
Abstract
Intestinal enterochromaffin (EC) cell hyperplasia and increased 5-hydroxytryptamine (5-HT) availability play key roles in the pathogenesis of abdominal hypersensitivity of irritable bowel syndrome (IBS). This study aims to study the effect of quercetin on visceral pain and 5-HT availability in postinflammatory IBS (PI-IBS) rats. PI-IBS model rats were administered quercetin by gavage at doses of 5, 10, and 20 mg/kg for 14 days. Compared with normal rats, the visceral pain threshold of PI-IBS rats was markedly decreased and the abdominal motor response to colon distension was markedly increased. The EC cell count and 5-HT level, as well as tryptophan hydroxylase (TPH) protein, were all significantly elevated in PI-IBS rats, while the 5-HT reuptake transporter (serotonin transporter) was reduced. Genes that are responsible for enteroendocrine cell differentiation, that is, Ngn3 and pdx1, were significantly increased in the PI-IBS group. Quercetin treatment markedly elevated the pain threshold pressure and decreased the visceral motor response of PI-IBS animals; and EC cell density and 5-HT level, as well as TPH expression, in the PI-IBS group were all reduced by quercetin. Quercetin treatment also significantly reduced colonic expression of Ngn3 and pdx1 of PI-IBS. Findings from the present study indicated that the analgesic effect of quercetin on PI-IBS may result from reduction of 5-HT availability in the colon, and the regulatory role of quercetin in endocrine progenitors may contribute to reduced EC cells.
Introduction
Postinflammatory irritable bowel syndrome (PI-IBS), a subset of IBS, is featured by symptoms (visceral pain, bloating, and changed bowel habits) that occur post initial episode of intestinal infection. 1 Nowadays, the pathogenesis of PI-IBS is relatively unclear, and therapies for PI-IBS are also unsatisfactory. Abdominal pain is a major symptom of IBS 2 ; elevated intestinal enterochromaffin (EC) cells and increased 5-hydroxytryptamine (5-HT) availability were found to be involved in the pathogenesis of abdominal pain. 3
Most of the body's 5-HT is located within the gut with about 90% in EC cells; tryptophan hydroxylase (TPH) controls 5-HT synthesis. 5-HT is released upon stimulation, and the released 5-HT activates multiple receptors expressed in nociceptive afferents and regulates gastrointestinal sensitivity. 4 After release, 5-HT is rapidly transported back to the enterocytes by the 5-HT reuptake transporter (serotonin transporter [SERT]). It is found that EC cell hyperplasia is always followed by elevated 5-HT availability, the increased 5-HT availability results in motility dysfunction and visceral hyperplasia in IBS. 5 Knowing the critical status of 5-HT in the development of abdominal pain, altering 5-HT bioavailability is proposed to be a promising target for IBS treatment. 6 EC cells are key cells that are responsible for 5-HT biosynthesis, storage, and release in the gut, thus EC cell density and/or 5-HT release occupy a very important place in 5-HT availability. EC cells originate from intestinal stem cells, 7 and intestinal stem cells divide and differentiate into secretory lineage cells (i.e., goblet cells, enteroendocrine cells, and Paneth cells). 8 Currently, the pathogenesis mechanism in the elevated EC cell count is found to be related to altered genes, diet, gut flora, and low-grade inflammation. It is proposed that the increased EC cells might arise from dysregulation of the clonogenic or polarization of the stem cell in the gut. 9,10 A previous study reported that altered differentiation of intestinal epithelial cells may result in EC cell hyperplasia, 11 indicating that EC cell hyperplasia may come from differential alteration of intestinal epithelial cells.
Quercetin, a subclass of flavonoids, is widely distributed in a variety of foods. 12 Quercetin had multiple pharmacological activities, such as anti-inflammatory, anticancer, and analgesic effects. 13 It is found that quercetin can attenuate formalin-induced pain-like behavior and thermal hypernociception in rodents, 14,15 but the analgesic mechanism has not been fully identified. A recent study showed that quercetin can modulate gut microbiota, 16 and intestinal microbiota have been known as key regulatory factors related to 5-HT biosynthesis. 17 It is also found that dietary quercetin can suppress intestinal stem cell proliferation, 18 suggesting that quercetin may have an effect on intestinal 5-HT availability.
In this present study, the pharmacological activity of quercetin on visceral hypersensitivity was assessed in the PI-IBS rat model, and we further investigated its effect on EC cell count and 5-HT availability in the gut. Moreover, the underlying mechanism of quercetin was also explored by evaluating the protein responsible for epithelial cell proliferation and the gene for development of endocrine progenitors.
Materials and Methods
Animals
Male Wistar rats (8 weeks of age, 250 ± 20 g) were purchased from the Animal Center of Gansu University of Tradition Chinese Medicine. Animals were housed in plastic cages at 25°C under a light–dark cycle with free access to diet. Animals were raised for about 7 days before the study for acclimatization. All the researchers followed guidelines of the Committee on Use of Animal Subjects in Teaching and Research of the First Hospital of Lanzhou University (LDYYLL2014-0028).
Induction of the animal model
The animal model was induced based on our previous report. 19,20 In brief, animals were narcotized by intraperitoneal injection of chloralic hydras (350 mg/kg body weight). A thin tube was inserted in the distal colon, and 2,4,6,-trinitrobenzenesulfonic acid (TNBS) solution (5 mg in 0.8 mL with 50% alcohol) was infused per rat. Rats in the control were infused with saline. After 30 days post-TNBS, abdominal pain thresholds were tested and the animals with abdominal hypersensitivity (pain threshold under 30 mmHg) were determined to be PI-IBS animals.
Group setting and sample collection
This study consists of two series of experiments. In the first study, PI-IBS animals were randomly grouped (n = 12). Rats were orally given 0.3% carboxymethyl cellulose sodium salt (CMC-Na, Group 2) or quercetin suspension (95% purity; Sigma-Aldrich, in 0.3% CMC-Na) at doses of 5, 10, and 20 mg/kg (Groups 3–5). The dosage of quercetin was set based on the result of our preliminary experiment. Normal rats in the control were orally treated with 0.3% CMC-Na (Group 1). Quercetin was orally given to rats once daily for two consecutive weeks. A total of six animals per group were selected for pain measurement by an abdominal withdrawal reflex (AWR) study, while the remaining animals were used for electromyography (EMG) recording. The impact of quercetin on intestinal 5-HT availability was tested in the other series of the study. Five groups were used, the grouping and drug treatment were the same as that in the first study, rats in each group (n = 6) were used for obtaining the data without colorectal distension (CRD). A 6-cm-long colon tissue was collected and cut into three parts, one part was put in 4% paraformaldehyde for histological examination; the middle one was preserved for 5-HT measurement; and the left was used for Western blot and real-time PCR analyses.
AWR test and EMG recording
AWR study was applied to measure the visceral pain threshold pressure. 21 In brief, animals were anesthetized using diethyl ether so that a balloon could be inserted into the descending colon. CRD was initiated with increases of 5 mmHg until obvious abdomen contraction was observed, and the threshold of CRD elicited was recorded and repeated five times. The data from each rat were averaged. The method for EMG recording was conducted as previously reported. 22,23 An implantation operation was performed to put electrodes in the external oblique muscle before the EMG study. After drug treatment, rats were subjected to CRD (20–80 mmHg; 2-min interstimulus intervals) for a total of three times. The alteration of area under the curve was calculated in the period of 20-s baselines.
5-HT content assay
5-HT concentration of the colon was detected according to the established procedure. 24 In brief, colon tissue homogenates were made using 15% trichloroacetic acid, and the homogenate was then centrifuged at 10,000 g for 10 min. Supernatants were collected and passed through a 0.22-μm filter, and samples were extracted by adding ether five times. Derivatization solution was added into the obtained samples and then incubated at 25°C for half an hour. The 5-HT concentration was detected by using a capillary electrophoresis apparatus with a laser-induced fluorescence detector. The results were presented as ng/mg tissue weight.
Western blot assay
Colon samples were homogenized to obtain the protein extract. The protein content was determined and then denatured for 5 min. Samples were separated using electrophoresis in 8–10% polyacrylamide gel electrophoresis gel, then transferred to a membrane (Bio-Rad). After blocking in 5% nonfat milk, the membrane was then incubated with primary antibodies (anti-SERT, 1:500; Calbiochem; anti-TPH, 1:1000; Santa Cruz; antiproliferation cell nuclear antigen [PCNA], 1:1000; Abcam; Anti-Ngn3, 1:500; Abcam; Anti-Pdx1, 1:500; Abcam; Anti-Pax6, 1:1000; Abcam; antiactin, 1:2000; Cell Signaling; and anti-GAPDH, 1:2000; Sigma) overnight at 4°C. After incubation with secondary antibodies, enhanced chemiluminescence reagents (Amersham) were added, and the immunoreaction was evaluated by the optical density assay.
Immunohistochemistry
Paraffin sections were prepared by dewaxing and antigen retrieval and then incubated with primary antibody (anti-5-HT, 1:4000; Sigma-Aldrich; and anti-PCNA, 1:500; Abcam) at 4°C. After rinsing, the sections were labeled with streptavidin–biotin (LSAB HRP kit; Dako). A total of five random fields at 200 × or 400 × magnification per section were observed using ImageJ (Wayne Rasband; NIH) by an investigator blinded to group settings. EC cell density was expressed as number/mm 2 of mucosa.
Silver staining
Slides were incubated with 5% Tollens’ reagent overnight at room temperature. After rinsing thoroughly in water, 5% hyposulfite solution was added on the slides for 5 min of incubation at 25°C. A total of five random fields captured at 200 × magnification were counted. ImageJ software was used to quantify the cell number, and the density of enteroendocrine cells was presented as No./mm 2 of mucosa.
Real-time PCR
Colon samples were prepared in TRIzol reagent (Invitrogen) to obtain total RNA extract. Measurement was performed by applying the real-time fluorescence detection method with a Power SYBR Green PCR Master Mix (Applied Biosystems). All reactions were normalized to GAPDH. Primers for target genes are described in Table 1. Quantitative comparison was obtained through the ΔΔCT. The final results were adjusted by setting the mean mRNA level as 1 in the control.
Primer Sequences Used for Real-Time Polymerase Chain Reaction
F, forward; R, reverse.
Statistical analysis
Data are shown as mean ± standard deviation. To compare data from multiple groups, data were analyzed using one-way analysis of variance, followed by a Student–Newman–Keuls test with SPSS 16.0. Differences were considered statistically significant when the P value was below .05.
Results
Quercetin attenuated visceral hypersensitivity in the PI-IBS model
The pain threshold of PI-IBS animals was markedly lowered when compared with the normal animal (31 ± 3 vs. 39 ± 3, P < .05, Fig. 1A); quercetin administration at high and median dosages significantly elevated the pain threshold of PI-IBS animals by 38 ± 2 and 34 ± 2, respectively. Results from the EMG test (Fig. 1B, C) also revealed that the abdominal motor response to CRD was significantly increased in PI-IBS animals, and high and median doses of quercetin administration markedly decreased the visceral motor responses of PI-IBS animals (P < .05).
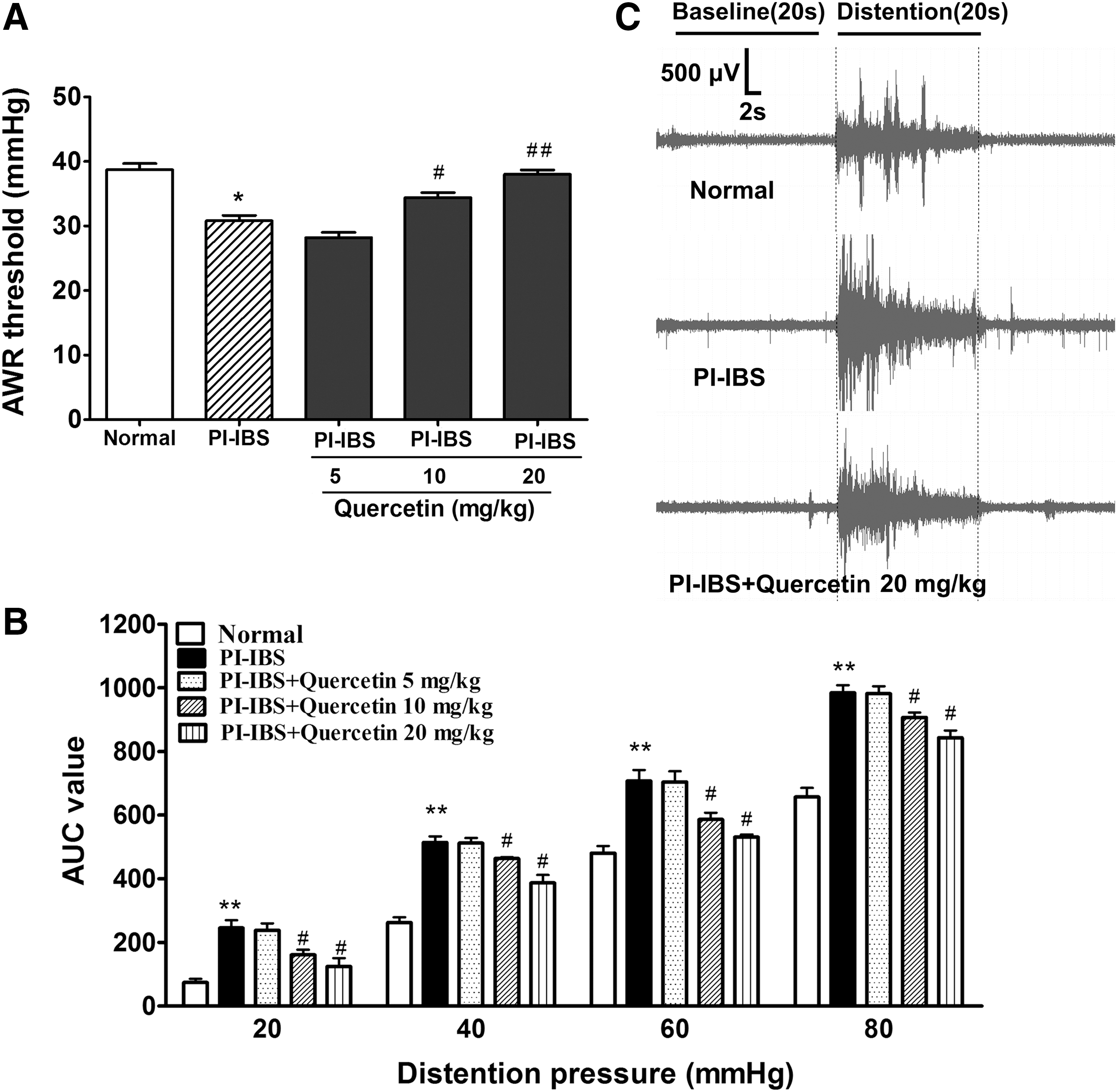
Effects of quercetin on visceral hypersensitivity in PI-IBS rats.
Quercetin reduced 5-HT content and 5-HT synthesis in the PI-IBS model
5-HT concentrations in colons of PI-IBS animals were significantly increased by 34% (Fig. 2A), and quercetin administration dose-dependently lowered the colonic 5-HT concentration (15–33%). TPH protein expressed in the PI-IBS model was elevated by 31% when compared with the normal animal (P < .05, Fig. 2B), the high dose of quercetin administration significantly reduced TPH expression by 23%. As shown in Figure 2C, SERT expression of PI-IBS animals was substantially reduced (P < .01), and high-dose quercetin administration did not have a significant impact on SERT protein of the PI-IBS animal.

Effects of quercetin on colonic 5-HT availability in PI-IBS rats.
Quercetin reduced colonic EC cell and endocrine cell density in the PI-IBS model
The EC cell density was markedly elevated by 60% in the PI-IBS model when compared with the normal animal (P < .05, Fig. 3A, B); high and medium dosages of quercetin markedly decreased the EC cell density by 28% and 21%, respectively (P < .05). The number of enteroendocrine cells was also significantly elevated by 48% in the PI-IBS group (Fig. 3C, D), and high and medium dosages of quercetin treatment significantly decreased the count of enteroendocrine cells in the PI-IBS model by 42% and 18%, respectively.
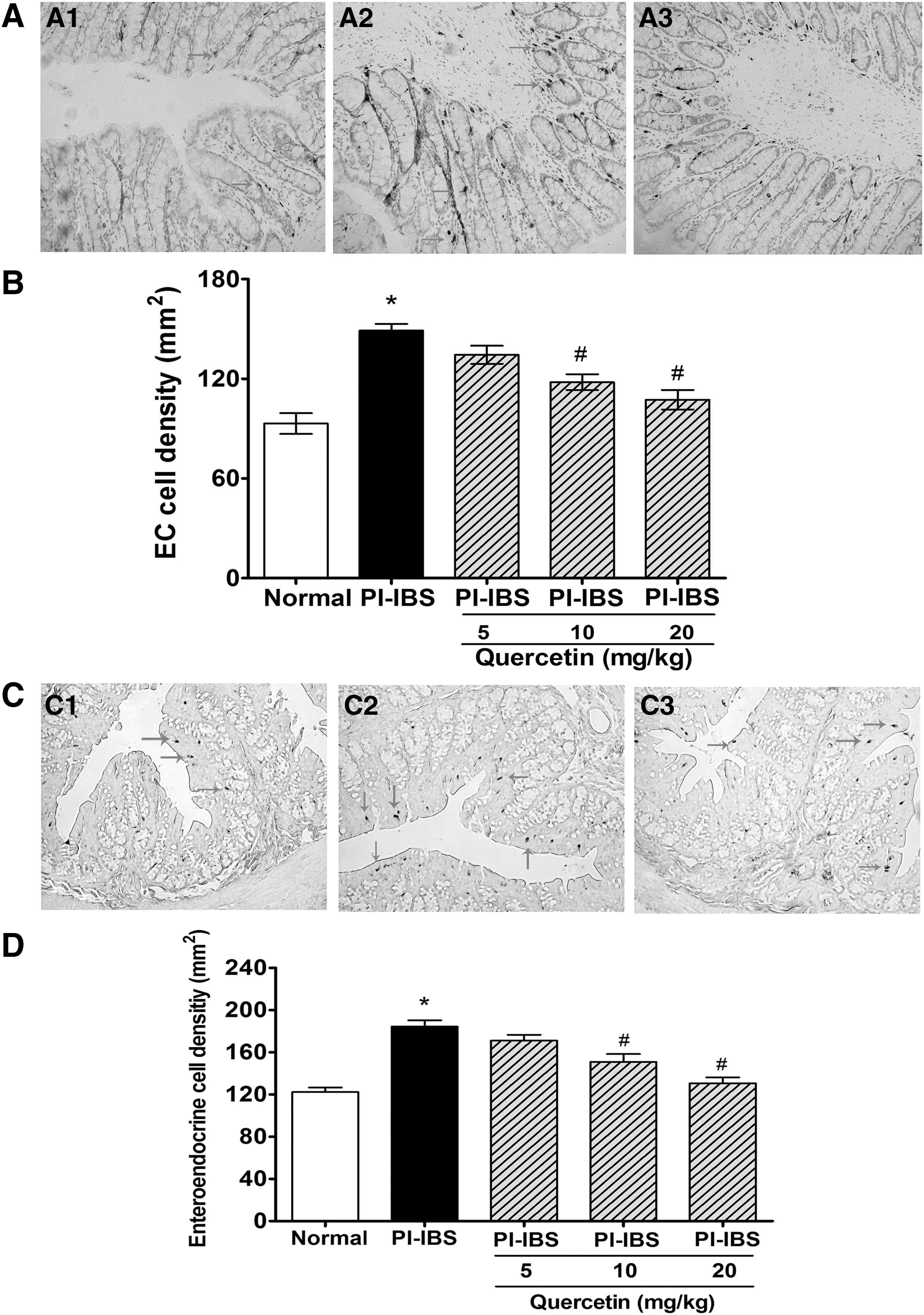
Effects of quercetin on the density of EC cells and enteroendocrine cells in the colon of PI-IBS rats.
Quercetin regulated proliferation of epithelial cells in the PI-IBS model
No significant difference was found in the protein level of intestinal PCNA among the normal animal, PI-IBS model, and high-dose quercetin-treated animal (Fig. 4), indicating that there was no alteration in proliferation of intestinal cells among the control group, PI-IBS group, and quercetin-treated group.

Effects of quercetin on mucosal PCNA expression in the colon of PI-IBS rats.
Quercetin regulated the gene expression of colonic endocrine cell development
As shown in Figure 5, data from the PCR test showed that the genes of Ngn3 and Pdx1, but not Pax6, were markedly increased in the PI-IBS model when compared with the normal animal (P < .05). High dose of quercetin administration significantly reduced mRNA levels of Ngn3 and Pdx1 of the PI-IBS animal, but had no effect on Pax6 expression. Data from Western blot analysis also revealed that proteins of Ngn3 and Pdx1 were both elevated in the PI-IBS model (P < .05) and Ngn3 and Pdx1 were reduced after quercetin treatment (P < .05).

Effects of quercetin on the expression of transcription factors that control differentiation of intestinal enteroendocrine cells in the colon tissue of the PI-IBS model.
Discussion
5-HT is one of the key neurotransmitters and paracrine passengers in the body; it has been linked to regulation or modulation of several symptoms of IBS. 25 In this present work, we first demonstrated that quercetin can lower abdominal hypersensitivity of PI-IBS animals by decreasing EC cell counts and 5-HT availability in the colon, and this effect may have close correlation with the regulatory role in epithelial secretory cell types in the gut (Fig. 6).
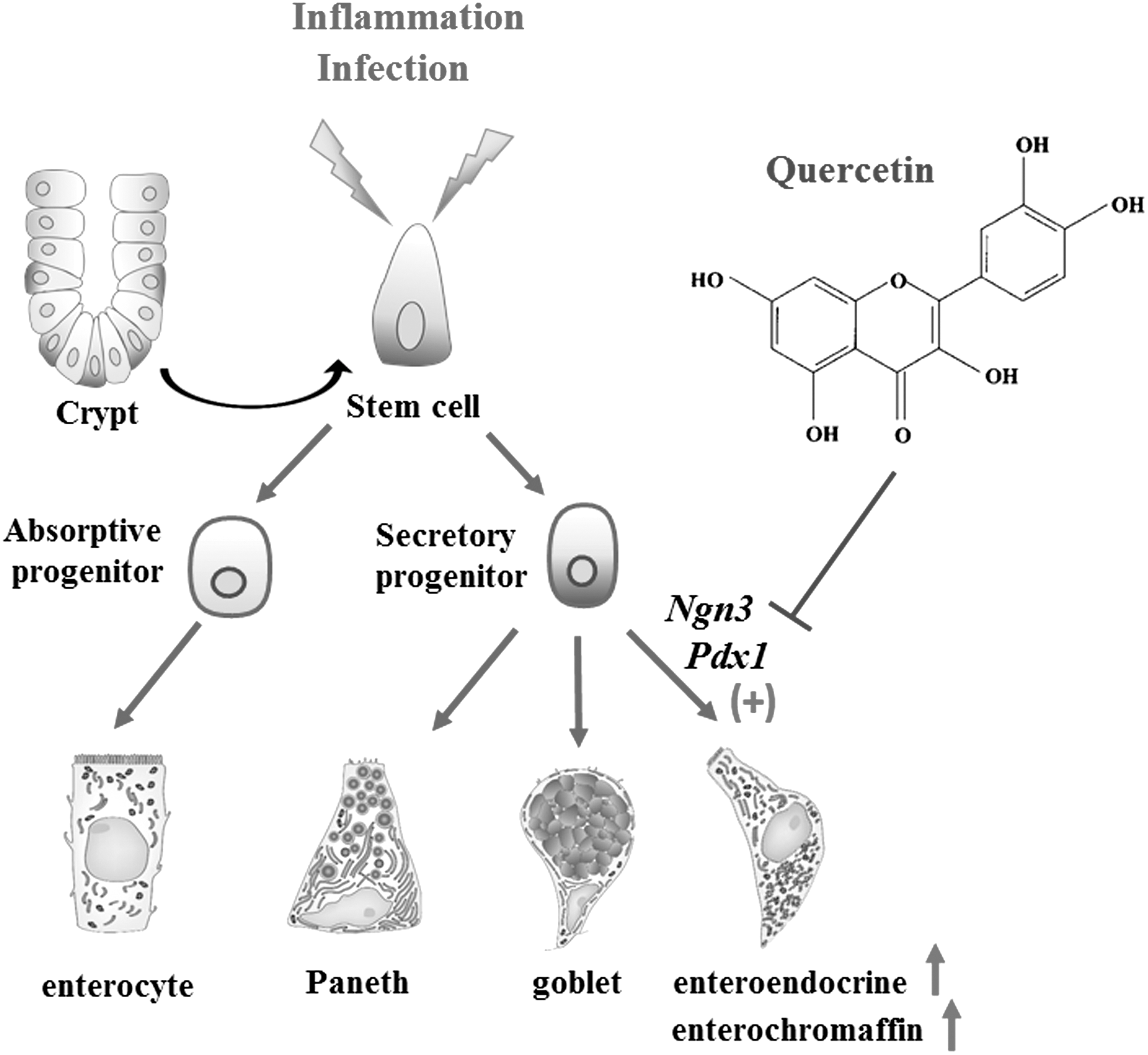
A proposed regulatory effect of quercetin on enteroendocrine cells and EC cells in the colon of PI-IBS rats. Intestinal stem cells divide and produce proliferating populations and then differentiate into enterocytes, goblet cells, Paneth cells, and enteroendocrine cells. EC cell is the main cell type of enteroendocrine cells; genes Ngn3, Pax6, and pdx1 of the Notch signaling pathway are responsible for differentiation of enteroendocrine cells in the gut. In PI-IBS rats, proliferation of stem cells is unchanged, but Ngn3 and pdx1 are upregulated, which finally lead to increase of enteroendocrine cells and EC cells in the colon. Quercetin administration downregulates the expression of Ngn3 and Pdx1 and thus reduces the density of intestinal enteroendocrine cells and EC cells of PI-IBS rats.
It is well known that elevated 5-HT availability is related to the pathogenesis of abdominal pain, and 5-HT synthesis, release, and reuptake can all affect intestinal 5-HT bioavailability. 2 Evidence of this study demonstrated that quercetin markedly relieved abdominal hypersensitivity in the PI-IBS model; the analgesic activity was consistent with the lowered 5-HT concentration of colon tissue, indicating that 5-HT concentration was closely correlated with visceral sensation. 26 We also found that quercetin treatment can reduce colonic EC cell densities and TPH expression of PI-IBS, which all contributed to reduction of colonic 5-HT bioavailability. The results also showed that SERT in the PI-IBS model was significantly reduced, which is in accordance with previous reports showing decreased SERT protein in IBS patients and an animal model. 27,28 However, quercetin administration showed no effect on the SERT protein level in PI-IBS animals. All of these results indicated that quercetin can alleviate splanchnic hypersensitivity of PI-IBS by decreasing colonic 5-HT availability, and the lowered colonic EC cell count may contribute to the decrease in 5-HT availability in the colon.
EC cells are the main enteroendocrine cells that are responsible for 5-HT biosynthesis, storage, and release into the gut 29 ; we further investigated the bioactivity of quercetin on intestinal endocrine cells of the colon. We found that the density of enteroendocrine cells of a PI-IBS model was also apparently elevated, and quercetin markedly reduced the count of enteroendocrine cells in a PI-IBS model. Based on these results, it seems that the increased EC cells may come from increase of enteroendocrine cells in the PI-IBS animal. These findings are in accordance with a previous study reporting that altered endocrine cells in postinfective IBS and increased EC cells, as well as endocrine cells, all contribute to clinical symptoms of PI-IBS. 10,30
Knowing that gastrointestinal epithelium is distinguished by its fast and persistent regeneration, the ability of proliferation or differentiation of epithelial cells plays a vital role in maintaining cellular homeostasis. 31 Based on the effect of quercetin on intestinal EC cells and enteroendocrine cells, we further explored the effect of quercetin on intestinal cell proliferation as well as the genes that control enteroendocrine cell development. We found that the expression of intestinal PCNA of PI-IBS animals was the same as that of the normal animal and quercetin showed no effect on PCNA. These findings indicated that proliferation of intestinal epithelial cells may be unchanged under PI-IBS conditions and quercetin has no effect on intestinal cell proliferation. However, the genes controlling intestinal enteroendocrine cell differentiation, that is, Ngn3 and pdx1, were all markedly increased in PI-IBS animals and quercetin treatment apparently reversed the alteration of Ngn3 and pdx1. These data were consistent with protein alteration, and results indicated that the altered development of enteroendocrine cells may contribute to EC cell hyperplasia in PI-IBS rats.
Quercetin has been found to have multifarious pharmacological activities. 13 Up to now, we do not know how quercetin exerts its regulatory effect on epithelial cell development. Recent studies reported that intestinal microbiota could adjust and control the proliferative and differential properties of stem cells in the gut 32 and quercetin can regulate intestinal microbiota. 33 It is possible that the effect of quercetin on intestinal endocrine cells and EC cells may come from its regulatory effect on intestinal microbiota. In view of the unclear mechanism about EC cell hyperplasia and the multiple bioactivities of quercetin, more studies are still needed in the future.
Conclusions
In this present work, we found that quercetin can relieve visceral hypersensitivity in the PI-IBS rat model by lowering intestinal EC cells and 5-HT availability; the molecular mechanism may have a correlation with the regulatory role of quercetin in development of intestinal endocrine cells and EC cells. These findings may provide preliminary data for the analgesic usage of quercetin in IBS, and quercetin may have the potential to be used for EC cell hyperplasia.
Footnotes
Acknowledgments
This work was supported by the National Science Foundation of China (81341145 and 81400596); the Natural Science Foundation of Gansu Province (grant number 1606RJZA117), and the Fundamental Research Funds for the Central Universities (lzujbky-2014-223 and lzujbky-2015-38).
Author Disclosure Statement
No competing financial interests exist.