
Abstract
Aging and lifestyle factors, including high-sugar and high-fat diets, promote a systemic metabolic imbalance that promotes neurodegeneration. Hericium erinaceus has long been used in traditional Chinese medicine. Recently, its functional activities, such as antimetabolic dysfunction, antineuroinflammatory activities, and stimulation of nerve growth factor (NGF) synthesis, have been revealed. This study demonstrated that Hericium erinaceus mycelium (HEM) and an isolated diterpenoid derivative, erinacine A (EA), may reverse spatial learning disabilities in aging mice (15 months old) fed with a high-fat and high-sucrose diet (HFSD). Aging mice were randomly assigned to one of four treatment groups: (1) a chow diet (control), (2) an HFSD, and an HFSD supplemented with either (3) HEM or (4) EA for 18 weeks. The Morris water maze (MWM) and Y-maze were used for behavioral assessments. Both HEM- and EA-treated mice had shorter mean daily escape latencies than HFSD-treated mice in the MWM. In addition, HEM-treated mice had a slightly increased exploratory time and frequency in the novel arm in the Y-maze. Quantitative PCR revealed that both HEM- and EA-treated mice exhibited reduced messenger RNA (mRNA) expression of tumor necrosis factor-α, interleukin-1β, and HEM-treated mice exhibited increased mRNA expression of NGF and NeuN in the hippocampus. Moreover, HEM and EA also decreased body weight, abdominal fat, plasma glucose, serum and liver total cholesterol, and liver triacylglycerol. Thus, HEM may be a potential health-promoting supplement for minimizing the progression of aging and obesity-induced neurodegeneration by reducing metabolic abnormalities and neuroinflammatory cytokines and increasing neurogenesis factors.
Introduction
Epidemiological surveys have revealed that the incidence of cognitive disorders, such as dementia, increases with aging. The prevalence of dementia is 2.5–5.0% in people 65 years of age or older; however, the prevalence increases to 20% in people 85 years of age. 1 As the elderly population increases, and with the consumption of diets rich in saturated fats and refined sugar, obesity in the elderly population is expected to increase rapidly. Diets with high levels of both fat and sucrose have been well documented to be risk factors for many medical conditions, including obesity, hypercholesterolemia, nonalcoholic fatty liver disease, and metabolic syndrome. 2 Furthermore, extensive evidence has linked high-fat and high-sucrose diets (HFSDs) with an increased risk of dementia in humans and rodents. 3 –6 Chronic inflammation resulting from obesity was reported to be related to the development of dementia and cognitive deficiencies. 7 Mice fed with a high-cholesterol diet tended to exhibit poor learning performance. 8 These data also imply a relationship between plasma cholesterol levels and Alzheimer's disease (AD). Moreover, nonalcoholic fatty liver disease resulting from a high-fructose diet has been documented to impair hippocampal-dependent memory function in rats. 9 Therefore, maintaining a healthy diet containing functional foods is very relevant for protecting the elderly population from developing dementia and other chronic medical conditions, and represents an effective approach to alleviating the burden and debilitating effects of dementia in the aging population.
Hericium erinaceus is a mushroom that grows on both living and dead broadleaf. This mushroom is popular for its use in different culinary preparations in China, Japan, and Korea. For example, H. erinaceus has been used in the treatment of gastritis in traditional Chinese medicine for >1000 years. 10 Thus, the chemical constituents of H. erinaceus have attracted interest because of their significant biological and pharmacological effects, including antioxidant effects, 11 hypolipidemic effects, 12 hemagglutinating activity, 13 antimicrobial activity, 14 antiaging effects, 15,16 immunomodulation, and anticancer activities. 17,18 Erinacines are important secondary metabolites isolated from the mycelium of H. erinaceus and are potent stimulators of nerve growth factor (NGF) biosynthesis. 19 –21 Erinacine A (EA) is abundant in the mycelium of H. erinaceus, and its profound NGF-inducing activity has been demonstrated both in vitro and in vivo. 16 NGF is a potential biological activator that prevents neuronal death, promotes neurite outgrowth, and maintains and organizes neuronal functions in the central nervous system. 22 The application of NGF in the treatment of AD has been proposed in many studies. 23,24 However, earlier studies on the effect of H. erinaceus on NGF biosynthesis were mostly restricted to the pure compounds extracted from its mycelia, and few studies have explored the potential of the cultured mycelial complex in neurodegenerative diseases. Moreover, a previous study demonstrated the hypolipidemic effect of an exo-biopolymer produced from a submerged mycelial culture of H. erinaceus. 12
Based on these observations, we investigated whether supplementation with the mycelium of H. erinaceus can prevent the HFSD-induced cognitive impairment and confer neuroprotection by rescuing metabolic abnormalities. We investigated and evaluated the efficacy of Hericium erinaceus mycelium (HEM) and EA against HFSD-induced cognitive impairment in aging mice. Furthermore, we provide evidence for the application of HEM and EA in the treatment of dementia caused by an unhealthy lifestyle.
Materials and Methods
H. erinaceus extracts and analysis of EA
Fresh mycelium of H. erinaceus was extracted with methanol. The extract was concentrated and partitioned between ethyl acetate and water. The ethyl acetate fraction was subjected to silica gel gravity column chromatography using n-Hexane-ethyl acetate of increasing polarity as the eluent.
The high performance liquid chromatography (HPLC) analytical column of EA used was a Mightysil RP-18 GP (4.6 × 250 mm; particle size 5 μm; Kanto Chemical CO., INC.). Separation was performed at 40°C using two different gradients for the mobile phase, which consisted of two solvents, water (A) and acetonitrile (B). The gradient elution had the following profile: 0–30 min, 80–50% (A), and 30–40 min, 50% (A). The retention time of EA was approximately 42 min at a flow rate of 1.0 mL/min with a scanning ultraviolet wavelength at 345 nm. The 5 mg/g concentration of EA in the H. erinaceus extracted with methanol (HPLC grade) was confirmed and quantified by HPLC as shown in Figure 1.
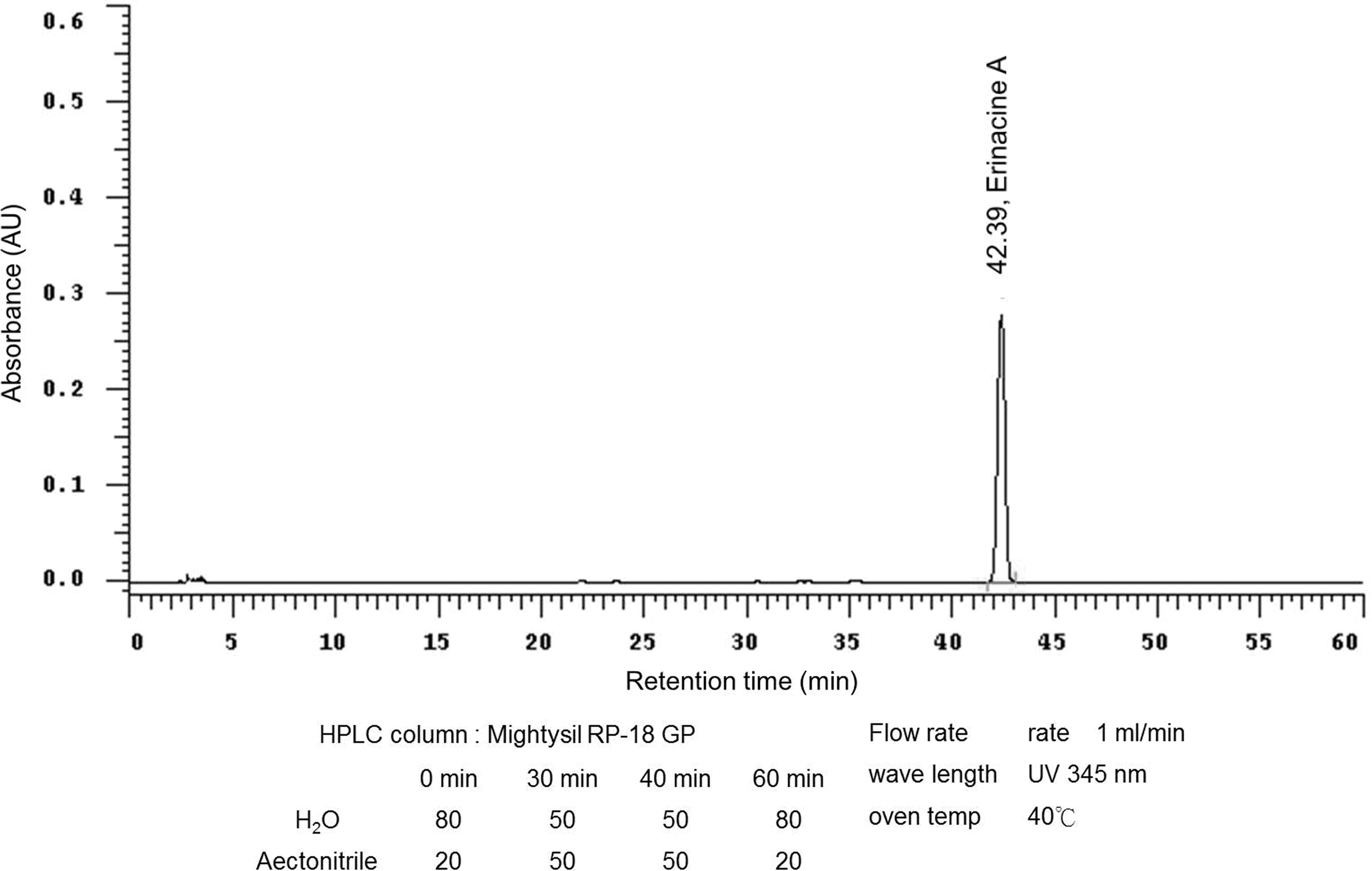
HPLC analysis of EA. The HPLC chromatograms of an EA standard sample and the methanol extract of HEM from a 5-ton bioreactor (UV detection at 345 nm). The retention time of EA is 42.4 min. EA, erinacine A; HEM, Hericium erinaceus mycelium; HPLC, high performance liquid chromatography; UV, ultraviolet.
Animals and treatment
C57BL/6 mice (15 months old, male) were obtained from National Laboratory Animal Center (Taipei, Taiwan). The animals were housed under a 12-h light/12-h dark cycle at a controlled temperature (24 ± 2°C) and humidity (60 ± 10%), and provided with food and water ad libitum according to the Institutional Animal Care and Use Committee of Chung Yuan Christian University (Taoyuan, Taiwan). The mice were randomly divided into four groups (n = 8–10), and each group received one of the following treatments for 18 weeks: group 1, the control group was fed a chow diet containing 13.5% of kilocalories from fat and 3.7% of kilocalories from sucrose (LabDiet 5001); group 2, the HFSD group was fed an HFSD containing 45% of kilocalories from fat and 17% of kilocalories from sucrose (TestDiet 58V8); group 3, the EA group was fed an HFSD mixed with EA (43 mg/kg); and group 4, the HEM group was fed an HFSD mixed with HEM (1 g/kg). The body weights of the mice were measured every week. Blood samples were collected every month to measure serum glucose, triacylglycerol (TG), and cholesterol levels. After 16 weeks of treatment, the mice underwent the Morris water maze (MWM) and Y-maze tests for the next 2 weeks and were then deprived of food for 12 h. The mice were then sacrificed by anesthesia with an intraperitoneal injection of a solution of tribromoethanol (250 mg/kg body weight; Avertin). The hippocampi and livers were immediately collected, snap-frozen in liquid nitrogen (N2), and stored at −80°C until analysis. All animal experimental procedures were carried out in accordance with the protocol approved by the Institutional Animal Care and Use Committee of Chung Yuan Christian University, Chungli, Taiwan (IACUC No: 103027).
MWM test
The MWM test was performed after 16 weeks of treatment. The apparatus consisted of a circular pool (diameter: 120 cm, height: 45 cm) filled with water (depth: 30 cm, 25 ± 0.5°C), and an escape platform (diameter: 10 cm) was placed in the middle of one of the quadrants 1 cm below the water surface and equidistant from the sidewall and the middle of the pool. The water tank was inside a test room containing spatial signs around the maze and remained undisturbed during the test. Each mouse was subjected to a series of four trials per day. For each trial, the mouse was placed into the water maze at four different positions. The trial began by placing the animal in the water facing the wall of the pool as one of the starting points. If the mouse failed to escape within 60 s, then it was gently directed to the platform. The mouse was allowed to stay on the platform for 15 s. The time between entering the water and climbing onto the platform was recorded as the latency to escape, and the mean latency until finding the invisible platform was calculated for each mouse. The probe trial was performed on the 6th day by removing the platform from the pool. The mice were allowed to swim for 60 s, and the time spent in the target quadrant was recorded. After each trial, the mice were dried, and they were returned to their cages at the end of the session. The animals were trained for 5 days. The performances were recorded and analyzed using video-tracking software (Ethovision XT 7; Noldus Information Technology, Wageningen, The Netherlands).
Y-maze test
Two days after the MWM test, the mice underwent the Y-maze test. The Y-shaped apparatus consisted of three equally spaced and enclosed arms (8 × 30 × 15 cm) with a 120° angle between adjacent arms. Several visual cues were placed on the inner walls of the maze. The three arms were designated as follows: the start arm (always open), where the mouse started to explore the maze; the familiar arm (always open); and the novel arm, which was blocked during the first trial and opened during the second trial. 25 The Y-maze test was performed in a dimly lit room and consisted of two trials separated by a 2-h intertrial interval. During the first trial (training phase), which lasted for 10 min, the mouse was allowed to explore two arms (the start arm and familiar arm) as the novel arm was blocked. During the second trial (retention phase), which lasted for 5 min, the three arms were opened and the mouse placed in the start arm was allowed to freely explore all three arms. The number of entries into each arm was recorded as each arm's entries and the sum of the time spent in each arm was recorded as the time spent in each arm. The maze floor and walls were cleaned after each trial to remove olfactory cues. The performances were recorded and analyzed using the video-tracking software Ethovision XT 7 as used in the MWM.
Quantitative real-time reverse transcriptase–polymerase chain reaction
Total RNA from the hippocampus was isolated using Trizol Reagent (Invitrogen, Carlsbad, CA) and reverse transcribed with RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Wilmington, DE, USA) according to the manufacturer's protocol. Real-time PCR (RT-PCR) was performed with an ABI 7300 system (Applied Biosystems, Foster City, CA, USA) using FastStart Universal SYBR Green Master Mix (Roche). Relative mRNA expression levels were calculated using the 2−ΔΔCt method. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the housekeeping gene for calculations. The primers used in this study are as follows: GAPDH: forward, 5′-GGGCCATCCACAGTCTTC-3′, reverse, 5′-CTGCACCACCAACTG CTTAG-3′; tumor necrosis factor-α (TNFα): forward, 5′-ATCTTCTCAAAATTCGAGTGACAAG-3′, reverse, 5′-TGGGAGTAGACAAGGTACAACCC-3′; interleukin-1β (IL-1β): forward, 5′-GAGCACCTTCTTTTCCT TCATCT-3′, reverse, 5′-GATATTCTGTCCATTGAGGTGGA-3′; NeuN: forward, 5′-TGAGGTCAATAATGCCACAGC-3′, reverse, 5′-ATAG AATTCAGGCCCATAGACT-3′; and NGF: forward, 5′-TCCACCCACCCAGTCTTCCA-3′, reverse, 5′-GCCTTCCTG CTGAGC ACACA-3′.
Serum chemistry analysis
After coagulation, mouse sera were collected from blood samples by centrifugation at 10,000g for 20 min at 4°C, and these sera samples were analyzed using a Fuji Dri-Chem 4000i analyzer (Tokyo, Japan) at the Taiwan Mouse Clinic. Biochemical parameters, such as glucose, total cholesterol (TC), TG, uric acid (UA), creatinine, glutamate oxaloacetate transaminase (GOT), and glutamate pyruvate transaminase (GPT) concentrations, were determined. Liver TC and TG were measured using an enzymatic photometric assay following the manufacturer's manual (DiaSys Diagnostic Systems GmbH, Germany).
Statistical analysis
All the experimental data were expressed as the mean ± standard deviation. The data from the training trial in the MWM were analyzed by repeated measures analysis of variance followed by Fisher's least-significant-difference post hoc test. Other significant differences between two groups (e.g., control and HFSD, HFSD and EA, or HFSD and HEM) were analyzed by the Mann–Whitney U test. *P < .05 indicated a statistically significant difference compared to the control group, while # P < .05 indicated a statistically significant difference compared to the HFSD group.
Results
Spatial learning performance in aged mice fed with the HFSD supplemented with HEM or EA
The aging mice (15 months old) were randomly assigned to one of four treatment groups: (1) a chow diet as the control group, (2) an HFSD, and an HFSD supplemented with either (3) HEM or (4) EA as the experimental groups for 16 weeks. Then, the mice underwent assessments for spatial learning and memory using the MWM and Y-maze tests, respectively. In the spatial learning test, all four groups showed a progressive decline in the learning curve during the acquisition phase of training (days 1–5). As shown in Figure 2A, the HFSD group had a prolonged escape latency time, which differed significantly compared to that in the control group. Interestingly, both the EA- and HEM-treated groups had significantly shorter escape latency times than the HFSD group and behaved analogously to the control group (Fig. 2A). In the probe trial on the sixth day, no difference was observed between any group (data not shown).
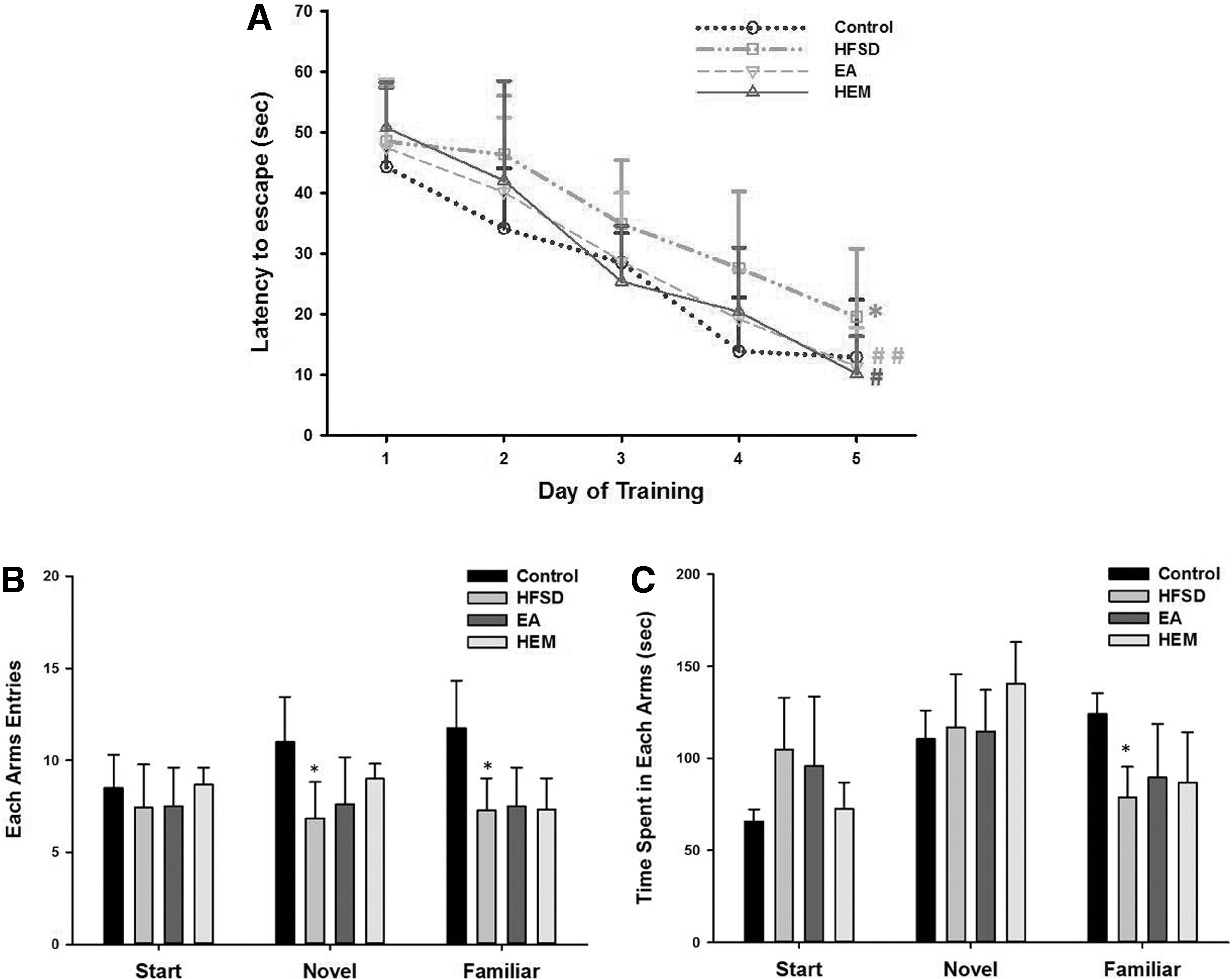
Effects of HEM and EA treatments on MWM and Y-maze performance in aging mice fed with HFSD.
In the memory test, the HFSD group showed a significantly decreased number of novel arm entries (Fig. 2B) and spent more exploratory time in the start arm compared with the control group (Fig. 2C), although the exploratory times were not significantly different. Interestingly, the HEM-treated group showed a trend toward an increasing number of novel arm entries (Fig. 2B) and a slightly shortened exploratory time in the start arm (Fig. 2C) compared with the HFSD group. Although the performances in the Y-maze were not significantly different, the results from the MWM assessments revealed that EA and HEM may play roles in improving spatial learning in aging mice fed with an HFSD.
Effects of HEM and EA on HFSD-induced neuroinflammation in the hippocampus
As HEM and EA improved the performance of aging mice fed with the HFSD in the MWM and Y-maze assessments (Fig. 2), the effects of HEM extract and EA on reduction of HFSD-induced neuroinflammation were investigated. The hippocampal mRNA expression levels of TNFα and IL-1β, which are primary proinflammatory factors, were analyzed by RT-PCR. Consistent with previous reports, the mRNA expression levels of TNFα and IL-1β were elevated in aging mice fed with the HFSD compared with the chow diet control group (Fig. 3). More specifically, the IL-1β levels were markedly increased compared to those in the control group (Fig. 3B). Notably, the administration of HEM and EA resulted in a reduction in the mRNA expression levels of TNFα and IL-1β (Fig. 3A, B) compared to those in the group of aging mice fed with the HFSD. Interestingly, the effect of EA was found to be more significant compared with HEM on the mRNA expression of both proinflammatory factors (Fig. 3A, B).

Effects of HEM and EA on HFSD-induced neuroinflammation in the aging mouse hippocampus. The relative mRNA abundance of TNFα
Effects of HEM extract and EA on mRNA expression levels of NGF and NeuN in the hippocampus
To determine whether HEM and EA improve learning abilities impaired by the HFSD regimen, we investigated whether treatment with HEM and EA can influence the expression of NGF mRNA in the hippocampus, which is critically related to the processes of learning and memory in the brain. RT-PCR analysis demonstrated that the expression of NGF mRNA was decreased twofold in aging mice fed with the HFSD compared to that in the control group of mice (Fig. 4A). However, the expression of NGF mRNA was significantly upregulated, nearly to the level of control mice, when aging mice fed with the HFSD were supplemented with HEM extract (Fig. 4A). NGF has been demonstrated to enhance the survival of new neurons in the granule cell layer of the dentate gyrus. 26 To further confirm whether the observed elevation of NGF mRNA after treatment with HEM and EA can decrease neuronal loss in the hippocampus of aging HFSD-fed mice, NeuN mRNA levels were monitored in the four groups. NeuN, an important marker of matured neurons, decreases under several pathological conditions as well as during aging, 27 which adversely affects neuronal viability. As expected, the expression of NeuN mRNA was significantly decreased in the hippocampus of the aging mice fed with the HFSD compared to that in the control group (Fig. 4B). However, upon treatment with HEM and EA in the mice fed with the HFSD, increased NeuN mRNA expression in the hippocampus was observed (Fig. 4B). Finally, treatment with HEM markedly increased the expression of NeuN mRNA by more than twofold compared with that in the HFSD group. In addition, as shown in Figure 4B, HEM could rescue the NeuN mRNA level to the control level and was more effective than EA.
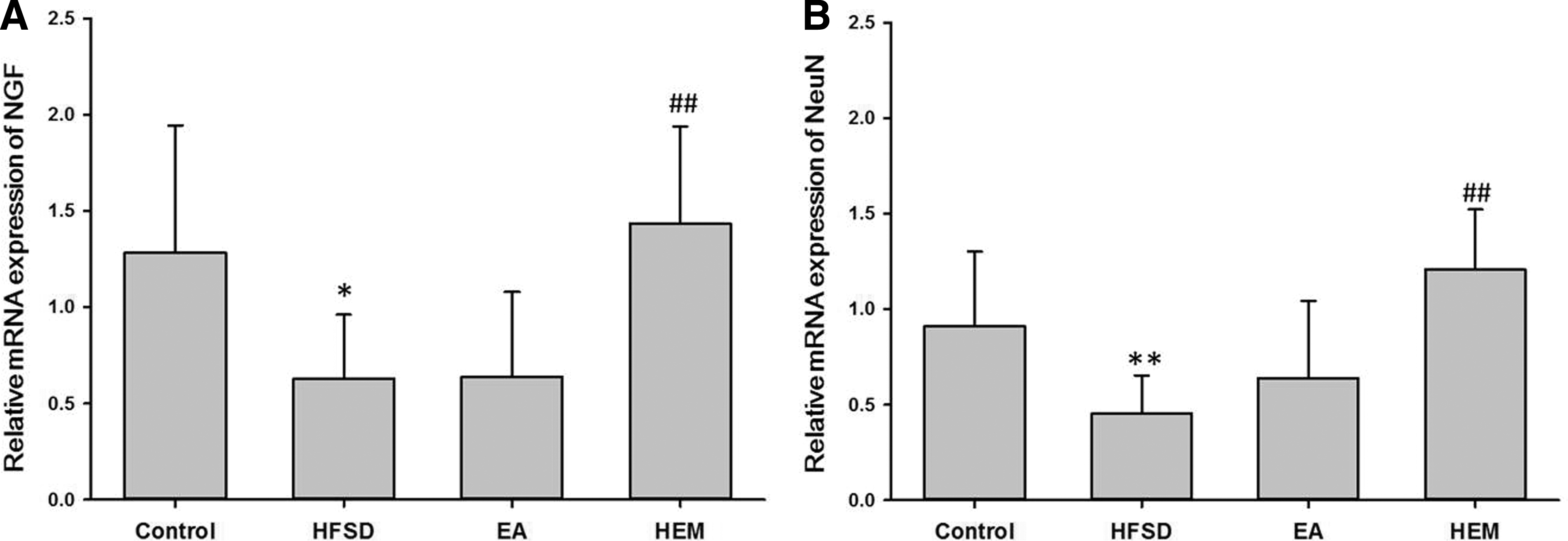
Effects of HEM and EA on neuroprotection in the aging mouse hippocampus. Relative mRNA abundance of NGF
Effects of HEM and EA on total body weight and fat pad weight in aging mice fed with the HFSD
Consumption of an HFSD has been found to be associated with obesity and poor cognition. 28 Consistent with this report, this study had found that the body weights in the HFSD group were significantly higher than those in the control group (Table 1). Interestingly, HEM and EA treatment in the aging mice fed with the HFSD for 18 weeks significantly suppressed body weight gain, although these mice were still slightly heavier than the control mice (Table 1).
Effects of Hericium erinaceus Mycelium and Erinacine A on Body Weight and Fat-Pad Weight of Aging Mice Fed with the High-Fat and High-Sucrose Diet for 18 Weeks
Each value represents the mean ± SD (n = 8–10 for each group).
See the footnote text of Table 1 for a detailed description of the experimental groups.
P < .05 and ** P < .005 compared with the control group.
P < .05 and ## P < .005 compared with the HFSD group.
CRE, creatinine; EA, erinacine A; GOT, glutamate oxaloacetate transaminase; GPT, glutamate pyruvate transaminase; HEM, Hericium erinaceus mycelium; HFSD, high-fat and high-sucrose diet; SD, standard deviation; TC, total cholesterol; TG, triacylglycerol; UA, uric acid.
Furthermore, the fat pad weights in the four groups were also evaluated (Table 1). The results showed that increases in mesenteric and perirenal fat pad weights were significantly higher compared to those in the control group. Notably, both HEM and EA treatment reduced the fat pad weights in the aging mice fed with the HFSD. In addition, the weights of the mesenteric fat pads in the HEM group decreased significantly and were similar to those in the control group.
Effects of HEM and EA on biochemical parameters in aging mice fed with the HFSD
As shown in Table 2, the HFSD caused elevated serum glucose, serum TC, liver TG, and TC concentrations compared to the control diet. Among the biochemical parameters measured, the serum TC, liver TG, and liver TC increased significantly in the HFSD group compared with those in the control group (Table 2). Similarly, GPT and UA levels in the HFSD group were increased significantly compared with those in the control group (Table 2). Importantly, HEM treatment significantly rescued the levels of serum glucose, TC, GOT, GPT, and UA nearly to the levels of the control mice (Table 2). EA treatment also significantly decreased the levels of serum glucose, TC, and GPT, but not GOT. Since the HEM and EA treatments markedly reduced the levels of GOT and GPT, the levels of liver TG and TC were also measured. HEM treatment significantly reduced the levels of liver TG and TC nearly to the levels of the control mice (Table 2, P < .005). EA treatment also reduced the levels of liver TG and TC, but only the liver TG level was significantly different (Table 2, P < .05).
Effects of Hericium erinaceus Mycelium and Erinacine A on Biochemical Parameters in Aging Mice Fed with the High-Fat and High-Sucrose Diet for 18 Weeks
Each value represents the mean ± SD (n = 8–10 for each group).
See the footnote text of Table 1 for a detailed description of the experimental groups.
P < .05 and ** P < .005 compared with the control group.
P < .05 and ## P < .005 compared with the HFSD group.
CRE, creatinine; GOT, glutamate oxaloacetate transaminase; GPT, glutamate pyruvate transaminase; TC, total cholesterol; TG, triacylglycerol; UA, uric acid.
Discussion
Short-term memory loss and cognitive impairment are commonly observed among individuals experiencing senile dementia. In this study, consumption of an HFSD for 18 weeks was found to cause deterioration of learning ability as well as elevated levels of inflammatory factors in the hippocampus of aging mice. Using this model, HEM extract and the mycelium-derived compound EA were demonstrated to facilitate the rescue of cognitive functions in aging mice fed with the HFSD as revealed in the MWM (Fig. 2). Furthermore, anti-inflammatory effects (Fig. 3) and mitigation of neuronal death through the NGF (Fig. 4) mechanism, as well as the effects of HEM and EA on biochemical parameters in the aging mice fed with the HFSD were also investigated (Table 2).
To induce obesity, 15-month-old mice were given an HFSD rich in saturated fats and refined sugar, similar in composition to the average popular diet of most industrialized Western societies. In this diet-induced-obesity model, the mice displayed deterioration of spatial learning as assessed by MWM (Fig. 2A); however, this effect was reversed by the addition of HEM or EA to the HFSD. These MWM assessments demonstrated that both EA and HEM may play roles in improving spatial learning in aging mice fed with an HFSD.
Recently, systematic studies and meta-analyses have revealed that brain proteomic modifications are associated with high-fat-diet-induced obesity in murine models, for example, calcium signaling, oxidative phosphorylation, glycolysis/gluconeogenesis, and hypoxia-inducible factor signaling. 29 Changes in protein expression and signaling pathways have been reported to be associated with brain dysfunction. 30 –32 Another major factor linking obesity and cognitive impairment is neuroinflammation. In this study, the 15-month-old mice fed with the HFSD showed increased mRNA expression levels of TNFα and IL-1β in the hippocampus, which is consistent with other previous studies reporting that obesity can lead to neuroinflammation. 33 Importantly, the addition of HEM or EA to the HFSD ameliorated the elevated neuroinflammation in the mouse hippocampus. These results may imply that EA and HEM improve spatial learning, as observed in the MWM test, by decreasing neuroinflammation. Interestingly, the EA group showed better learning performance in the MWM test (significant, P < .005) and decreased expression levels of both TNFα and IL-1β, while the HEM group showed improved spatial learning (significant, P < .05) and decreased expression of only TNFα. The different effects of EA and HEM are speculated to be caused by interference from other components of HEM.
Notably, decreased mRNA expression levels of NGF and NeuN were observed in the aging mice that consumed the HFSD for 18 weeks, but the addition of HEM reversed these harmful effects, even causing these levels to be slightly higher than those of the control mice (Fig. 4). NGF plays an important role in neuronal plasticity and survival. Recent studies have demonstrated that NGF triggers the ERK/CREB signaling pathway, which is involved in the regulation of protein synthesis required for synaptic remodeling and memory formation. 34,35 High levels of NGF decrease neuronal death and improve learning and memory. 36 Intracerebroventricular infusion of NGF can increase synaptogenesis in the rodent hippocampus. 37 Considering that NGF is important for cognitive functions and decreases with aging, it may contribute to an age-dependent decline in cognitive function. However, NGF is a high-molecular-weight protein that cannot cross the blood–brain barrier (BBB). Hericenones and erinacines extracted from HEM have been reported to have the ability to cross the BBB and promote NGF synthesis in cultured astrocytes. 38 Thus, HEM can be used as a functional food to enhance NGF expression and improve impairment in the aging brain due to HFSD consumption.
In this study, body weight and abdominal fat were significantly higher in the aging mice that consumed the HFSD than those in the aging mice that consumed the chow diet (Table 1). A recent study reported that patients with AD had more abdominal fat than patients without AD. 39 Adipose tissue is the largest endocrine organ in the body, and adipocytes secrete adipocytokines including leptin, IL-1, IL-6, and TNFα, which have been reported to be associated with cognitive impairment. 40 –43 These inflammatory cytokines derived from adipose tissue reach the cerebral microcirculation through the bloodstream, alter the function of cerebromicrovascular endothelial cells, and may disrupt the BBB. 44,45 BBB damage is thought to promote neuroinflammation and oxidative stress, which are associated with neurodegeneration. 44,46 In this study, both neuroinflammation (Fig. 3) and abdominal fat (Table 1) were significantly reduced in the aging mice that consumed the HFSD with HEM or EA. These results imply that the reversal of neuroinflammation by HEM and EA is associated with a decrease in abdominal fat. However, the detailed mechanism requires further elucidation.
Consumption of an HFSD induces not only obesity but also hyperlipidemia. 47 According to our data, the HFSD group had a threefold higher serum cholesterol concentration compared with the control group (Table 2). A previous report indicated that upon elevation of the cholesterol concentration in the serum, the cholesterol concentration in the brain will also increase. 48 Excess cholesterol must be eliminated from the central nervous system because of its potential toxic effects on neurons. Cholesterol is converted to 24-hydroxycholesterol by 24-hydroxylase before being transported out of the brain. Epidemiological studies have indicated that 24-hydroxycholesterol concentrations in the cerebrospinal fluid are increased in AD patients. 49 Evidence from earlier studies has also shown that high cholesterol-induced neuroinflammation is correlated with loss of working memory. 50 In this study, both serum cholesterol (Table 2) and inflammatory factors in the hippocampus (Fig. 3) were elevated in the HFSD group compared with those in the control group. Furthermore, GOT and GPT levels in the aging mice that consumed the HFSD were higher than those in the aging mice that consumed the chow diet, indicating that consumption of an HFSD resulted in liver dysfunction in aging mice (Table 2). We also observed higher liver TG and liver TC in the aging mice given the HFSD (Table 2). The results indicated that the excess fat accumulated in the liver. Previous studies have demonstrated that HFSD-induced obesity is often accompanied by nonalcoholic fatty liver disease and hepatic meta-inflammation. 51 Nonalcoholic fatty liver disease impairs hippocampal-dependent memory function and interferes with hippocampal neurogenesis. 9 However, nonalcoholic fatty liver disease can be improved by modulating the MAPK-JNK/p38 signaling pathway. 52 HEM and EA have also been reported to decrease neuronal cell death by inhibiting the MAPK-JNK/p38 signaling pathway in an ischemia-injury model. 53 Thus, HEM and EA may ameliorate nonalcoholic fatty liver disease through the MAPK-JNK/p38 signaling pathway to rescue hippocampal-dependent memory function.
This study corroborates earlier reports indicating that consumption of an HFSD resulted in obesity, increased serum cholesterol, elevated TG and TC in the liver, and impaired cognitive function in aging mice. However, the aforementioned conditions can be reversed by the addition of HEM and EA to the HFSD (Tables 1 and 2). Interestingly, both the EA- and HEM-containing diets ameliorated spatial learning deficits in the aging mice; however, their efficacy may vary. EA had a more pronounced inhibitory effect on neuroinflammation. On the other hand, HEM had a significant promoting effect on NGF biosynthesis, thus reducing neuron loss, abdominal fat, and metabolic abnormalities. The results indicated that the protective effect of HEM against HFSD-induced brain impairment involved multiple metabolic mechanisms. HEM has been reported to improve lipid metabolism by the activation of peroxisome proliferator-activated receptor alpha. 54 Although EA was not able to promote NGF biosynthesis in our animal model, which is inconsistent with previous studies, the discrepancy may be caused by the different models or extraction processes used. However, several chemical constituents can be isolated from HEM, and further isolation and identification of other efficacious elements that may contribute to these biological effects are warranted. 55
In conclusion, aging mice that consumed an HFSD supplemented with EA or HEM exhibited a reduced inflammatory response, decreased neuronal loss and higher NGF biosynthesis, and performed better in the spatial learning tests compared to the mice that did not consume an EA- or HEM-containing diet. Furthermore, these findings demonstrated that consumption of an HFSD with EA or HEM can significantly reduce metabolic abnormalities, including obesity, higher serum cholesterol and TG and TC in the liver. The improved metabolic conditions and parameters also benefit the cognitive function of aging mice fed with an HFSD, suggesting that EA- and HEM-containing diets may be beneficial for preventing metabolic syndrome-related cognitive impairment. Thus, the mycelium of H. erinaceus and its isolated compound, EA, improved spatial learning deficits by reversing neuroinflammation and neuronal loss in aging mice fed with the HFSD, reflecting this mushroom's potential to be developed into nutritional supplements and adjuvant drugs for the treatment of age- and obesity-related neurodegeneration.
Footnotes
Acknowledgments
The authors gratefully acknowledge the Taiwan Mouse Clinic (Ministry of Science and Technology [MOST] 105-2325-B-001-010), which is funded by the National Research Program for Biopharmaceuticals at the MOST of Taiwan, for technical support in serum chemistry analyses. This work was supported by Ministry of Science and Technology (MOST 102-2632-M-033-001-MY3 and MOST 103-2632-M-033-001-MY3), Chung Yuan Christian University, Taiwan, Republic of China.
Author Disclosure Statement
The authors declare no conflicts of interest in this study.