
Abstract
Changes in weather often trigger a myriad of negative impacts on the environment, which eventually affect human health. During the early months of 2016, Malaysia experienced El Niño, with an extremely dry season of almost zero rainfall. At the same time, an increase of more than twofold in fecal secretary immunoglobulin-A (SIgA) levels of healthy preschool children aged 2–6 years was observed, accompanied by an increase in phylum Bacteroidetes, predominantly attributed to genus Bacteroides and Odoribacter, which also positively correlated with fecal SIgA levels. Here, we present evidence to illustrate the detrimental effects of weather change on a microscopic “environment,” the human gut ecosystem. We also discuss the protective effects of probiotic against dysbiosis as induced by weather change. The increase in Bacteroidetes was at an expense of decreased genus Faecalibacterium and Veillonella (phylum Firmicutes), whereas children consuming probiotic had a decrease in genus Collinsella, Atopobium, and Eggerthella (phylum Actinobacteria) instead.
Weather Change: a Need to Evaluate the Gut Microenvironment
W
The establishment of the gut microbiota in infants is affected by factors such as delivery mode, methods of milk feeding, and the introduction of solid foods. 1 These factors also affect gut microbiota compositions in different populations and age groups around the world. The diversity of gut microbiota increases with age until it becomes a stable adult microbiota. It is generally reported that within a 3-year period after birth, the phylogenetic composition of the bacterial communities of a healthy child evolves toward an adult-like gut microbiota. 2 Bifidobacteria, for instance, are the main indication for age-specific changes in microbial composition. 3 In the gut of breastfed infants, bifidobacteria contribute up to 95%, while only 4.3 ± 4.4% bifidobacteria are present in the adult colon. 4,5
As reviewed by D'Argenio and Salvatore, the core gut microbiome (phyla Actinobacteria, Proteobacteria, Firmicutes, and Bacteroidetes) in healthy conditions vary across age. 6 Despite individual and geographical variations, in infants aged up to 2–3 years, the composition of Actinobacteria is the most abundant, followed by Proteobacteria, Firmicutes, and Bacteroidetes; while in healthy adults, the sequence of the phylum level is: Firmicutes > Bacteroidetes > Actinobacteria > Proteobacteria. Although the gut microbiome appears stable throughout adulthood, it becomes more unstable with lower diversity in the elderly. 6,7
Although healthy children in Japan have a higher abundance of Actinobacteria in the early ages (1–2 years), a shift was observed in the dominance of Firmicutes at the ages of 3–4 years, which was maintained throughout adulthood. 8 Meanwhile, Firmicutes dominated the gut microbiota composition of Malaysian children at the ages of 2–6 years, with a higher abundance at age 2 and reduced at age 3–6 years (Fig. 1). However, their microbiota composition remains stable at age 3–6 years.

Phyla of fecal microbiota from healthy preschool Malaysian children aged 2–6 years (April/May 2015). Total N = 100; N = 3 for age 2, N = 18 for age 3, N = 30 for age 4, N = 24 for age 5, N = 25 for age 6.
During a human clinical study involving a probiotic (Bifidobacterium longum BB536) and preschool children aged 2–6 years for a 10-month period, 8 we observed that at the end of study (10th month; February/March 2016), an increase of more than twofold in fecal secretary immunoglobulin-A (SIgA) levels of all children occurred, regardless of whether they were consuming placebo or probiotic (Fig. 2A). SIgA levels of different age groups were similar within the same period, and only differed upon comparison between different periods of time (April/May 2015 vs. February/March 2016; Fig. 2B). At the end of the study, Malaysia experienced El Niño, with an extremely dry season of almost zero rainfall. IgA is crucial for maintenance of intestinal homeostasis and gut immunology, 9 whereas certain gut microbiota such as Bacteroides have been reported to induce IgA production in the gut. 10 –12
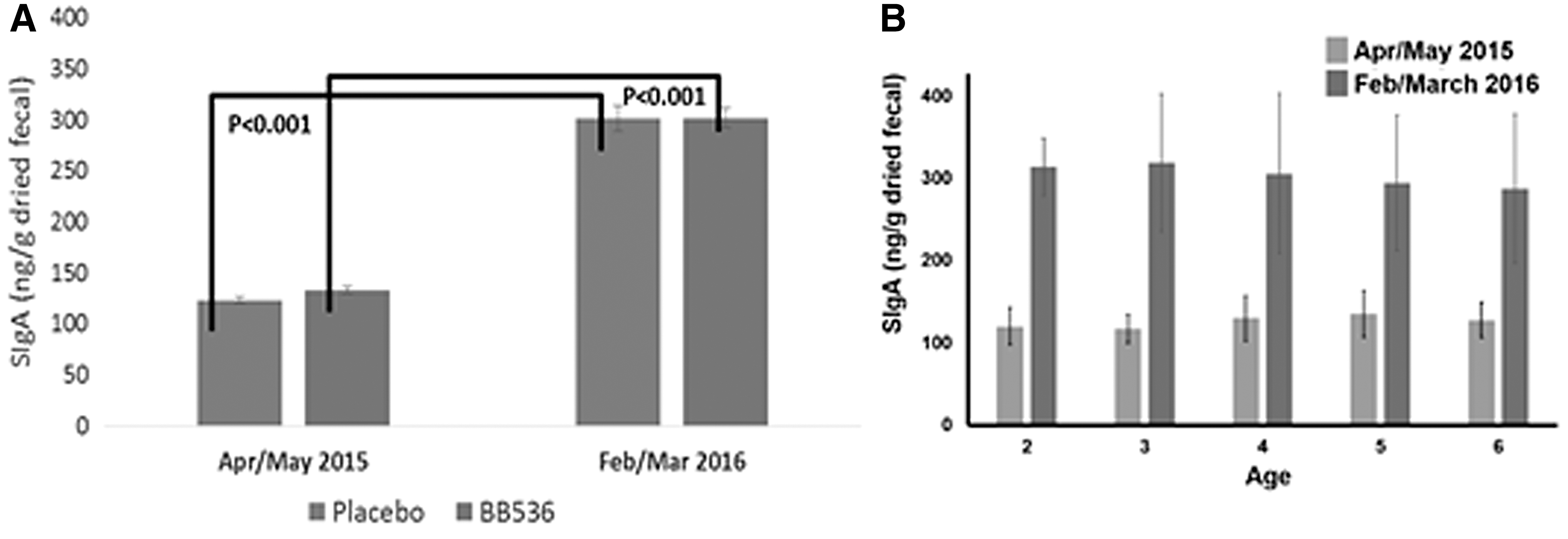
In this review, we present the evidence to illustrate the effects of weather change on an unexpected perspective of humanity,changes of gut microenvironment in healthy young children (Fig. 3).

Devastating effects caused by weather changes toward environment, ecosystem, and human.
There is a need to evaluate the impact of weather change on human gut ecosystem, as changes in the gut microbiota are often “silence,” without extremely clear displays of a destructive health condition. Contrary to most beliefs that gut microbiota are primarily affected by diseases and diets, it is also observed that a close relationship between weather and gut microbiota lies in the aspects of nutritional scarcity as affected by extreme external changes (temperature and rainfall). Malnutrition, particularly in children below 5 years old, resulted in dysfunction of intestinal microbiota to exert beneficial effects. This dysfunction is known as dysbiosis. 13
Probiotics, being one of the frontiers in gut health and modulation, are another aspect closely related to gut microbiota. This review also presents the evidence of protective effects of probiotics against detrimental gut microbiota changes as induced by weather changes. We also highlight the importance of beneficial bacteria in the gut as buffers amid unexpected external changes. Altogether, this serves as new knowledge combining a global agenda of weather change with a microscopic matter of gut microbiota and probiotic.
El NiÑo in Malaysia
El Niño is a climate pattern in which the ocean atmosphere is disrupted with a warm current of water that replaces the usual cold current of water in the eastern tropical Pacific Ocean. 14,15 Under normal conditions, in the tropical Pacific, the trade winds blow from east to west across the Pacific Ocean along the equator. 16,17 The strong trade winds push the warm current of water from the eastern Pacific toward the western Pacific (eastern coast of Asia, Indonesia, and Australia), while the cold current of water in the east wells up from below, leading to a temperature imbalance across the ocean. This promotes the trade winds to keep blowing, 16 resulting in the accumulation of warm water in the western regions (∼0.5 m higher), increasing heat (∼7°C warmer) and moisture, causing warmer and abundant rainfall in the western regions, and cooler and drier air in the eastern regions. This is also known as an upwelling process, 16 which occurs irregularly at 2–7-year intervals. 15
However, during El Niño, the abnormal warming of the ocean leads to the reversal or weakening of the prevailing trade winds and air pressure in the western Pacific. The warm water washes back to the eastern Pacific, thickening the depth of the warmer upper layer as compared with the cold water below, thus deepening the thermocline. 16,17 The disruption of the ocean atmosphere impacts rainfall patterns throughout the world. The increase in ocean temperature in the central and eastern Pacific brings additional heat and moisture, thereby increasing precipitation and larger areas of thunderstorm clouds. 15,17 Contrarily, the atmospheric pressure increases and precipitation reduces in the western Pacific, leading to drier weather. 15
The great impact of the El Niño spread throughout the globe, affecting the Pacific region, Asia, Australia, South and Central America, North America, and Africa. 18 The 2015/2016 El Niño resulted in drought, which reduced crop production and food shortages in Australia, Philippines, Indonesia, Thailand, Vietnam, Papua New Guinea, India, and Africa. 19 Malaysia experienced the most intense El Niño events in 1997–1998, recording the country's maximum temperature at 40.1°C, causing drought and haze conditions. 20,21
In the 2016 El Niño, an average maximum temperature of 32°C was recorded in Penang (northern Malaysia), whereas the highest temperature of 39.3°C was recorded in Pahang (central Malaysia). 21 Although the maximum temperature was not as high as that during 1997/1998, <1.0 mm precipitation was observed from January to April 2016 in Penang. An average of 0.5 mm precipitation in January, 1.0 mm in February, 0 mm in March, and 0.8 mm in April (Fig. 4) were observed. This indicates that the severity of the 2016 El Niño was attributed to severe drought rather than temperature. A Similar situation was observed in eastern and southern Australia in the first 4 months of 2016, recording rainfall in the lowest 10% in large parts of the northern coastline. 22 Comparing with December 2015, the average precipitation in 2016 was reduced drastically: as much as 85% in January, 71% in February, 100% in March, and 76% in April. A similar condition was also reported in other areas: Kedah, Perlis, north Perak, Kelantan, and east Pahang, with reduced rainfall up to 60%. 21
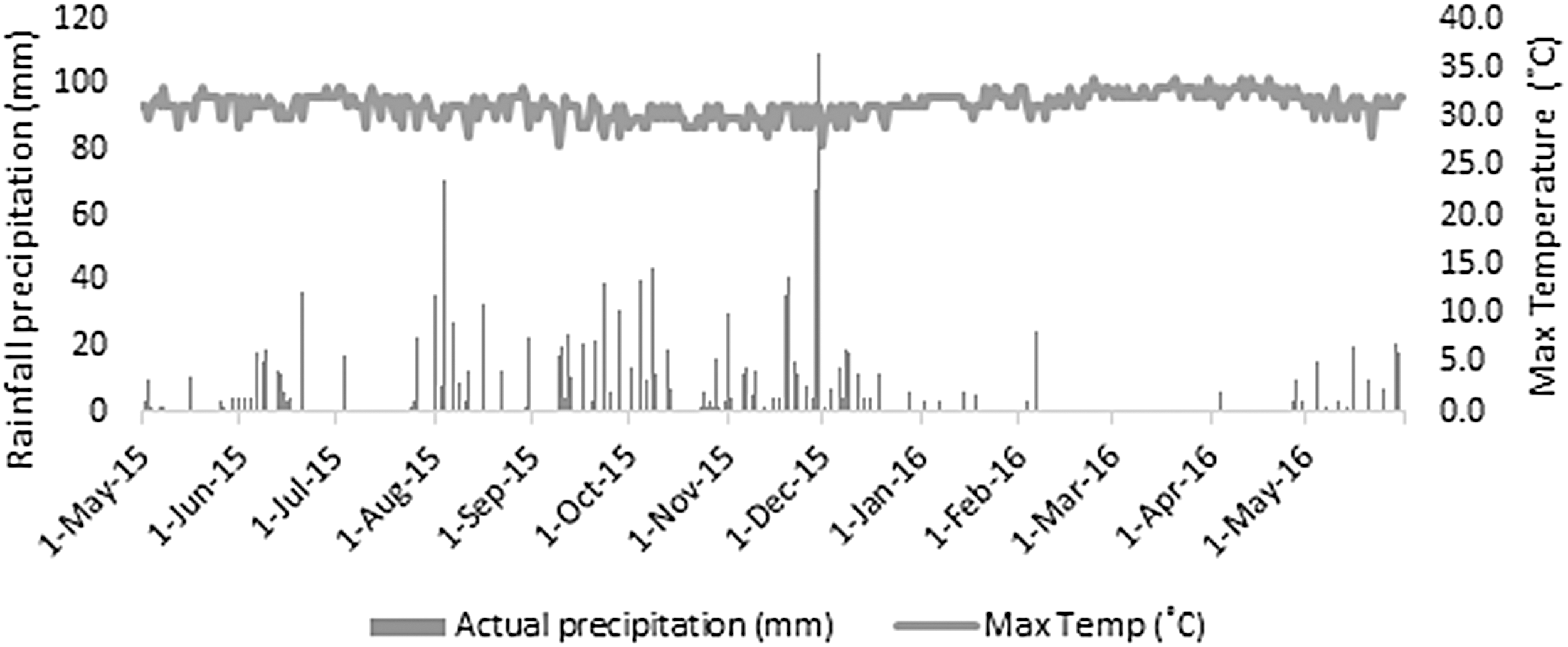
Average rainfall precipitation and average maximum temperature recorded in Penang (northern Malaysia). Results were obtained and tabulated from:
The average temperature in Malaysia increased up to 2°C from January to April 2016, compared with that during December 2015, the most drastic change experienced during the past 120 years, as compared with the usual annual increase of 0.2–0.6°C. 23 An increase of 1–2°C would cause devastating changes, although barely noticeable. Biologically, an additional increase of 0.5°C would result in greater droughts and prolonged heatwaves, reducing crop yield and endangering all the coral reefs, 24 whereas an increase of 1–3°C could affect protein folding, energy metabolism, circadian rhythms, and immune responses of many living species. 25
The 2015/2016 El Niño was among the strongest event that had affected >60 million people worldwide 26 and has increased global average temperatures. 27 Owing to drought conditions, temperature warming, flooding, and heavy rains in eastern Pacific regions during the El Niño, an increase in disease outbreaks, heat stroke, and respiratory diseases was reported. 26,28 Infectious diseases, including dengue, malaria, and cholera outbreaks, were often elevated amid increasing temperature and floods. 29 In South and Central America, El Niño flooding in December 2015 resulted in outbreaks of mosquito-borne diseases, such as Zika virus, and Rift Valley fever in East Africa and cholera outbreaks in Kenya. 18 In Malaysia, dengue incidences were high during El Niño 2016, with as much as 328.3 cases in 100,000 populations, and a total of 101,357 cases with 0.23% fatality. 30 Respiratory diseases were also increased, contributing to 13.78% of hospitalization cases, whereas 14.57% of hospitalization cases were attributed to infectious and parasitic diseases, and 6.14% attributed to diseases of the digestive system. 31
The External Environment Affects the Gut Microenvironment
Gut microbiota eventually affect the gut microenvironment and thus have attracted increasing interests. A recent cross-sectional study involving 367 healthy subjects aged 0 (infants) to 104 years has illustrated the sequential changes of human gut microbiota as a factor of time and aging, 2 indicating that, similar to aging, changes in gut microbiota are an inevitable natural process. Many studies have emphasized and reported on intrinsic factors that change the gut microenvironment: genetics, 32 diet, 33 diseases, 34 lifestyles, 35 sexual preference, 36 and social emotional status. 37
One of the major extrinsic factors is the surrounding environment, typically weather changes. The hypothesis remains that the outer environment affects the gut microenvironment. Studies to date have shown that although seasonal changes have caused temporal variations in microbial community structure within natural populations, such a shift was not entirely and primarily attributed to changes in weather, but rather, an intrinsic factor of diet as well. Populations ranging from animals to humans often experience a change in diets across different seasons.
A 2-year observation of gut microbiota from wild wood mice (Apodemus sylvaticus) inhabiting two different woodlands showed that seasonal changes dominated the shifts in gut microenvironment. It was postulated that a strong seasonal shift caused transitions in diets, where mice moved from an insect- to a seed-based diet upon seasonal changes. The abundance of Lactobacillus was decreased, accompanied by increased levels of Alistipes (Bacteroidetes phylum) and Helicobacter. 38
Similar conclusions were also observed in larger explorations on wild primates, wherein seasonal shifts have changed their gut microbiota. Gut microbiota of the Tibetan macaque (Macaca thibetana) varied during winter and spring seasons. The genus Succinivibrio, which promotes the digestion of cellulose and hemicellulose, was significantly increased during winter, whereas the genus Prevotella, which is associated with digestion of carbohydrates and simple sugars, was significantly increased in spring. In winter, the primary diets of these primates revolve around cellulosic and hemicellulosic carbohydrates, yielding the need to extract glycan for digestion and, thus, the increased abundance in microbiota that are capable of digesting glycan. Meanwhile in spring, an increased abundance of carbohydrate-metabolizing and energy-metabolizing microbiota is needed to facilitate recovery from acute energy loss during winter. 39
In an attempt to evaluate the effects of seasonal changes on human gut microenvironment, Davenport et al. examined variation in gut microbiota between winter and summer for 12 months in 60 members of the Hutterites in North America. 40 Hutterites practice a communal lifestyle and thus have similar and stable diets across individuals throughout the year. Fresh produce is primarily served during the summer and autumn months. Amid overall gut microbiome stability within individuals over time, there were consistent and significant population-wide shifts in microbiome composition across winter and summer seasons.
Bacteroidetes is more prevalent during summer than during winter, whereas Actinobacteria is more abundant during winter than during summer. Such a shift in microbiome composition is attributed to changes in diets during different seasons; high levels of produce containing complex carbohydrates consumed during the summer months encouraged the growth of Bacteroidetes (complex carbohydrate digesters), and Actinobacteria do not thrive well in fibrous foods. Seasonal shifts have defied dietary needs across populations ranging from animals to humans, causing the gut microenvironment to evolve as a mean to buffer against energy and nutrient challenges, enabling the adaptation and survival of species.
There are also a few animal studies reported on changes in gut microbiota affected primarily by weather changes. It is challenging to carry out direct correlation studies of gut microbiota and weather changes in human model as it is impractical to control the subject's environment for a long period of time. In human models, there are a number of variables such as diet, geographical, genetic background, access to medical care, sanitary practices, and pathogen exposure across populations, in addition to weather changes. 40
A laboratory set experiment involving cold exposure of 10 days on C57BI/6J mice led to a marked shift of microbiota composition, producing “cold microbiota.” 41 The cold microbiota consisted of increased Firmicutes, reduced Bacteroidetes, and absence of Verrucomicrobia as compared with mice not subjected to cold treatment. Transplantation of the cold microbiota to germ-free mice increased the host's insulin sensitivity and enabled cold tolerance, by promoting white fat browning, leading to increased energy expenditure and fat loss. Mammalian white adipose tissue stores energy in the form of triglycerides, whereas brown adipose tissue catabolizes lipids to produce heat. A loss of brown fat function has been linked to obesity and metabolic diseases. 42
In response to cold or exercise, brown fat emerges in white fat through white fat browning. In addition, transplantation of cold microbiota also led to an increased caloric uptake through increased intestinal absorptive surface and intestinal gene expressions for tissue remodeling, accompanied by increased lengths of intestines, villi, and microvilli. These strongly illustrate that microbiota also buffer against temperature-induced energy demand, protecting hosts against homeostatic fluctuations.
Another latest study on weather change affecting gut microbiota was done on an ectotherm, the common lizard (Zootoca vivipara). 43 A large seminatural enclosure in which climate manipulation was used and a mere 2–3°C warmer temperatures caused a 34% loss of the lizards' microbiota diversity. Warming caused a decrease in Bacteroidetes (Bacteroidales) and Firmicutes (Lachnospiraceae), while increased in Proteobacteria. Prolonged warm temperatures (3 months) had long-lasting deleterious effects (8 months) on gut bacterial diversity, resulting in lower bacterial richness. A greater gut bacterial diversity is crucial, whereas a weather-driven diversity reduction is detrimental for hosts, predominantly if it involves a loss of vital functions. Bacteroidales and Lachnospiraceae are two important reptile microbiota, and thus their diminution may affect nutrient assimilation or inhibition of pathogens, leading to endangerment against survivability. These may be one of the reasons to explain species extinction amid weather changes. Such investigations disclose the buffering roles of the gut microenvironment against deviations raised from changes in external surroundings.
El NiÑo, The Gut Microenvironment of Malaysian Children, and Possible Contribution of Probiotics
Rise of Bacteroidetes: Bacteroides and Odoribacter
In Malaysian preschool children, the phylum Bacteroidetes increased in fecal samples of all children comparing April/May 2015 (before El Niño) and February/March 2016 (during El Niño), regardless of placebo or probiotic consumption, accompanied by a significant positive and low correlation with fecal SIgA levels (Fig. 5A–C; full abundance data are shown in Supplementary Table S1. Further evaluations revealed that this is attributed to the increase of two genera within this phylum, Bacteroides and Odoribacter, which both also showed significant positive and low correlations with fecal SIgA levels (Fig. 6A–C).
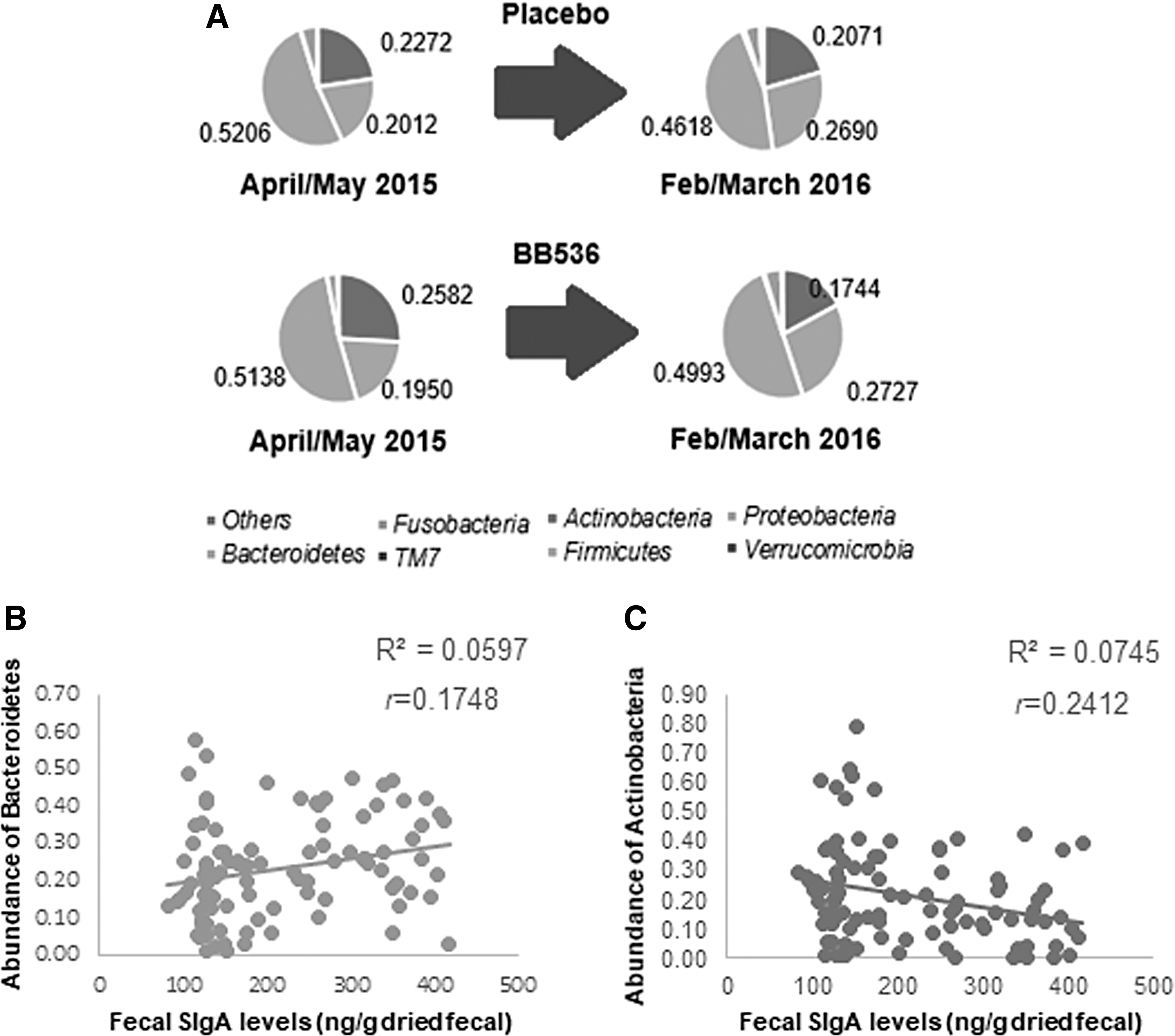
Characterization of microbiota phyla in fecal samples from Malaysian preschool children aged 2–6 years before El Niño (April/May 2015) and during El Niño (February/March 2016) who were consuming placebo and probiotic B. longum BB536

Genera of the phylum Bacteroidetes are characterized in fecal samples from Malaysian preschool children aged 2–6 years before El Niño (April/May 2015) and during El Niño (February/March 2016) who were consuming placebo and probiotic B. longum BB536
Bacteroides primarily breaks down host polysaccharides and dietary plant-based undigested carbohydrates. Certain Bacteroides are also producers of polysaccharides that activate T cell-dependent immune responses, thus limiting pathogenic gut colonization. A zwitterionic polysaccharide produced by Bacteroides fragilis reportedly activated T helper cells (CD4+) that bind onto antigen-presenting cells, thus presenting the antigens to T cell receptors. 44 Splenocytes from germ-free mice had lower proportions of CD4+ cells than splenocytes from conventional mice, whereas colonization with B. fragilis in germ-free mice corrected this proportion, indicating the role of B. fragilis in the activation of T cell-dependent immune responses. 45 Meanwhile, Bacteroides thetaiotomicron reportedly induced Paneth cells (immune system cells located in small intestinal crypts) to produce antibacterial peptides that bind to the peptidoglycan of gram-positive pathogens (such as Listeria monocytogenes), thus preventing colonization in the small intestines. 46
There is a need for gut microflora to competitively exclude the foreign invasions caused by illnesses triggered by El Niño through host immune systems, and this may include the production of SIgA. The levels of SIgA vary predominantly between different populations but are less affected by children's age, 47 thus the long-term study of almost 1 year would not have attributed to such a drastic change in SIgA concentrations.
Intestinal mucosae possess independent humoral immune systems, with IgA being produced by mucosal plasma cells. IgA is secreted into the gut to function as a binder for antigens, foods, and bacteria to prevent their adhesion to or penetration into the lamina propria in a process known as immune exclusion. 48 The bacterial, viral, or parasitic infections of childhood are associated with an increased rate of IgA production in response to antigenic stimulation; thus, mucosal SIgA levels are good markers of immunological events in the digestive tract. Considering that our gut is a reservoir of microorganisms, any changes in gut IgA levels must be directly associated with gut microbiota. In germ-free mice, IgA production was barely detectable in the small intestines and was completely undetected in the large intestines. 49 Gut microbiota such as Bacteroides acidifaciens are predominant in promoting IgA production in the large intestines.
Although Odoribacter is not known for modulation of host IgA levels, a reduced abundance was observed in patients with Crohn's disease and ulcerative colitis, 50 suggesting their roles in intestinal immunological functions. Infections of Clostridium difficile lead to gut inflammation, as witnessed by increased fecal excretion of human DNA in hospitalized patients. A high proportion of pathogenic genera such as Enterococcus and Escherichia, but depleted of Odoribacter, was also detected in host fecal DNA. 51 As Odoribacter splanchnus was hypothesized to actively produce acetate, propionate, and butyrate, 52 a reduced abundance of Odoribacter and availability of short-chain fatty acids (SCFAs) eventually fail to buffer against host inflammation. SCFAs promote colonocyte differentiation and proliferation, which boosts the integrity of the intestinal epithelium against inflammation.
Fall in Firmicutes and Actinobacteria: Influences of probiotic
An increase in abundance of one phylum would naturally be at the expense of another phylum/phyla. In a clinical study, it was found that consumption of probiotic B. longum BB536 affected the abundance of Bacteroidetes and Actinobacteria. It was noticed that the increased abundance of Bacteroidetes was due to a decrease in various phyla, such as Actinobacteria. Children consuming placebo showed reduced phylum Firmicutes between April/May 2015 (before El Niño) and February/March 2016 (during El Niño), whereas children consuming BB536 showed reduced phylum Actinobacteria (Fig. 5A, and full abundance data in Supplementary Table S1). Actinobacteria also showed a significant negative and low correlation with fecal SIgA levels (Fig. 5C).
A proportional decrease in Firmicutes has been reported to be highly beneficial against various disorders and diseases, including obesity, 53 insulin resistance, 54 and asthma. 55 Meanwhile, a decrease in Actinobacteria, which is predominated by the genus Bifidobacterium, has been reported to affect host health, typically in children. The proportion of Actinobacteria was lower in malnourished, 56 type I diabetes, 57 asthma, and rhinitis children 55 and was associated with the pathogenesis of food allergy in children. 58 Thus, at first glance, the impact of El Niño on gut diversity in children seemed more beneficial than detrimental, whereas the administration of probiotic seemed not to have benefited the children's gut ecosystem.
Subsequent evaluations revealed that a marginal decrease of Firmicutes in placebo group children was attributed to a marginal decrease of two genera within this phylum: Faecalibacterium and Veillonella (Fig. 7A).

Genera of the phylum Firmicutes are characterized in fecal samples from Malaysian preschool children aged 2–6 years before El Niño (April/May 2015) and during El Niño (February/March 2016), who were consuming placebo and probiotic B. longum BB536
Faecalibacterium also showed a significant negative and low correlation with fecal SIgA levels (Fig. 7B). Faecalibacterium, a known butyrate producer typically under increased consumption of resistant starch, has the ability to metabolize complex carbohydrates, including legumes and whole grains. Butyrate serves as a crucial energy source for colonocytes, reduces the risks of colon cancer, and is used in treatments of colitis and irritable bowel disease with diminishing intestinal inflammatory properties. 59 Faecalibacterium is important in gut homeostasis and has been observed to decrease in patients with gut dysbiosis and Crohn's disease 60 and irritable bowel disease with ileal involvement. 61
The genus Veillonella (a minor component of the phylum Firmicutes) has been reported to exert immunomodulatory properties. A common species of the human small intestinal microbial communities, Veillonella parvula, portrays a strong capacity to induce IL-6 individually. Meanwhile, in combination with some streptococci, Veillonella reportedly reduced IL-12p70 production and augmented the responses of the anti-inflammatory cytokine IL-10. 62 IL-12p70, a cytotoxic lymphocyte maturation factor and natural killer cell stimulatory factor, is primarily produced by dendritic cells and macrophages as a powerful inducer of interferon gamma, a proinflammatory cytokine.
Thus, amid El Niño, Malaysian children experienced a decrease in the beneficial genera of Faecalibacterium and Veillonella, which was not observed in children who were consuming the probiotic BB536 (Fig. 7A). Several hypotheses are raised: (1) probiotic exerted a buffer against declining Faecalibacterium and Veillonella populations through symbiosis efforts with these genera and/or other gut microbial communities; (2) IgA might have exerted some growth inhibitory properties while probiotic buffered against such an effect; (3) health changes as induced by El Niño might have included foreign pathogenic invasions that competitively reduced the growth of Faecalibacterium and Veillonella, whereas the probiotic inhibited the growth of these pathogens. Evidence to support these hypotheses remains scarce, as the effects of weather changes on the human gut ecosystem remain unknown.
Although Faecalibacterium and Veillonella are beneficial for gut health, they are not reported to be IgA producers. Also, although IgA is stable within the intestinal tract to crucially mediate host–microbiota homeostasis and regulate transient antigenic stimulation, there is no necessity for IgA to promote and/or protect the growth of gut-friendly microorganisms. The implications of IgA on gut microbial community are not well understood. Here, we are presenting the possibility of reduced gut protective genera such as Faecalibacterium and Veillonella that are detrimentally reduced amid increasing IgA levels, whereas the presence of probiotic buffered against such an effect. A recent finding has reported that the administration of a probiotic, Enterococcus faecium EP1, in mice led to increased fecal SIgA, but, at the same time, also increased fecal Faecalibacterium prausnitzii counts. 63
Meanwhile, consumption of probiotic BB536 resulted in a decrease in the abundance of three genera: Collinsella, Eggerthella, and Atopobium, which resulted in a decrease in the abundance of Actinobacteria in children's feces, whereas children consuming the placebo did not show any changes in all genera (Fig. 8A). Eggerthella also showed a significant negative and low correlation with fecal SIgA levels (Fig. 8B).
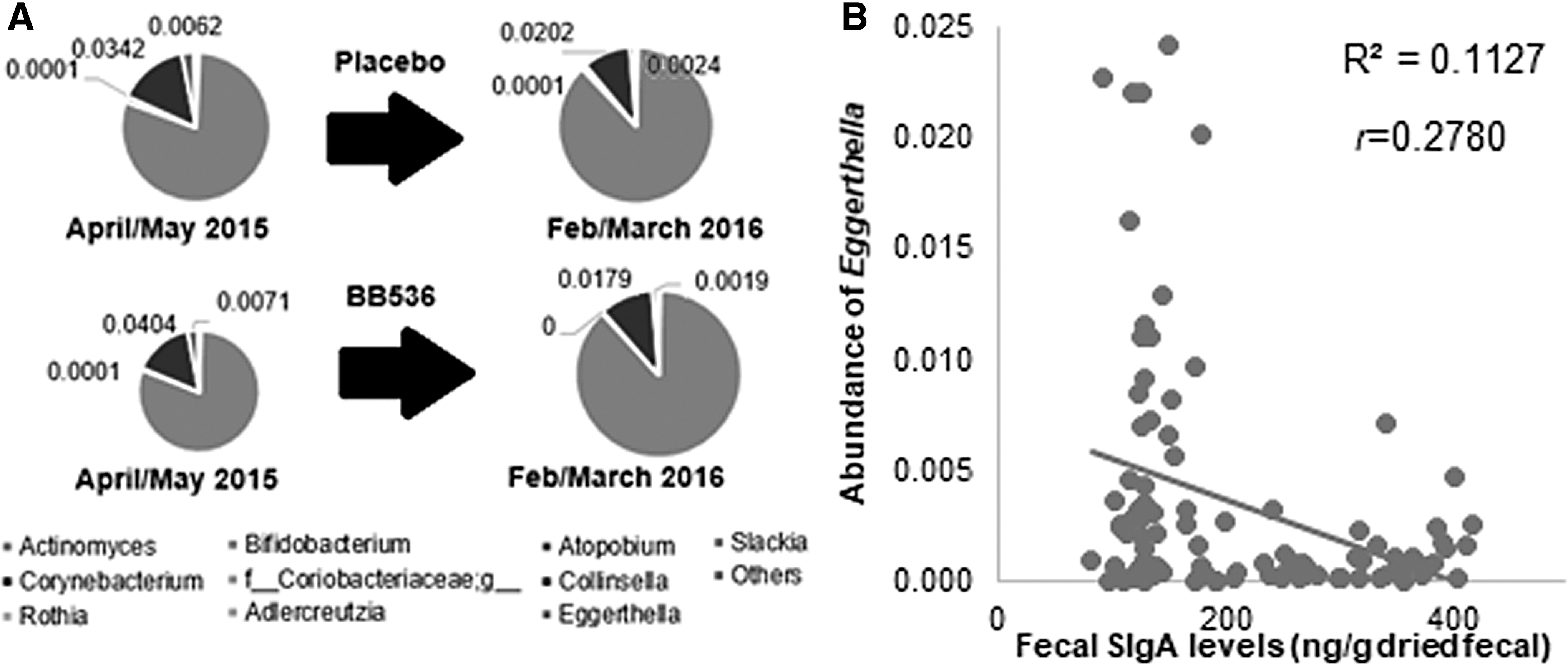
Genera of the phylum Actinobacteria are characterized in fecal samples from Malaysian preschool children aged 2–6 years before El Niño (April/May 2015) and during El Niño (February/March 2016), who were consuming probiotic B. longum BB536 and placebo
Eggerthella commonly resides in the human colon and has been dubbed as an emerging pathogen. Certain species such as Eggerthella lenta, Eggerthella hongkongensis, and Eggerthella sinensis have been implicated in cases of ulcerative colitis, liver and anal abscesses, abdominal sepsis, and systemic bacteremia. 64,65 Although Collinsella has received less publicity as a gut pathogen, emerging pieces of evidences are showing that the abundance of this genus was higher in patients with rheumatoid arthritis than in healthy controls, with prominent roles in altering gut permeability possibly attributed to its metabolites such as beta-alanine, alpha-aminoadipic acid, and asparagine. Meanwhile, experiments using humanized mice have found that Collinsella aerofaciens enhanced rheumatoid arthritis severity. 66
During the process of immune exclusion, SIgA in the gut binds to foreign matters, including antigens and bacteria, before their detection and elimination. 48 IgA coating has been reported to be a useful strategy to distinguish colitogenic members among intestinal microbiota. In mice with intestinal dysbiosis, and patients with Crohn's disease and ulcerative colitis, the proportion of intestinal bacteria that were coated with IgA was higher than in healthy controls. 67
Palm et al. reported that although many species were present in patients with gut disorders, only several species were highly coated with IgA, including C. aerofaciens in patients with inflammatory bowel diseases and E. lenta in ulcerative colitis patients. 68 Sugahara et al. reported the possible contribution of decreased IgA response in the increased abundance of bacterial taxa with potential pathogenicity in the intestinal environment of the elderly. 69 Considering that the reduction of Collinsella and Eggerthella in Malaysian children amid El Niño was only observed in the probiotic group, it can be hypothesized that the probiotic enhanced IgA binding onto these bacteria, leading to their increased elimination and reduced counts in feces.
All these observations highlighted the need for deeper evaluations beyond merely higher levels, such as phyla and/or families, which may provide misleading conclusions. The evaluations on genus and species would yield a better perspective on the complex nature of the human gut microbiota. The changes in weather and temperature often trigger biological and physiological stress responses in animals and humans. 70 Along the bilateral relationships of the gut–brain axis, evidence has manifested the roles of stressors in changing compositions of the gut microbiota 71 and the roles of gut microbiota in influencing anxiety and depression. 72 We believe that the changes in weather and temperature during El Niño have triggered physiological stresses in these Malaysian children, leading to changes in gut microbiota and SIgA levels.
Concluding Remarks
This review has provided evidence on the possibility that weather changes could affect the human gut microenvironment and indiscriminatively also affected young children. Invasions by pathogens and antigens that increased during the dry period of El Niño may have triggered the need for the gut microbiota to buffer against changes as a gut modulatory response. The increase in fecal SIgA, Bacteroidetes, and Bacteroides supports such a possibility. Weather changes also detrimentally reduced gut beneficial bacteria such as Faecalibacterium and Veillonella, whereas the administration of the probiotic protected against such an onslaught in addition to promoting a decrease in less beneficial bacteria such as Collinsella and Eggerthella.
As changes in gut microbiota are often “silence,” this review illustrates an important impact of weather changes that is surreal, may exert long-term implications, and should not be ignored. Meanwhile, probiotic remains beneficial to buffer hosts against unexpected external changes and could be better designed as a health strategy in alternative medicine.
Footnotes
Acknowledgments
This work was supported by Morinaga Milk Industry Co. Ltd. (Grant No. 304/PTEKIND/650689) and USM Fellowship provided by Universiti Sains Malaysia, Penang, Malaysia.
Authors' Contributions
M.T.L., J.-Z.X., and A.S.Y.L. performed the clinical trial, analyzed the data, and drafted and revised the article. T.O. and E.M. conducted microbiota analysis.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.