
Abstract
Piceatannol (PIC) is a natural hydroxylated analog of resveratrol (RSV) and considered as a potential metabolic regulator. The purpose of this study was to compare the effects of PIC and RSV on parameters affecting inflammation, oxidative stress, and sirtuins (Sirt). Male C57BL/6J mice, 20 weeks old, were assigned to the following groups; (1) lean control, (2) high-fat diet control (HF), (3) HF_PIC, and (4) HF_RSV. Oral administration of PIC and RSV (10 mg/kg/day) for 4 weeks improved glucose control as shown by decreasing levels of area under the curve (AUC) during the oral glucose tolerance test compared with HF group. PIC improved glycemic control by increasing hepatic levels of insulin receptor and AMP-activated protein kinase. PIC increased the levels of Sirt1, Sirt3, and Sirt6 and also increased two downstream targets of Sirt, peroxisome proliferator-activated receptor gamma coactivator 1-alpha and forkhead box O1, in the liver. The inflammatory markers, interleukin (IL)-1 and IL-6, in the liver were downregulated by RSV treatment. Exposure to PIC and RSV significantly lowered hepatic levels of tumor necrosis factor-alpha. However, PIC and RSV treatments showed minimal effects on hepatic markers of oxidative stress. The levels of antioxidant enzyme, NAD(P)H:quinone oxidoreductase 1 (NQO1), were only increased in livers of RSV-treated mice compared with HF control mice. In conclusion, PIC was superior to an equal concentration of RSV in the regulation of Sirt and its downstream targets as well as insulin signaling-related parameters, while RSV potentially suppressed levels of proinflammatory markers and increased NQO1 protein levels.
Introduction
Piceatannol (PIC) is a natural analog of resveratrol (RSV) and its metabolic derivative. Both are polyphenolic compounds belonging to the stilbene class. 1 PIC and RSV are present in grapes, mulberry leaves/fruits, nuts, passion fruits, and berries 2 –4 and exhibit cardioprotective, antitumor, and antiaging effects. 1,5,6 Nonetheless, their use in pharmaceuticals and functional foods is limited by their relatively low solubility and bioavailability. PIC has an extra hydroxyl (OH) group compared with RSV, and this structural difference does not contribute to improved solubility. 7
PIC is derived from RSV through hydrolysis of cytochrome P4501B1 (CYP1B1), CYP1A1, and CYP1A2. 8,9 PIC exhibits antioxidative, antidiabetic, anti-inflammatory, and antitumor effects in various experimental models. 10 –14 A recent human intervention study demonstrated that supplementation of PIC from passion fruit improved several metabolic parameters of insulin sensitivity, blood pressure, and heart rate especially in overweight men. 15
Similarly, RSV improved several pathophysiological parameters in cardiovascular disease, cancer, and Alzheimer's disease. 16 –20 Moreover, RSV exerted antiobesity effects by inhibiting lipogenesis and increasing brown adipocyte differentiation in a mouse model of high-fat (HF) diet-induced obesity. 21,22 Evidence suggested that the regulatory effects of PIC are similar or often superior to those of RSV depending on experimental conditions. Previous experiments reported that PIC treatment led to higher metabolic stability in rats, 23 as well as enhanced astrocyte differentiation, 24 and stronger inhibitory activity of tyrosinase, monophenol oxidase in melanoma cells 25 compared with RSV. The difference may be due to the additional OH group in PIC compared with RSV. 26,27
PIC and RSV display anti-inflammatory, antioxidative, and antiaging activities. 12,14,26 Inflammation is an immunological protection mechanism against external stimulus. Infection exacerbates inflammatory response. However, metabolic inflammatory response by HF, high sugar, and high alcohol intake is generally slow and low grade without apparent symptoms in the early stages. However, long-term exposure to metabolic stress eventually triggers diseases accompanying excessive inflammation, including hepatitis, obesity, and atherosclerosis. 28,29
Experiments involving endothelial cells, lung epithelial cells, and mouse skin demonstrated anti-inflammatory effects of PIC through coordinated regulation of activator protein 1, cytochrome c oxidase subunit II, heme oxygenease-1, inducible nitric oxide synthase, and nuclear factor kappa B (NF-κB). 12,30,31 Furthermore, RSV and curcumin are food components with anti-inflammatory activities. 32,33 Metabolic inflammation often induces oxidative stress, or vice versa, through mutual factors (e.g., NF-κB). 34
Sirtuin (Sirt) is an antiaging protein acting through NAD+-dependent mono-ADP-ribosyltransferase or deacetylase activities. 35 Sirtuins inhibit the inflammatory state by suppressing activities of NF-κB and expression of cyclooxygenase-2 and inducible nitric oxide synthase, as well as by deacetylating the p65 subunit of NF-κB and receptor-interacting protein 1. 36 Moreover, sirtuin 1 had protective effects against lung oxidative stress by modulating forkhead box O3 (foxo3). 37 Existing evidence, including these findings, suggests that sirtuins may be key molecules mediating the anti-inflammatory and antioxidative activities of PIC and RSV. So far, findings regarding PIC- and RSV-mediated anti-inflammatory and antioxidative activities in the liver are limited, and the underlying mechanisms are unclear. Therefore, we compared the effects of PIC and RSV on factors related to inflammation, oxidative stress, and sirtuins in mice exposed to HF diet.
Materials and Methods
Animals
Male C57BL/6 mice were purchased from Raon Bio (Yongin, Korea) and housed under standard conditions (20°C ± 5°C, humidity 55% ± 5%, and a 12-h light/12-h dark cycle). The mice were group housed if possible. Standard rodent chow and water were made available ad libitum. All the experiments and animal care were performed strictly in accordance with animal welfare and other related ethical regulations approved by the Institutional Animal Care Committee of Chonnam National University (approval No. CNU IACUC-YB-2015-6).
Mice at 20 weeks of age were randomly divided into four groups (n = 37): (1) normal control (CON, n = 8), (2) high-fat diet control (HF, n = 12), (3) HF+PIC (HF_PIC, n = 8), and (4) HF+RSV (HF_RSV, n = 9). Normal control mice were fed a standard chow diet (10% kcal from fat; Research Diets, New Brunswick, NJ, USA), and the other groups were fed an HF diet (45% kcal from fat; Research Diets). PIC (10 mg/kg/day; FutureChem, Seoul, Korea) or RSV (10 mg/kg/day; Cayman Chemical, Ann Arbor, MI, USA) was administered through oral gavage for 4 weeks. Body weights and food intake were measured daily. Behaviors and activities were monitored. After a 4-week treatment period, the mice were sacrificed after overnight fasting, and blood and tissues were harvested and stored until analysis.
Determination of blood metabolic parameters
An oral glucose tolerance test (OGTT) was performed 2 days before the final day of treatment. After overnight fasting, glucose solutions were given through stomach gavage (2 g/kg body weight). Blood glucose concentrations were measured at 0, 15, 30, 60, 90, and 120 min after the glucose challenge using a Glucocard X-Meter (Arkray, Kyoto, Japan). At the end of the treatment period, blood was collected through cardiac puncture. After centrifugation, the supernatant serum was separated for further analysis. Fasting serum glucose and insulin concentrations were analyzed using commercial kits from Sigma-Aldrich (St. Louis, MO, USA) and Crystal Chem (Downers Grove, IL, USA), respectively. The homeostasis model assessment of insulin resistance (HOMA-IR) and HOMA β-cell were calculated as follows: HOMA-IR = fasting insulin (μU/mL) × fasting glucose (mmol/L)/22.5; HOMA β-cell = (20 × fasting insulin [μIU/mL])/(fasting glucose [mmol/L] −3.5).
Determination of oxidative stress markers
Liver tissues were homogenized in cold buffer (pH 7.4). Then, the mixture was centrifuged at 9520 g for 10 min at 4°C, and the supernatant was obtained for further biochemical analyses. The levels of malondialdehyde (MDA), superoxide dismutase (SOD) activities, and catalase activities were determined using commercial reagent kits based on the manufacturer's recommendations (Cayman Chemical).
RNA extraction and real-time polymerase chain reaction
Total RNA was extracted from liver tissues using the PureLink RNA Mini Kit (Ambion, Waltham, MA, USA). Complementary DNA (cDNA) was synthesized from extracted RNA using the High-Capacity RNA-to-cDNA Kit (Applied Biosystems, Waltham, MA, USA). Gene expression was analyzed by real-time polymerase chain reaction (PCR; StepOnePlus; Life Technologies, Carlsbad, CA, USA) by mixing SYBR Green (Power SYBR Green PCR Master Mix; Applied Biosystems) and gene-specific primers (Bioneer, Daejeon, Korea). The conditions used were as follows: 95°C for 10 min, followed by 40 cycles of denaturation (95°C for 10 sec), annealing (Tm [°C] for 15 sec), and extension (72°C for 60 sec). The cycle number for threshold of detection was determined using StepOne software (Life Technologies). Messenger RNA (mRNA) expression of each target was normalized to that of the 18S gene and expressed as fold change relative to controls.
Western blot analyses
Liver tissues were prepared, and Western blot analyses were performed as described previously. 38 Liver tissues were lysed using a cold lysis buffer containing protease and phosphatase inhibitors (Santa Cruz Biotechnology, Santa Cruz, CA, USA). The lysates were centrifuged, and supernatants were collected. Western blotting analysis was performed by denaturing 25 μg of protein at 95°C for 5 min in a Laemmli sample buffer and 2-mercaptoethanol. Proteins were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto PVDF membranes (Millipore, Billerica, MA, USA). The membranes were blocked and incubated with antibodies against beta-actin, catalase, glutathione peroxidase (GPx), NAD(P)H:quinone oxidoreductase 1 (NQO1), and tumor necrosis factor (TNF)-alpha (Cell Signaling Technology, Danvers, MA, USA). Membranes were exposed to the horseradish peroxidase-conjugated secondary antibody (Santa Cruz Biotechnology). Signals were developed using the ECL luminol reagent (Santa Cruz Biotechnology), recorded, and quantified with the Syngene G box (Syngene, Cambridge, United Kingdom).
Statistical analysis
Statistical analysis was performed using SPSS statistics 24 (IBM Corp., Armonk, NY, USA). Data are expressed as mean ± standard error of the mean. Differences among experimental groups were analyzed using one-way analysis of variance, followed by Bonferroni's post hoc test, where P < .05 was considered significant.
Results
Effects of PIC and RSV on body weight, food intake, and glucose control in HF diet-fed mice
HF feeding for 4 weeks increased body weight and weights of epididymal and subcutaneous fat pads compared with the control group, confirming that HF feeding induced obesity in mice (Table 1). Oral administration of PIC and RSV (10 mg/kg/day) for 4 weeks did not affect body weight, food intake, and weights of liver, epididymal, and subcutaneous fat pads (Table 1).
Effect of Piceatannol and Resveratrol on Body Weight, Food Intake, and Tissue Weight
Data are expressed as mean ± SEM (n = 8–12 per group). Different letters within a variable are significantly different at P < .05.
CON, control; HF, high-fat diet control; PIC, piceatannol; RSV, resveratrol; SEM, standard error of the mean.
Fasting serum glucose and the AUC during the 2-h OGTT increased upon HF feeding (Table 2). Mice treated with PIC and RSV showed improved glucose control based on lower levels of AUC during the OGTT compared with HF groups (Table 2). However, fasting serum insulin concentrations, HOMA-IR, and HOMA-beta were not significantly affected by HF diet, PIC, and RSV treatments. PIC-mediated improvement in glycemic control occurred along with increased hepatic gene expression levels of insulin receptor (IR) and AMP-activated protein kinase (AMPK) (Fig. 1). RSV treatment tended to increase gene expression of IR (55.86%) and AMPK (231.33%) in livers of HF-fed mice; however, the increases were not statistically significant (Fig. 1).

Effects of PIC and RSV on gene expression levels of glucose control-related targets.
Effect of Piceatannol and Resveratrol on Glucose Control
Data are expressed as mean ± SEM (n = 8–12 per group). Different letters within a variable are significantly different at P < .05.
AUC, area under the curve; HOMA-IR, homeostasis model assessment of insulin resistance; OGTT, oral glucose tolerance test.
PIC upregulates hepatic gene expression levels of sirtuins and downstream targets in HF diet-fed mice
The effects of PIC and RSV on hepatic gene expression levels of sirtuins and their downstream targets are shown in Figure 2. Four weeks of HF feeding significantly decreased Sirt1 gene expression in the liver, and PIC treatment normalized the levels (Fig. 2A). Furthermore, PIC upregulated the hepatic gene expression of other isotypes of Sirt, Sirt3 and Sirt6 (Fig. 2B, C), which are considered to regulate metabolism especially in mitochondria and the nucleus, respectively. The increases in Sirt1, Sirt3, and Sirt6 were accompanied by concomitant upregulation of two downstream targets of Sirt, peroxisome proliferator-activated receptor gamma coactivator 1 (PGC1)-alpha and foxo1, in HF diet-fed mice (Fig. 2D, E). Moreover, similar doses of PIC exerted more potent effects on hepatic expression of Sirt1 and Sirt6 compared with RSV-treated mice (Fig. 2A, C).
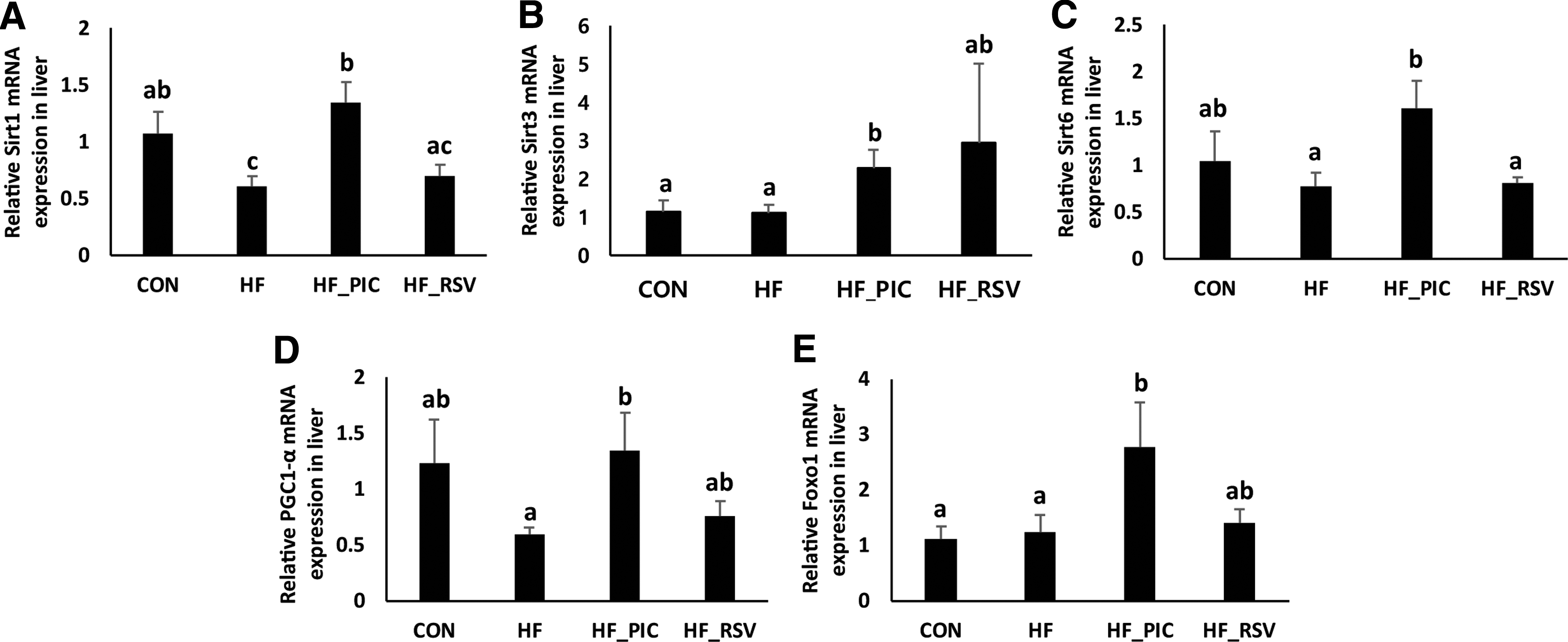
Effects of PIC and RSV on sirtuins and their downstream targets.
PIC and RSV suppress levels of inflammatory markers in livers of HF diet-fed mice
Hepatic protein levels of a proinflammatory marker, TNF-alpha, increased with the HF diet (Fig. 3C, D). The hepatic inflammatory markers, including interleukin (IL)-1 and IL-6, were downregulated by 4 weeks of RSV treatment (Fig. 3A, B). PIC and RSV treatments significantly lowered the hepatic protein levels of TNF-alpha in HF diet-fed mice (Fig. 3C, D), and the PIC-mediated effects on TNF-alpha were significantly stronger compared with those by RSV in livers of HF diet-fed mice.
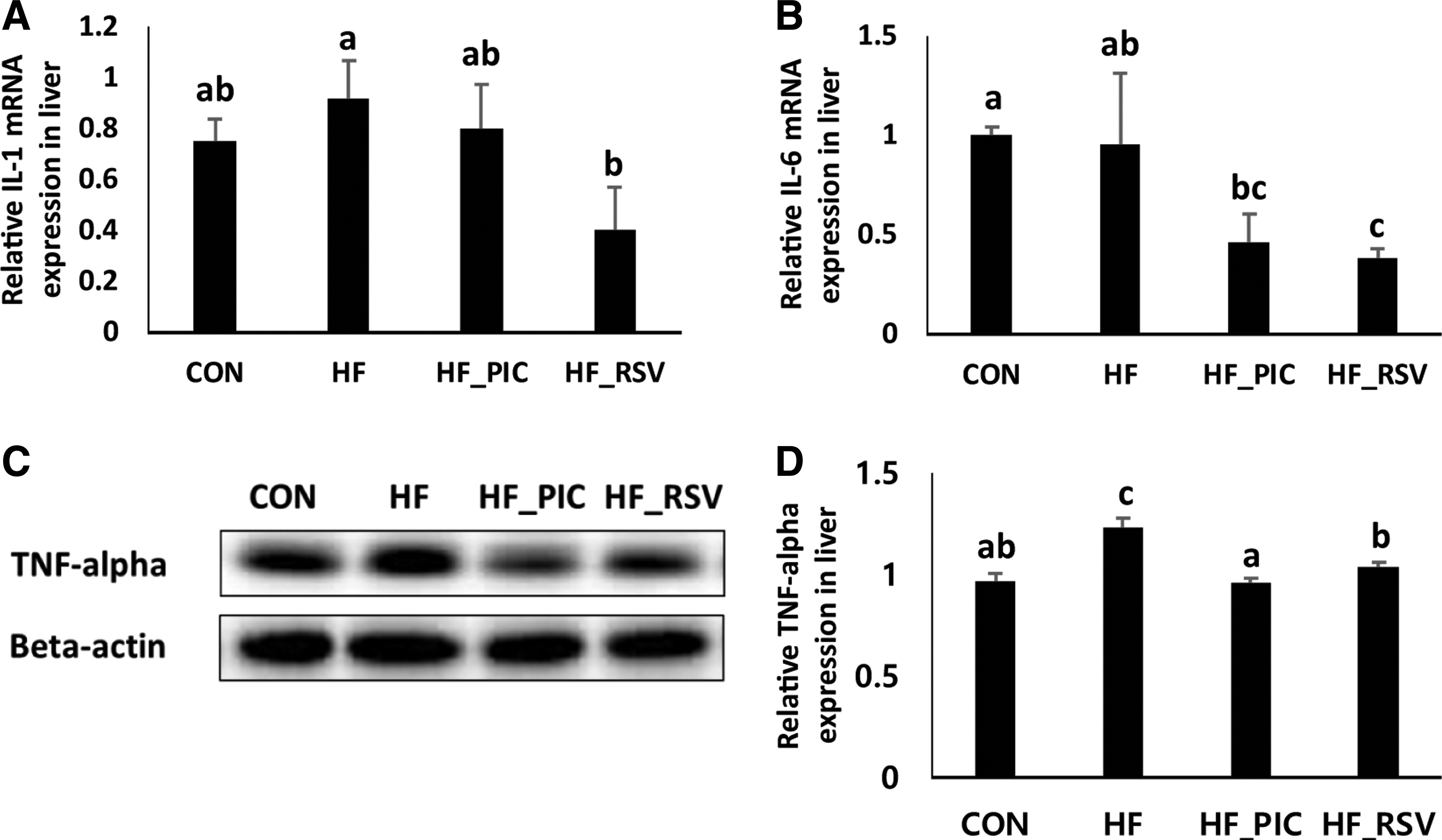
Effects of PIC and RSV on hepatic inflammation in mice. Gene expression levels of
RSV increases protein levels of NQO1 in livers of HF diet-fed mice
Inflammation and oxidative stress affect each other in various experimental conditions. Therefore, we analyzed several markers (MDA and enzymatic activities of SOD and catalase) of oxidative stress. However, PIC and RSV treatments resulted in minimal effects on oxidative stress-related markers in livers (Table 3). The levels of hepatic MDA and activities of SOD were not affected. The HF diet significantly reduced hepatic activities of catalase, which were similar between HF_PIC and HF_RSV groups. Moreover, protein levels of antioxidant enzymes, NQO1, catalase, and GPx, were analyzed by Western blot (Fig. 4). Among the markers analyzed, the HF diet increased protein levels of antioxidant enzymes, NQO1 and GPx, but not catalase (Fig. 4). NQO1 expression increased only in livers of RSV-treated mice compared with HF control mice (Fig. 4B).
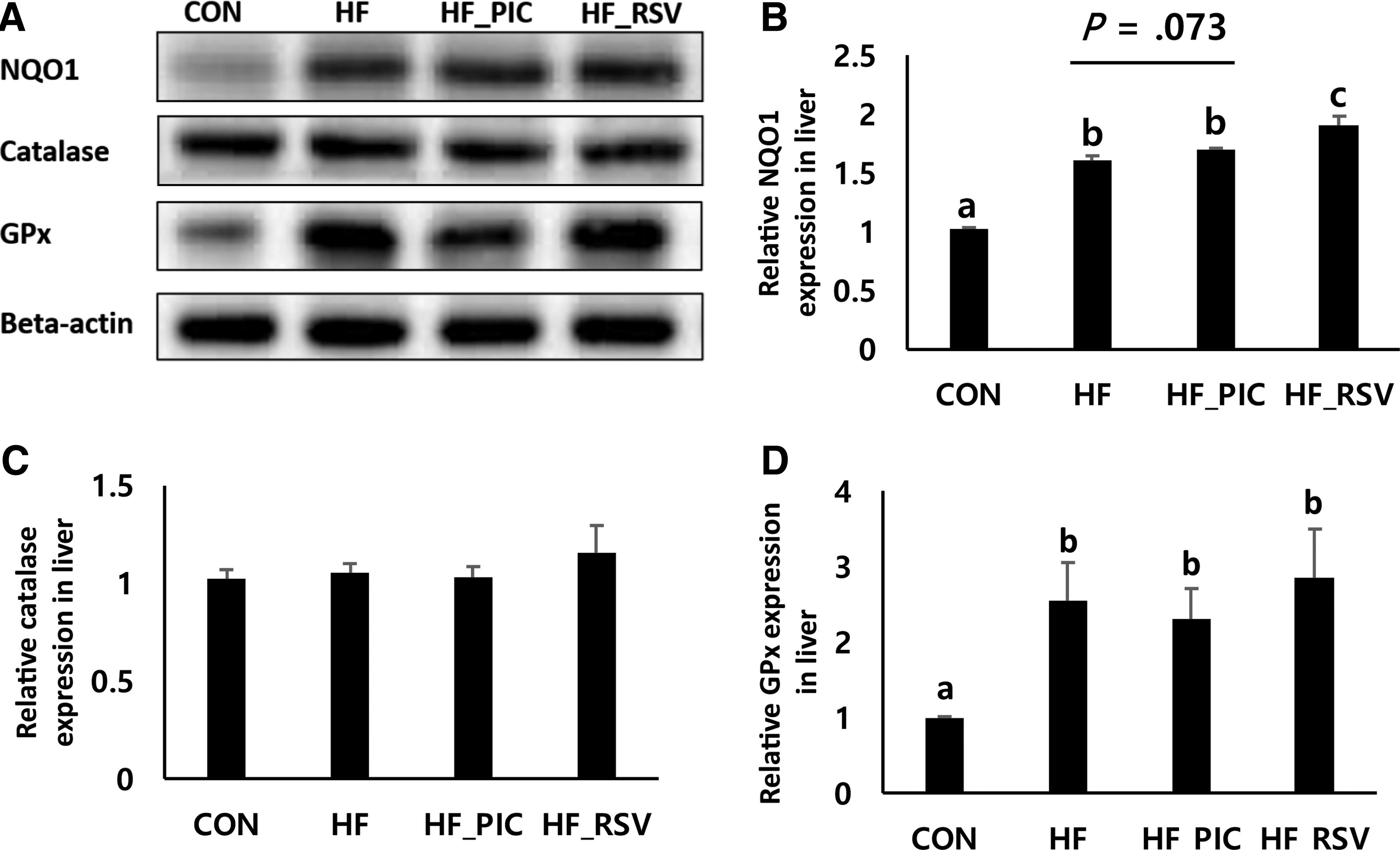
Effects of PIC and RSV on oxidative stress-related targets.
Effects of Piceatannol and Resveratrol on Oxidative Stress in Livers
Data are expressed as mean ± SEM (n = 8–12 per group). Different letters within a variable are significantly different at P < .05.
CAT, catalase; MDA, malondialdehyde; SOD, superoxide dismutase.
Discussion
Our findings document beneficial effects of PIC and RSV on various markers of glucose control, sirtuins, inflammation, and oxidative stress in HF diet-fed mice. HF feeding increased body weight and weights of major fat pads and impaired glucose control, inflammation, and oxidative stress. The beneficial effects of PIC on hepatic markers were generally stronger in HF diet-fed mice compared with those exposed to RSV. Specifically, PIC showed greater alterations of sirtuins and their downstream targets, PGC1-alpha and foxo1, and a proinflammatory marker, TNF-alpha, as well as an antioxidant enzyme, NQO1, warranting additional investigation into the role of RSV and its derivatives, including PIC, in regulation of sirtuins and their metabolic pathways.
Treatment dosage was based on previous experiments and a literature review. 39 –42 Preliminary experiments of PIC and RSV treatments up to 15 mg/kg/day for 28 days through oral feeding or subcutaneous perfusion using an osmotic pump did not yield any negative signs or symptoms. Therefore, 10 mg/kg/day was used in the current experiments, considering the solubility and volume of treated solutions. Four weeks of PIC and RSV treatments did not cause any abnormal alterations in body weights and behaviors/activities as well or unanticipated changes.
PIC exerts beneficial effects in rodent models of prostate cancer, type 2 diabetes, and diet-induced obesity, as well as overweight men. 11,13,15 RSV also improves weight control, brown adipose tissue formation, and amyloid beta (Aβ) accumulation, as well as Aβ-related pathological changes. 17,18,20 –22 These biological activities suggest that RSV and its derivative PIC may have regulatory effects on obesity and aging-related metabolism, mediated through several metabolic pathways, including sirtuins and inflammation, as well as oxidative stress.
In our study, glycemic control during the 2-h OGTT was enhanced by PIC and RSV treatments. Maintenance of glycemic control requires a well-orchestrated endocrine mechanism controlling glucose metabolism and related cellular signaling. The feedback loop of insulin and glucagon directly responds to internal and external stimuli, and indirectly, various cytokines, including adiponectin and TNF-alpha, affect insulin signaling. 43 PIC and RSV treatments significantly reduced the 2-h OGTT AUC, accompanied by increased hepatic expression of IR and AMPK especially in PIC-treated mice. Similar effects occurred in a diet-induced obese mouse model and overweight men. 15,44 These reports suggest that improved glycemic control by PIC treatment correlates with the action of insulin signaling molecules, including IR, and activation of AMPK, as well as inhibition of α-glucosidase activity. 15,44
Sirtuins act as NAD+-dependent deacetylase and ADP-ribosyltransferase, which are parts of working mechanisms of Sirt-mediated effects. 45 Sirt1 is located in the cytosol and nucleus, regulating inflammation and glucose/lipid metabolism. 46 Involvement of Sirt1 in AMPK activation was revealed from the whole-body knockout mouse model of Sirt1, and RSV improved metabolism and mitochondrial function in vitro and in vivo through Sirt1-AMPK signaling. 47 Sirt3 is mainly located in mitochondria as well as, in part, in the cytosol. Initial reports on Sirt3 were focused on mitochondrial biogenesis and functions; however, recently, its diverse roles in iron/glucose/lipid metabolism, cell cycle/death, insulin resistance, cancer, cardiovascular diseases, and aging were also demonstrated. 48 –50 Sirt6 is expressed in the cytosol and nucleus and is known to regulate nutrient metabolism and inflammation. 51,52 Its actions are often overlapped with those of Sirt1; Sirt1 and Sirt6 are considered as cooperative and backup regulatory components through reciprocal action. Previous reports on RSV and Sirt demonstrated that RSV increased expression and activities of Sirt1; however, research evidence on other sirtuins is limited. In the current study, we found that PIC has a dramatic effect on sirtuin gene expression, especially Sirt1, Sirt3, and Sirt6. On the other hand, similar doses of RSV had no effect on sirtuins and their downstream targets (PGC1-alpha and foxo1). Based on these results, PIC may have a higher biological activity than RSV regarding regulation of Sirt. However, the superior efficacy of PIC compared with RSV is not fully apparent and may be due, in part, to structural differences associated with the additional OH group. Findings regarding the RSV effects on sirtuins and their downstream targets in the current study were inconsistent with previous reports. 40 The different results involving sirtuins may arise from the altered treatment route (oral gavage versus subcutaneous osmotic pump implantation). In the current experiment, oral gavage feeding delivered 10 mg/kg/day RSV, which was not fully available following digestion and absorption through the gastrointestinal tract. However, subcutaneous perfusion through the osmotic pump is involuntary, guarantees continuous exposure to the infused RSV during the entire experimental period, and yields greater metabolic effects compared with oral feeding.
Sirtuins regulate inflammation responses in various experimental models. Sirt1 overexpression protects against hepatotoxicity by alleviating inflammation and oxidative stress in mice and hepatocytes. 53 Excessive proinflammatory responses exacerbate oxidative stress; increased levels of proinflammatory markers, TNF-alpha, IL-1, and IL-6, disrupt the balance between oxidative species and antioxidative enzymes. Inflammation may increase MDA concentrations and reduce activities and expression levels of antioxidative enzymes, SOD, catalase, NQO1, and GPx. In the current study, PIC and RSV effectively suppressed the TNF-alpha level, and RSV reduced the expression of additional proinflammatory markers, IL-1 and IL-6. Moreover, RSV exhibited superior regulation of antioxidative enzyme NQO1. As expected, findings from the current study demonstrated the close relationship between inflammation and oxidative stress through sirtuins.
In conclusion, PIC was superior to an equal concentration of RSV in regulating Sirt and its downstream targets as well as insulin signaling-related parameters, while RSV potentially suppressed the levels of proinflammatory markers and increased NQO1 protein levels. Although the regulatory effects of RSV on metabolism were not entirely consistent with previous reports, it is evident that oral gavage feeding is more physiological compared with subcutaneous infusion through the osmotic pump. Current evidence regarding the superior efficacy of PIC on sirtuins and glucose control/insulin signaling-related parameters as well as the regulatory effect of RSV on inflammation and oxidative stress provides additional insights into the role of RSV and its analogs in metabolic health.
Footnotes
Acknowledgments
This work was supported by a research grant from the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2017R1D1A1B03030617). The funder had no role in study design; data collection, analysis, and interpretation; decision to publish; or preparation of the manuscript.
Author Disclosure Statement
There are no conflicts of interest.