model (Ts65Dn), dietary choline supplementation improved AHN and spatial learning performance. 18
In this study, the MCD diet impaired AHN, indicating either a direct or an indirect correlation with pathological status. CREB is a ubiquitous transcription factor that is highly expressed in the brain, and it serves the neuronal survival and memory formation in the hippocampus. 55 In this study, we observed the pCREB immunoreactivity to investigate the mechanisms of reduction of neurogenesis by MCS diets because cholinergic activation by carbachol significantly increased pCREB in the hippocampal slice. 56 pCREB immunoreactivity was dramatically decreased in the MCD diet-fed group compared with that in the MCS diet-fed group. This result suggests that MCD diets impair hippocampal neurogenesis by decreasing pCREB expression in the dentate gyrus.
In this study, we observed the reduction of neurogenesis in the MCS diet-fed mice compared with that in the NC diet-fed mice. One possibility for the decreased AHN is hardness of diets. Soft diet-fed mice showed impairment of AHN, and those results were caused by decreased mastication.
57
The composition of the MCD diet was based on an
Another possibility that may explain the decreased AHN in the MCD diet-fed group was the different contents of methionine and choline in the MCD diet compared with the NC diet. From methionine and choline intake data, NC and MCD diet-fed groups showed different levels of methionine and choline intake. The MCS group showed lower methionine and higher choline intake than the NC feeding group. Differences in methionine and choline composition compared with the NC diet may be the cause of differences in the phenotypical changes of AHN; therefore, we needed more investigation of the balance of methionine and choline.
Another possibility is related to the fact that dietary choline plays a role in the synthesis and, thus, regulation of the neurotransmitter ACh in brain tissue. 58 ACh is synthesized from choline used as a neurotransmitter and compound of cell membrane in neuronal cells. 5,59 ACh and its receptor play a key role in the encoding of new memory in the hippocampus. 60 The hippocampal network circuit receives abundant cholinergic inputs, and the forebrain ACh is known to promote neurogenesis. 61 In a recent study, MCD diet administration increased acetylcholinesterase (AChE), an enzyme that degrades ACh, after 2, 4, and 6 weeks in the brain. 8 Transgenic mice, with inactivated AChE, show an increase in proliferation of hippocampal NSCs. 62 The impairment of AHN in this study provides evidence that the dietary supply of methionine and choline plays a role in hippocampus and NSCs.
Our results redefine the role that deficient and sufficient methionine and choline levels play in proliferation, differentiation, and survival of NSCs in adult mouse hippocampus. NSCs in the hippocampus transiently undergo cell mitosis and maturation process, and, thus, might need a source of nutrient choline and methionine. Our results demonstrated that MCD and MCS impaired AHN. This effect was most prominent in the MCD diet-fed mice. Further studies are warranted to elucidate the direct modulatory mechanism of methionine and choline during AHN.
Acknowledgments
This work was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2015R1D1A1A01059314) and by the Korea Mouse Phenotyping Project (NRF-2015M3A9D5A01076747) of the Ministry of Science, ICT and Future Planning through the National Research Foundation (NRF), Korea. This study was partially supported by the Research Institute for Veterinary Science, Seoul National University.
AUTHORS' CONTRIBUTIONS
All authors conceived the study. J.W.K. and Y.S.Y. designed the study. J.W.K. wrote the article, and I.K.H. and Y.S.Y. edited the article. H.Y.J., D.Y.Y., and J.W.K. conducted the animal experiments. N.S.M. and J.K.S. participated in designing and discussing the study. All authors have read and approved the final article.
Footnotes
Author Disclosure Statement
No competing financial interests exist.