
Abstract
Glyceollins are soybean-derived phytoalexins that induce the nuclear factor (erythroid-derived 2)-like 2 (Nrf2) signaling pathway, which is involved in the detoxification of carcinogens and the removal of reactive oxygen species (ROS). Recent studies, however, have indicated that Nrf2 induction stimulates the development of pre-existing tumors and confers resistance to chemotherapy by elevating drug metabolism and by efficient scavenging of ROS produced by the Warburg effect, which is regulated, in turn, by the p53 tumor suppressor. This study, therefore, aimed at examining whether glyceollins could accelerate tumor growth in the presence of active p53, using a xenograft BALB/c nude mouse model transplanted subcutaneously with p53 wild-type and p53 null HCT116 human colon cancer cells. Glyceollins were orally administered at a dose of either 1 or 4 mg/kg body weight after xenografting HCT116 cells, and tumor growth and volume were monitored for 2 weeks. A high dose of glyceollins resulted in a significant increase in the average volume of p53 wild-type HCT116 xenografts, but not of p53 null HCT116 xenografts. However, a low dose of glyceollins had no effect on the tumor growth regardless of p53 presence. Interestingly, antioxidant enzymes, including heme oxygenase-1 (HO-1) and NAD(P)H: quinone oxidoreductase, were prominently induced by glyceollins in p53 wild-type xenografts, compared with p53 null xenografts. These results suggest that a high dose of glyceollins possibly promotes the growth of p53 wild-type colon cancer through activation of the Nrf2-mediated signaling pathway and, in particular, strong induction of HO-1 expression. Therefore, the consumption of Nrf2 activators, including glyceollins, should be carefully monitored for patients suffering from certain types of cancer and/or receiving chemotherapy.
Introduction
Nuclear factor (erythroid-derived 2)-like 2 (Nrf2) is a transcription factor that responds to cellular stimuli by directing various transcription programs, including cellular stress signals. 1 Studies have shown that Nrf2 activation is associated with cytoprotection against reactive oxygen species (ROS) and detoxification in normal cells by inducing anti-oxidative genes, such as heme oxygenase-1 (HO-1), glutathione reductase (GR), NAD(P)H: quinone oxidoreductase-1 (NQO1), 2 phase 2 detoxifying enzymes, and anti-inflammatory proteins. Enhancement of Nrf2 activity in malignant cells, however, could protect tumor cells against the cytotoxicity of high endogenous ROS levels, thereby leading to increased resistance to chemotherapy and increased survival of tumor cells. The beneficial effects of activating the Nrf2 signaling pathway in cancer prevention have, thus, become a controversial issue in cancer intervention. 3,4
Although p53 is a tumor suppressor and guardian of the genome, its mutation leads to loss of wild-type p53 activity, which is frequently found in many different tumor types. p53 and Nrf2 co-regulate each other both positively and negatively. 5 It has been reported that p21, a p53 target protein, binds to Kelch-like ECH-associated protein 1 (Keap1) and eventually stabilizes Nrf2 by blocking proteasome degradation. NQO1, a major downstream protein of Nrf2 transcription factor, also reportedly activates Nrf2 by interacting with p53, thereby blocking proteosomal degradation of p53. In addition, there is evidence suggesting that p53 binds the promoters of genes activated by Nrf2 and represses Nrf2-downstream genes. 5,6
Glyceollins, which are soybean-derived phytoalexins, are synthesized de novo and accumulate in soybean after exposure to environmental stresses, such as microbial attack, fungal infection, physical damage, or ultraviolet light radiation. 7 These compounds have three major isomeric forms (glyceollin I, II, and III), and they are derived from the precursor daidzein. 8 Glyceollins were reported to have various beneficial biological functions, including anti-inflammatory activities, 9 antioxidant properties, 10 and anti-carcinogenic and estrogenic effects. 11,12 In addition, this group of compounds exhibits a range of toxic effects against fungi, insects, and bacterial disease. In particular, glyceollins were reported to activate the Nrf2 signaling pathway, including HO-1 in tumor cells, 13 which could promote tumor growth through efficient scavenging of ROS produced by the Warburg effect. Further, recent studies indicated that carbon monoxide (CO) produced by HO-1 activity could have a variety of biological effects such as mitochondrial biogenesis, anti-inflammatory activity, and cytoprotection, thus favoring tumor cell survival and growth. 14 –16
In this study, we investigated whether glyceollins could promote tumor growth in a xenograft mouse model and whether this ability was affected by the status of p53.
Materials and Methods
Cell culture
The human colorectal cancer p53 wild-type HCT116 cell line was obtained from the Korean Cell Line Bank (Seoul, Korea). The p53 null HCT116 cell line was a generous gift from Prof. Young Ho Kim (Kyungpook National University, Daegu, Korea; originally provided by Dr. Bert Vogelstein, Johns Hopkins University, Baltimore, MD, USA). Both cell lines were maintained in Dulbecco's modified eagle's medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (Welgene, Gyeongsan, Korea) and 1% penicillin-streptomycin (100 U/mL penicillin and 100 μg/mL streptomycin; Welgene). The cells were cultured at 37°C in a CO2 incubator (MCO-19 AIC; Sanyo, Osaka, Japan) with 5% CO2/95% air with 100% humidity. For sub-cultures, the cells that reached about 80% confluency were rinsed in phosphate buffered saline (PBS), detached by using 0.05% trypsin-EDTA, harvested, and plated onto culture dishes (all from Thermo Fisher Scientific).
Animals
Animal experiments were conducted according to the guidelines of the Committee on Care and Use of Laboratory Animals of the Kyungpook National University (Approval Number 2016-0142). BALB/c nude mice (male, 6 weeks old, 17–19 g of body weight, BW) were purchased from Orient Bio, Inc. (Seongnam, Korea). All animals were housed under standard conditions (temperature, 22°C ± 2°C; humidity, relative humidity 45% ± 5%; 12-h light/dark cycles) during the whole experimental period. Mice were allowed free access to drinking water and chow pellets (Daehan BioLink, Eumseong, Korea).
Experimental design
After an adaption period of 1 week, p53 wild-type (p53+/+) and p53 null (p53−/−) HCT116 cells (2 × 10 6 cells per injection) were resuspended in 100 μL of a mixture of PBS and Matrigel (Corning, Inc., Corning, NY, USA) at a ratio of 1:1 (v/v), and they were injected subcutaneously on the left and right flanks of the same mouse, respectively (Fig. 1A). A week after cell transplantation, mice bearing the xenografted tumors were divided randomly into three groups (n = 5–6) and the day was set as day 0; group 1 (referred to as “Control”) received the vehicle (composed of 0.5% v/v tween-80, 0.5% v/v absolute ethanol, and 99% sterilized drinking water), group 2 (referred to as “Gly_Low”) received a low dose of glyceollins at 1 mg/kg BW, and group 3 (referred to as “Gly_ High”) received a high dose of glyceollins at 4 mg/kg BW (Fig. 1B).
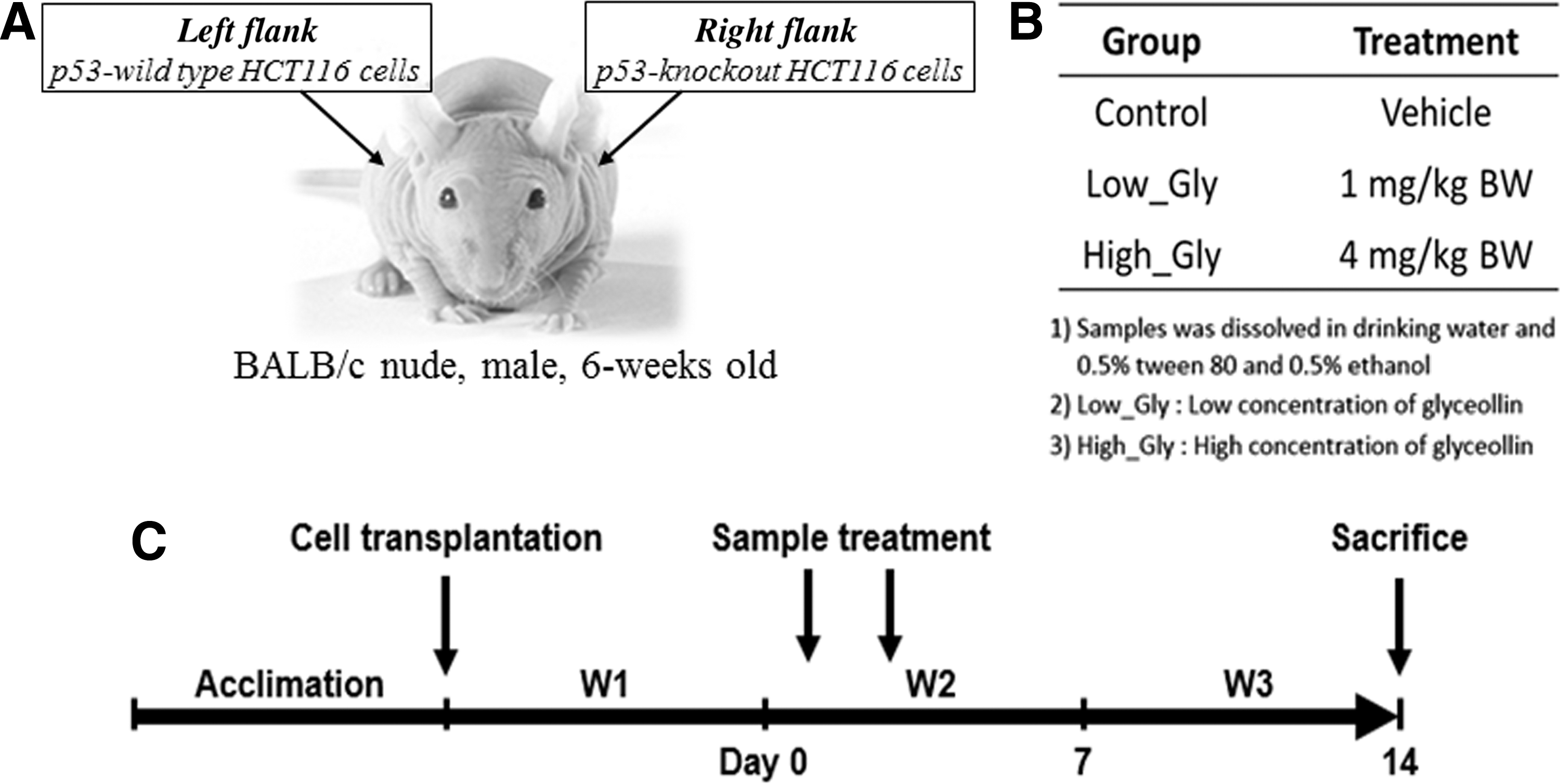
Experimental design.
For treatment, glyceollins were dissolved in the vehicle and orally administered twice at days 1 and 3. At termination of the experiment (at day 14), mice were sacrificed (Fig. 1C) and the xenografted tumor tissues were dissected for further biochemical analyses.
Measurement of BW and tumor growth
The BW of mice was recorded thrice per week. The tumor volume was estimated by measuring the width and length of each tumor using a caliper (Mitutoyo Corp., Kawasaki, Kanagawa, Japan) and calculated according to the formula: tumor volume (mm 3 ) = (L × W 2 )/2, where L = length in mm, and W = width in mm. The tumor tissues dissected from the sacrificed mice were weighed and frozen at −80°C for subsequent assays, including western blot.
Extraction of total proteins
The total proteins were obtained from each tumor tissue by homogenizing in pre-cooled lysis buffer (20 mM Tris-Cl, 145 mM NaCl, 10% glycerol, 5 mM EDTA, 1% Triton X-100, and 0.5% Nonidet P-40), including 1 × EDTA-free protease inhibitor cocktail (Roche Diagnostics, Mannheim Germany) by using a micro-ultrasonic cell disrupter (KT50; Kimble Kontes, Vineland, NJ, USA). After centrifugation at 12,000 g for 10 min at 4°C, the amount of proteins in the supernatant was quantified by the Bradford assay. 17
Fractionation of nuclear and cytoplasmic proteins
The dissected tumor tissues were lysed by using NE-PER® Nuclear and Cytoplasmic Extraction reagents (Thermo Fisher Scientific) according to the manufacturer's instructions. The protein quantity in the extracted nuclear and cytoplasmic fractions was determined by the Bradford assay. 17
Western blotting analysis
The proteins extracted from the tumor tissues (30 μg) were quantified, denatured, electrophoretically separated on sodium dodecyl sulfate-polyacrylamide gels, and transferred onto polyvinylidene fluoride membranes (Millipore, Bedford, MA, USA) as previously described. 18 The primary antibodies used in this study were against Nrf2, HO-1 (from Abcam, Cambridge, UK), p53, GR, lamin B, and β-actin (from Santa Cruz Biotechnology, Santa Cruz, CA, USA). The secondary antibodies used were anti-rabbit or anti-mouse immunoglobulin G, conjugated to horseradish peroxidase (from Santa Cruz Biotechnology).
Assay of 8-hydroxy-2′-deoxyguanosine levels in tumor tissue
To examine the level of 8-hydroxy-2′-deoxyguanosine (8-OHdG), a biomarker for DNA damage, the frozen tumor tissues were thawed on ice, rinsed in cold PBS, homogenized by using a micro-ultrasonic cell disrupter, and, subsequently, centrifuged at 900 g for 15 min. After the quantity of total proteins in the supernatants was determined, the 8-OHdG level was assayed by using a commercially available DNA damage ELISA kit (Enzo Life Sciences, Inc., Farmingdale, NY, USA), according to the manufacturer's instructions.
Statistical analysis
Statistical significance was determined by one-way analysis of variance followed by post hoc Duncan's multiple-range test, using SPSS software (SPSS, Inc., Chicago, IL, USA). The P values <.05 (P < .05) were considered statistically significant.
Results
Expression of p53 protein in two types of HCT116 cells
Before the establishment of HCT116 xenograft mouse models by transplantation of both types of HCT116 cells, the p53 protein expression was confirmed by western blotting analysis. The expression of p53 was observed only in p53 wild-type (p53+/+) HCT116 cells but not in p53 null (p53−/−) HCT116 cells (Fig. 2). Based on this result, both types (p53 wild-type and p53 null) of HCT116 cells were subcutaneously injected into the left and right flanks of BALB/c nude mice, respectively.
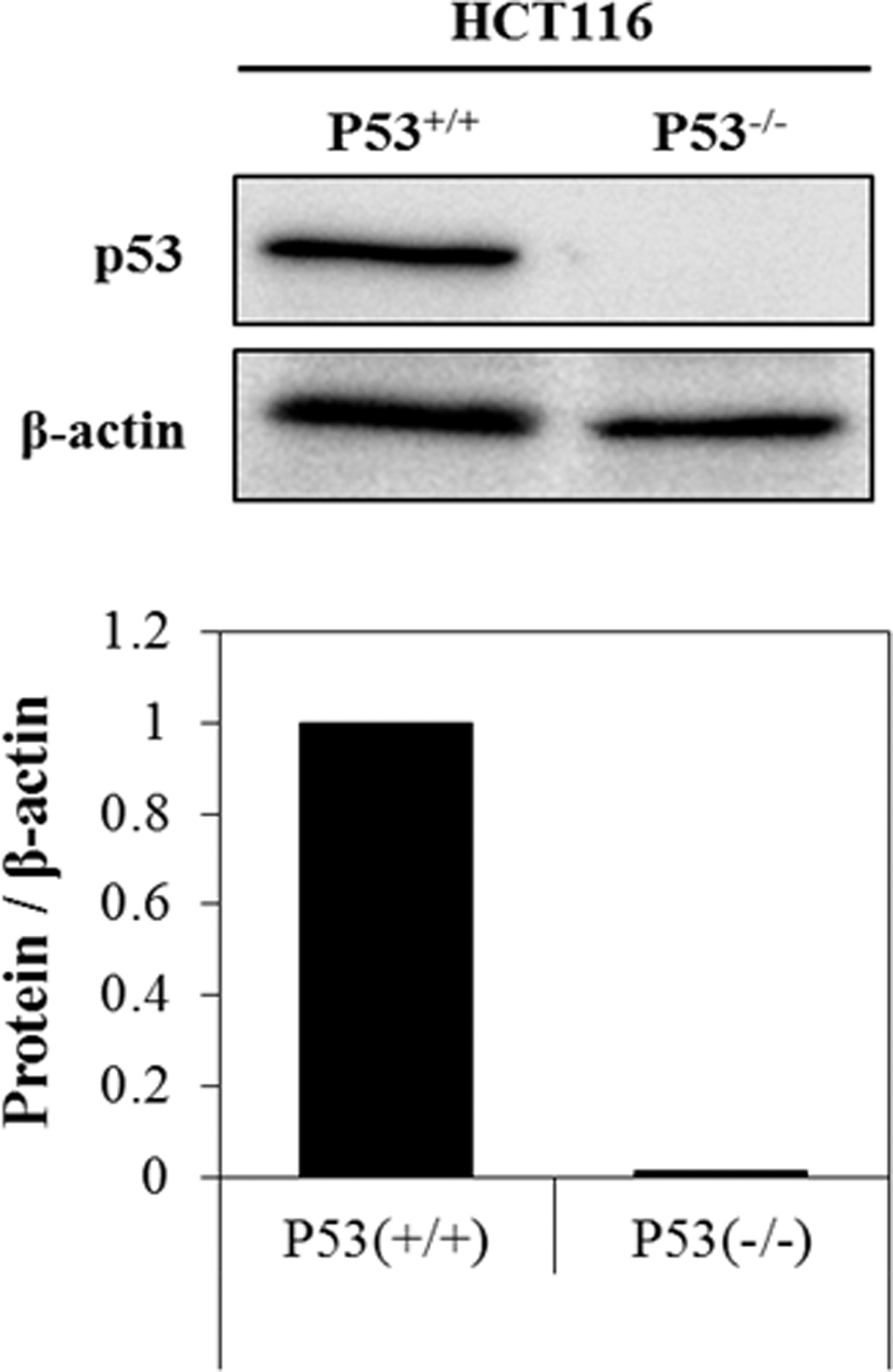
Expression of p53 protein in two types of HCT116 cells. The p53 proteins were expressed only in p53 wild-type HCT116 cells but not in p53 null HCT116 cells.
Effect of glyceollins on BW of BALB/c nude mice transplanted with two types of HCT116 cells
After injection, mice bearing the xenografted tumor were randomly assigned into three groups: (1) “control” (vehicle-treated; no glyceollins treatment), (2) “Gly_Low” (oral treatment with a low concentration of glyceollins at 1 mg/kg BW), and (3) “Gly_High” (oral treatment with a high concentration of glyceollins at 4 mg/kg BW). The average BW of each group was monitored on a regular basis during the entire experimental period. No significant difference in BW was observed among the three groups (Fig. 3).

Effect of glyceollins on BW of BALB/c nude mice transplanted with p53 wild-type and p53 null HCT116 cells. BALB/c nude mice bearing xenografted tumors on their flanks were divided into three groups (5–6 mice per group): Control (vehicle-treated), Gly_Low (glyceollins-treated at a low dose, 1 mg/kg BW), and Gly_High (glyceollins-treated at a high dose, 4 mg/kg BW). The BW of the mice was monitored thrice a week for 14 days. Data are shown as mean ± SD (n = 5–6). No significant difference in BW was observed among experimental groups. BW, body weight; SD, standard deviation.
Effect of glyceollins on the size, volume, density, and growth rate of xenografted tumors
At termination of the experiment, the xenografted tumors were dissected and measured for their volume and weight. No substantial difference in the apparent sizes of the xenografts derived from p53 wild-type and p53 null HCT116 cells was observed (Fig. 4A). The size and volume of p53 wild-type xenografts were significantly increased by treatment with a high dose of glyceollins (4 mg/kg BW), compared with those of control (vehicle-treated) or of p53 wild-type xenografts treated with a low dose of glyceollins (1 mg/kg BW) (Fig. 4A, B). On the other hand, p53 null xenografts exhibited a similar size and volume to the vehicle-treated control, and their volumes were not affected by treatment with either low or high doses of glyceollins (Fig. 4A, B).

Effect of glyceollins on the size, volume, density, and growth rate of xenografted tumors. The mice bearing xenografted tumors were orally administered glyceollins twice at days 1 and 3. After termination of the experiment at day 14, the mice were sacrificed and the xenograft tumors were dissected and measured for their size and weight.
In addition, treatment with glyceollins significantly increased tumor density of p53 wild-type xenografts, but not of p53 null xenografts (Fig. 4C). A similar trend was observed for tumor growth rate; that is, p53 wild-type xenografts in mice treated with a high dose of glyceollins displayed a significantly higher growth rate than the other experimental groups (Fig. 4D).
Effect of glyceollins on nuclear accumulation of Nrf2 in the liver of glyceollins-treated mice
Nrf2 signaling is responsible for the expression of and phase 2 antioxidant and detoxifying enzymes. 19 The Nrf2-Keap1 complex is separated by glyceollins in the cytosol, with the released Nrf2 migrating into the nucleus where it binds the antioxidant response element sequence of phase 2 antioxidant/detoxifying enzymes, thereby activating their transcription. 13 Consistent with the previous findings, the result from this study demonstrated that both cytoplasmic and nuclear Nrf2 levels were enhanced in liver tissue of mice treated with both low and high doses of glyceollins (Fig. 5A).

Effect of glyceollins on nuclear translocation of Nrf2 in the liver and expression of Nrf2-dependent enzymes in xenografted tumor tissues. Cytoplasmic and nuclear Nrf2 levels in the liver tissues obtained from the experimental mice
Effect of glyceollins on expression of Nrf2-dependent enzymes in xenografted tumor tissue
The Nrf2-dependent antioxidant/detoxifying enzymes, such as HO-1, NQO1, and GR, have the potential to promote tumor growth through cytoprotective effects from ROS in cancer cells. 3,20 We, thus, examined whether glyceollins could induce the expression of phase 2 detoxifying enzymes, including HO-1, NQO1, and GR, in the xenografted tumor tissue. Indeed, a high dose of glyceollins significantly increased the expression of HO-1 and NQO1 in the p53 wild-type xenografts; however, it did not affect the expression in the p53 null xenografts (Fig. 5B). In addition, a low dose of glyceollins caused a slight but significant increase in NQO1 expression in p53 null xenografts (Fig. 5B).
Moreover, consistent with our previous studies, 13,21 in vitro results demonstrated that glyceollins significantly increased the nuclear Nrf2 levels in both types of HCT116 cells in culture (Supplementary Fig. S1). Interestingly, the mRNA levels of NQO1 and HO-1 were significantly upregulated only in p53 wild-type HCT116 cells but not in p53-null HCT116 cells (Supplementary Fig. S2). These findings comprehensively suggest that glyceollins may promote the p53-dependent cytoprotective effect besides Nrf2-mediated antioxidant response in the xenografted tumors.
Effect of glyceollins on 8-OHdG levels in xenografted tumors
To assess whether glyceollins caused DNA damage in xenografted tumor tissues, the 8-OHdG levels were measured in the xenograft homogenates. In p53 wild-type xenografts, there was no significant difference in the level of 8-OHdG between the control and the glyceollins-administered groups (Fig. 6A). However, the 8-OHdG levels were increased in p53 null xenografts by glyceollin treatment (Fig. 6B). These findings indicated that glyceollin treatment caused DNA damage, highly correlated with cell death, in p53 null xenografts rather than in p53 wild-type xenografts, which was consistent with a greater tumor volume and a higher tumor growth rate of p53 wild-type xenografts than those of p53 null xenografts.

Effect of glyceollins on 8-OHdG levels in xenografted tumors. The 8-OHdG levels in p53 wild-type xenografts
Discussion
Nrf2 is widely recognized to be associated with cancer prevention. However, recent studies suggest that Nrf2 activation could, in fact, further promote the development of existing tumors and induce drug resistance to chemotherapy. 3,20 It has been reported that cancer cells produce high levels of ROS due to increased metabolic activity, mitochondrial dysfunction, and peroxisome activity; increased cellular receptor signaling and oncogene activity; and increased activity of oxidases, cyclooxygenases, lipoxigenases, and thymidine phosphorylase, or through crosstalk with infiltrating immune cells. 22 –24 To secure cell survival, cancer cells usually develop the ability to overexpress antioxidant enzymes, which, in turn, are regulated by Nrf2. Thus, it is highly feasible that the activation of Nrf2 signaling can facilitate tumor survival and development through efficient removal of ROS, which would otherwise have contributed to the suppression of tumor growth.
As expected, high doses of glyceollins (4 mg/kg BW), an Nrf2 activator, were shown to accelerate the growth of HCT116 human colon cancer cells carrying wild-type p53 xenografted into BALB/c nude mice. Conversely, tumor growth of p53 null xenografts was not affected by glyceollins, which is suggestive of a crosstalk between Nrf2 signaling and p53. p53 is known to activate apoptosis, whereas its direct downstream target protein, p21, is able to interact with a motif of Nrf2, thereby attenuating Keap1-mediated ubiquitination and subsequent proteosomal degradation. 25,26 That is, p53 associated with the function of p21 could further augment the Nrf2 signaling activation induced by glyceollins, leading to the increased survival of cancer cells by lowering ROS level within tumor tissue. Consistent with this rationale, our data demonstrated that glyceollin treatment significantly increased the volume, weight, and growth rate of p53 wild-type xenografts but not those of p53 null xenografts. Moreover, the expression levels of inducible antioxidant enzymes HO-1 and NQO1, which are downstream proteins of Nrf2, were significantly enhanced in p53 wild-type xenografted tumor tissues when a high dose of glyceollins was administered. In contrast, p53 null xenografted tumor tissues showed limited change in antioxidant enzyme expression in response to glyceollins, although NQO1 expression was enhanced by a low dose of glyceollins. These findings suggest that induction of antioxidant enzymes by a high dose of glyceollins promoted tumor growth, likely through efficient elimination of ROS produced in tumor tissues.
CO, a gaseous molecule produced from the enzymatic reaction of HO-1 of which expression was prominently induced by glyceollins, has emerged as a versatile player in determining cell fate and modulating critical biological processes, including autophagy, mitochondrial biogenesis, programmed cell death, cellular proliferation, inflammation, and innate immune responses. 27,28 Low-dose CO has demonstrated cytoprotective and tissue-protective effects in animal models of organ injury and disease. Therefore, it is also possible that the xenografted tumor growth by glyceollins could have been facilitated via a CO-mediated mechanism promoted by HO-1 activity. However, the detailed mechanism of CO-mediated cytoprotection causing tumor growth needs further elucidation.
One of the interesting findings was that glyceollin treatment increased the 8-OHdG level, an indicator of DNA damage, in the p53 null xenografts but not in p53 wild-type xenografts. Considering that DNA damage is usually prevented or repaired through the function of p53, which is the guardian of the genome, glyceollins are assumed to cause DNA damage in the absence of p53. This observation was supportive of the tumor growth only in p53 wild-type xenografts, but not in p53 null xenografts. However, it remains to be clarified whether the glyceollins-induced DNA damage is limited to the tumor cells or can occur in normal cells as well.
In conclusion, Nrf2 activators, including glyceollins, may act as tumor promoters in cancer patients, although they remain useful chemopreventive agents in healthy people. Further studies that are more extensive, however, are required to confirm that Nrf2 activators exert tumor promotion activity, in general, as they usually interact with multiple targets within the cell.
Footnotes
Acknowledgments
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Ministry of Science and ICT (MSIT), Republic of Korea (Project No. 2017R1A2B4005087).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.