
Abstract
The leaves of Aster glehni Fr. Schm. (Asteraceae) have been used to treat insomnia in Korea. Insomnia is a common adverse effect of therapeutic agents for Alzheimer's disease (AD), and the control of sleep disturbance may prevent dementia. We hypothesized that the leaves of A. glehni can attenuate cognitive dysfunctions observed in AD. We observed the ameliorating effects of the ethanolic extract of leaves of A. glehni (AG-D) on memory dysfunction through the Morris water maze test, the passive avoidance test, and the Y-maze test. We performed acetylcholinesterase (AChE) activity assay and Western blotting to determine the mechanism of action of AG-D. AG-D significantly attenuated memory dysfunction observed in the above behavior studies and inhibited the activity of AChE. AG-D also increased the levels of phosphorylation extracellular signal-regulated kinase (ERK), cAMP response element-binding protein (CREB), phosphatidylinositol 3-kinase (PI3K), protein kinase B (Akt), and glycogen synthase kinase 3β (GSK-3β) and the expression levels of brain-derived neurotrophic factor (BDNF) in the hippocampi. These results suggest that AG-D ameliorates memory impairments by AChE inhibition and activation of ERK-CREB-BDNF and PI3K-Akt-GSK-3β signaling pathways. Taken together, this study suggests that AG-D could be used as a potential treatment for cognitive dysfunction.
Introduction
Owing to the aging society in the world, Alzheimer's disease (AD), characterized by cognitive dysfunction with behavioral impairments, is a severe public health problem. 1,2 Although the exact etiology of AD is still unclear, several hypotheses have been suggested, including the cholinergic hypothesis, Tau protein hypothesis, β-amyloid protein hypothesis, and calcium hypothesis, 3 to explain the pathophysiology of AD.
The cholinergic hypothesis receives the most attention because the cholinergic neurotransmission system is vulnerable to β-amyloid protein, Tau protein, and calcium homeostasis, 4,5 and plays a key role in cognitive function. 6 Thus, the acetylcholinesterase (AChE) inhibitors donepezil, galantamine, and rivastigmine are prescribed to treat symptoms of AD. 7 However, those drugs induce many side effects, such as nausea, vomiting, insomnia, diarrhea, and headache, which can be inconvenient to AD patients. 8,9 These limitations underscore the urgent need for AChE inhibitors with fewer side effects for the treatment of AD. In view of this, we have been investigating such agents from traditional herbal materials based on ethnopharmacological knowledge.
Some herbal materials belonging to the genus Aster have been used as traditional medicines to treat insomnia and headache. Recently, it has been reported that Aster yomena Honda (Asteraceae), which is cultivated inland rather than on an island, has an inhibitory activity on AChE. 10 Aster glehni Fr. Schm. (Asteraceae), another species of the genus Aster, has the same Korean name as A. yomena. It is a region-specific edible plant in Korea. It is only distributed and cultivated in Ulleung-do Island, Republic of Korea. In traditional medicine, the leaves of A. glehni have been included in prescribed formulations as an antipyretic and analgesic agent through removal of the wind, as described in traditional Chinese medicine. 11 In addition, the leaves of A. glehni have also been used as a folk medicine for the therapy of insomnia or hypercholesterolemia in Korea. 12,13 Studies reported that rapid-eye-movement (REM) sleep is reduced in AD patients, while it is relatively preserved in normal aging. 14 Thus, slow activity on electroencephalography during REM sleep has been proposed as a biological marker of AD. 15 These specific REM sleep disturbances are characterized by cholinergic innervation degeneration in the forebrain and brainstem. 16 Others studies also reported that the severity of such sleep disturbance appears to parallel the severity of dementia. 17 Therefore, the control of sleep disturbances has also been reported to be a tool for the prevention of dementia. 18,19 Recently, the leaves of A. glehni were reported to have antioxidative, anti-inflammatory, sedative, and anticonvulsant effects. 11,20 Considering that A. glehni is of the same genus as A. yomena Honda (Asteraceae), we hypothesized that the plant may also inhibit AChE, which may improve cognition in AD. If the leaves of A. glehni are effective in ameliorating memory impairment, they could be applied to AD therapy or for attenuating the adverse effects of AD therapy, such as insomnia. According to our knowledge, no published studies have investigated the effects of the leaves of A. glehni on learning and memory functions.
In this study, we generated an ethanolic extract from the leaves of A. glehni. We then observed the ameliorating effects of the leaves of A. glehni on memory impairment caused by scopolamine in the Morris water maze test (MWM), the passive avoidance test (PAT), and the Y-maze test (Y-M). In addition, we also observed the enhancing effects of the leaves of A. glehni on memory function through the PAT. We employed an AChE activity assay and the Western blotting to verify the mode of action of the leaves of A. glehni.
Materials and Methods
Animals
We procured male CD-1 6-week-old mice (25–30 g) from the Orient Co., Ltd, a division of Charles River Laboratories (Gyeonggi-do, Korea). Each cage housed five animals. Animals were free to ingest feedstuff and water, and reared in a 12-h illumination/12-h nonillumination cycle (light on time 7:00–19:00) for 7 days in a room with temperature at 23°C ± 1°C and humidity of 60% ± 10% before the animal experiments. Animal care and drug treatment were implemented according to the guidelines issued by Kyung Hee University, Korea. In addition, experimental protocols were ratified by the Institutional Animal Care and Use Committee of Kyung Hee University [approval No. KHUASP (SE)-16-131].
Materials
All chemical materials were purchased from Sigma Chemical Co. (St Louis., MO, USA). Brain-derived neurotrophic factor (BDNF) was purchased from Abcam (Cambridge, United Kingdom). Other antibodies were purchased from Cell Signaling Technology, Inc., (Danvers, MA, USA) or Santa Cruz Biotechnology, Inc., (Santa Cruz, CA, USA). Anti-goat, anti-mouse, and anti-rabbit IgG antibodies were purchased from GeneTex, Inc. (Irvine, CA, USA). All materials were of the highest available qualities and obtained from common commercial sources. All drugs were made freshly before each test. Donepezil, scopolamine, and piracetam were dissolved in 0.9% saline solution. The extract of A. glehni leaves was dissolved in Tween 80 solution (10% v/v).
Standardization of the ethanolic extract of the leaves of A. glehni
The ethanolic extract of A. glehni leaves was donated by Koreaeundan Co. (Seongnam, Korea) and prepared as follows: in brief, leaves of A. glehni were collected from Ulleung-do Island in September 2016 and dried naturally. The 2kg smashed sample was extracted using 25 L of 70% ethanol in a fermenter (KFC-30L; Korea Fermentor) at 80°C for 4 h, while being stirred at 150 rpm. After the first extracts were collected, the second extractions were performed with 70% ethanol (20 L) at 80°C for 2 h. Then, 15% perlite was added to the mixture of first and second extracts, which was then filtered through filter paper (pore size of 20 mm) and concentrated to 20 Brix using a concentrator (R10028; Dooyoung Hi-Tech). After the addition of equal amount of dextrin to the concentrates, the mixtures were disinfected at 95°C for 1 h using the autoclave (AC-03; Jeio Tech). A solid granule was obtained by spray-drying and filtered through a 60-mesh sieve. The specimen was kept in the herbarium of the Koreaeundan Co. (specimen No. AG-013). For quality assurance, the final A. glehni extract was standardized by 3,5-dicaffeoylquinic acid (3,5-DCQA) based on high-performance liquid chromatography (Agilent 1100 series, Agilent G1365B MWD [Multiple wavelength detector], Santa Clara, CA, USA) at 330 nm (Supplementary Data). The content of the marker compound (3,5-DCQA) in the final A. glehni extract (called AG-D) was 2.4 mg/g, as shown in Supplementary Fig. S1.
Passive avoidance test
The PAT consisted of two trials: acquisition and retention, performed on 2 consecutive days. Two similar sized chambers (20 × 20 × 20 cm), nonilluminated and illuminated chambers with a 50-W bulb, were separated by a guillotine door (5 × 5 cm), as described elsewhere. 21 One hour before an acquisition trial, mice were treated with AG-D (30, 100 or 300 mg/kg, p.o.) or donepezil (5 mg/kg, p.o.). The control group received vehicle solution (10% Tween 80 solution). Thirty minutes before the acquisition trial, animals also administered vehicle solution (0.9% saline solution, i.p.) or scopolamine (1 mg/kg, i.p.). Animals were initially put in the illuminated chamber and the guillotine door was opened after 10 sec during the acquisition trial. After the mouse got into the nonilluminated chamber, the guillotine door was closed and a 0.5-mA electroshock (3 sec) was transmitted to the mouse. The interval between the acquisition trial and the retention trial was 24 h. Each mouse was located in the illuminated chamber as in the acquisition trial; the guillotine door was opened 10 sec after placing the mouse. The definition of latency was the time taken for each animal to get into the nonilluminated chamber after the door opened. The record of latency was up to 300 sec.
To elucidate the memory-enhancing effect of AG-D, 1 h before an acquisition trial, mice were treated with AG-D (30, 100, or 300 mg/kg, p.o.) or piracetam (200 mg/kg, i.p.) for positive control, as described elsewhere. 22 The control group received vehicle solution (10% Tween 80 solution). After the mouse got into the nonilluminated chamber, the guillotine door closed and a 0.25-mA electroshock (3 sec), subeffective electroshock, was transmitted to the mouse. The maximum record of latency was 600 sec. The other procedures were same as those described above.
Y-maze test
We used a maze with 3 arms (length of 40 cm, width of 3 cm, and height of 12 cm) in which the angle among each arm was 120° to conduct the Y-M. As described previously, 23 the walls and floors of Y-M were made with black plastic without being transparent. One hour before the test, mice were treated with AG-D (30, 100, or 300 mg/kg, p.o.) or donepezil (5 mg/kg, p.o.). The control group received vehicle solution (10% Tween 80 solution). Thirty minutes before the test, mice were also administered vehicle solution (0.9% saline solution, i.p.) or scopolamine (1 mg/kg, i.p.). Animals were initially placed in one arm of the Y-M, and all behaviors were recorded within 8 min using video camera. After each test, arms were cleaned by 70% ethanol sprayed onto the surface to remove residues and odors. The sequences and total numbers of arm entries were analyzed manually by another researcher who was ignorant of treatment condition. The definition of spontaneous alternation is entries into all arms on consecutive choices (i.e., BCA, ABC, or CBA, but not CAC). The alternation score (%) is shown as follows: percent of alternation = 100 × ([numbers of alternation]/[total numbers of arm entries–2]). The total number of arm entries was used to indicate locomotor activity of mouse. 24
Morris water maze test
The MWM was conducted in a dark room (50 Lux). The maze used for the test is a round tank (height of 45 cm and diameter of 90 cm) with four different visual cues on the wall. One hidden black platform (height of 29 cm and diameter of 6 cm) was placed in the center of a quadrant in the tank, which was conceptually divided. Water (24°C ± 1°C) was filled to the tank until 1 cm above the hidden black platform and dark dye was dissolved in water to prevent the platform from being visible. The MWM was performed over 7 days, as described elsewhere. 25 On the first experimental day, animals were allowed to swim freely for 60 sec in the tank for habituation without the platform. During the 5 training days, each mouse was practiced to find the platform within 60 sec. When a mouse found the platform, it was allowed to stand on the platform for 10 sec. Each mouse was given training trials twice a day, and the intertrial interval was 30 min. For each trial, the mouse was randomly placed in the water facing a visual cue on the wall, and the time the animal took to seek out the hidden platform (latency) was recorded. After each trial, the animal was returned to its original cage and allowed to dry under an infrared lamp. On the probe test day, the platform was taken away from the tank. Each mouse was permitted to seek out the platform in 60 sec. During the probe test, the movement of the mouse was recorded using the video camera-based EthoVision tracking system (Nodulus) to analyze the swimming velocity as well as the swimming time in the target area where the platform was previously set. During the 5 training days, 1 h before the first training trial, each day, mice were treated with AG-D (30, 100, or 300 mg/kg, p.o.) or donepezil (5 mg/kg, p.o.). The control group received vehicle solution (10% Tween 80 solution). Thirty minutes before the first training trial, each day, mice were also administered vehicle solution (0.9% saline solution, i.p.) or scopolamine (1 mg/kg, i.p.).
AChE activity assay
The AChE activity was assayed by a colorimetric analysis through an ex vivo study. 26 One hour postadministration of AG-D (100 or 300 mg/kg, p.o.) or donepezil (5 mg/kg, p.o.), mice were sacrificed and the whole brains, except the cerebellums, were isolated for the assay. 27 Brain tissues were homogenized in 10 × volume of homogenization buffer (0.1 M phosphate buffer, pH 8.0) by Teflon homogenizer (Eyela). Homogenates were centrifuged at 14,000 g at 4°C for 10 min. Each supernatant was used for the assay as the source of enzyme. It was mixed with 22 μL of Ellman's buffering reagent [10 mM 5,5′-dithiobis(2-nitrobenzoic acid) and 15 mM sodium bicarbonate], 144 μL of 0.1 M phosphate buffer (pH 8.0), and 1.1 μL of acetylthiocholine iodide solution, and reacted at room temperature for 10 min in 96 well. After 10 min of incubation, the reaction mixture in each well was stopped by adding 4.4 μL of the neostigmine solution. By using an ultraviolet (UV) spectrophotometer (OPTIZEN 2120UV; Mecasys Co., Ltd., Daejeon, Korea), the absorbance was measured at 412 nm.
Western blot analysis
One hour after a single administration of AG-D (100 or 300 mg/kg, p.o.) or donepezil (5 mg/kg, p.o.), a positive control, mice were sacrificed by cervical dislocation and brain tissues were isolated for Western blot analysis (WB). The control group received vehicle solution (10% Tween 80 solution). Isolated hippocampal tissues were homogenized in 200 μL of ice-cold 20 mM Tris-HCl buffer solution (pH 7.4), including a protease inhibitor cocktail (Roche, Seoul, Korea), 1 mM EGTA, 1 mM EDTA, 1 mM sodium orthovanadate, 1 mM PMSF, and 0.32 M sucrose per 50 mL of buffer. Homogenized lysates were centrifuged at 15,000 g for 20 min at 4°C twice. Then, the supernatant containing 15 μg protein was added into 10% gel of sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) with a reducing condition. All separated proteins were transferred to PVDF membranes by using the transfer buffer solution (25 mM Tris-HCl [pH 7.4] with 20% v/v methanol and 192 mM glycine) at 0.3 A for 2 h at 4°C. Thereafter, all the membranes were probed as described in the previous study. 28 The primary antibodies and its dilution factor for this study were as follows: cAMP response element-binding protein (CREB) or phosphorylated CREB (1:2000); extracellular signal-regulated kinase (ERK) or phosphorylated ERK (1:5000); protein kinase B (Akt) or phosphorylated Akt (1:3000); phosphatidylinositol 3-kinase (PI3K) or phosphorylated PI3K (1:2000); and glycogen synthase kinase 3β (GSK-3β) or phosphorylated GSK-3β (1:3000).
For the evaluation of temporal changes in BDNF expression, mice were sacrificed 3, 6, 9, and 12 h after a single administration of AG-D (100 or 300 mg/kg, p.o.) or 10% Tween 80 solution in the same volume. The 15 μg of crude proteins from homogenates was added into SDS-PAGE (12% gel) to determine the expression level of BDNF. Primary antibodies were used as the following dilution: BDNF (1:1000) and β-actin (1:3000). The other procedures were same as those described above.
Statistics
All data were expressed as the mean ± standard error of the mean and analyzed using Prism 5.0 software (GraphPad, La Jolla, CA). The data from each behavior test, and the AChE inhibition assay and the WB were analyzed by using one-way analysis of variance (ANOVA). For multiple comparisons, the Student-Newman-Keuls post hoc test was selected. The latencies of the MWM in the training trials and the temporal changes of BDNF expression levels were analyzed using two-way ANOVA, with time condition in the case of MWM or BDNF level set as the first variable and treatment conditions set as the second variable. For multiple comparisons, the Bonferroni post hoc test was selected. Significance of statistical differences was set to P < .05.
Results
AG-D ameliorated cholinergic blockade-induced memory impairment and enhanced cognitive function in the PAT
To evaluate whether the AG-D ameliorates the memory function on scopolamine-induced memory impairment in mice, we conducted PAT (Fig. 1A). The latencies during the acquisition trial showed no significant difference among all groups [F(5, 59) = 1.034, P > .05], but the latencies during retention trial were notable group effects across all groups [F(5, 57) = 21.68, P < .05]. The latency in the scopolamine (1 mg/kg, i.p.)-treated group was evidently shorter than that in the control group (P < .05). The AG-D (100 or 300 mg/kg, p.o.) significantly increased the latency, which was reduced by scopolamine (P < 0.05), as observed in the donepezil (5 mg/kg, p.o.) administration.

The effects of AG-D and marker compound, AG-D, 3,5-DCQA, on scopolamine-caused memory impairment
In addition, the enhancing effects of AG-D on memory function were also examined by using the PAT (Fig. 1B). During the acquisition trial, there were no significant group differences in the latencies among all groups [F(4, 49) = 0.3057], but notable group differences were observed in the retention trial [F(4, 48) = 11.99]. The latencies of AG-D-treated groups (100 or 300 mg/kg, p.o.) and the piracetam-treated group (200 mg/kg, i.p.), as a positive control, were significantly increased compared to the latency of the control group.
Furthermore, we also determined the effect of 3,5-DCQA, a marker compound of AG-D, on cholinergic blockade-induced memory impairment in mice (Fig. 1C). No significant differences in latencies between treatment groups were observed during the acquisition trial [F(5, 50) = 2.372]; however, significant group effects were observed in the retention trial [F(5, 50) = 27.21]. Scopolamine (1 mg/kg, i.p.)-treated mice showed lower latency time than those of control group (P < .05). The reduced latency caused by scopolamine was significantly reversed by 3,5-DCQA treatment (1 or 3 mg/kg, p.o.), as shown in the donepezil-treated group (5 mg/kg, p.o.). These results suggested that the administration of AG-D and 3,5-DCQA ameliorated the memory impairment induced by scopolamine and enhanced memory function in normal naive mice.
AG-D improved scopolamine-caused memory impairment in the Y-M
To know the effects of AG-D on working memory, we conducted the Y-M (Fig. 2A). Notable group differences were observed in the spontaneous alternation [F(5, 55) = 6.132, P < .05]. In the scopolamine (1 mg/kg, i.p.)-treated group, the spontaneous alternation was significantly less compared with the control group. In addition, the treatment with AG-D (100 or 300 mg/kg, p.o.) and donepezil (5 mg/kg, p.o.) significantly increased the reduced percentage of spontaneous alternation caused by scopolamine. However, the total number of arm entries among all groups was not significantly different [F(5, 58) = 2.283] (Fig. 2B). These results demonstrated that AG-D treatment enhanced the working memory ability in scopolamine-induced memory dysfunction in mice. The improvement in spontaneous alternation was not the result of the activation of locomotor behavior.
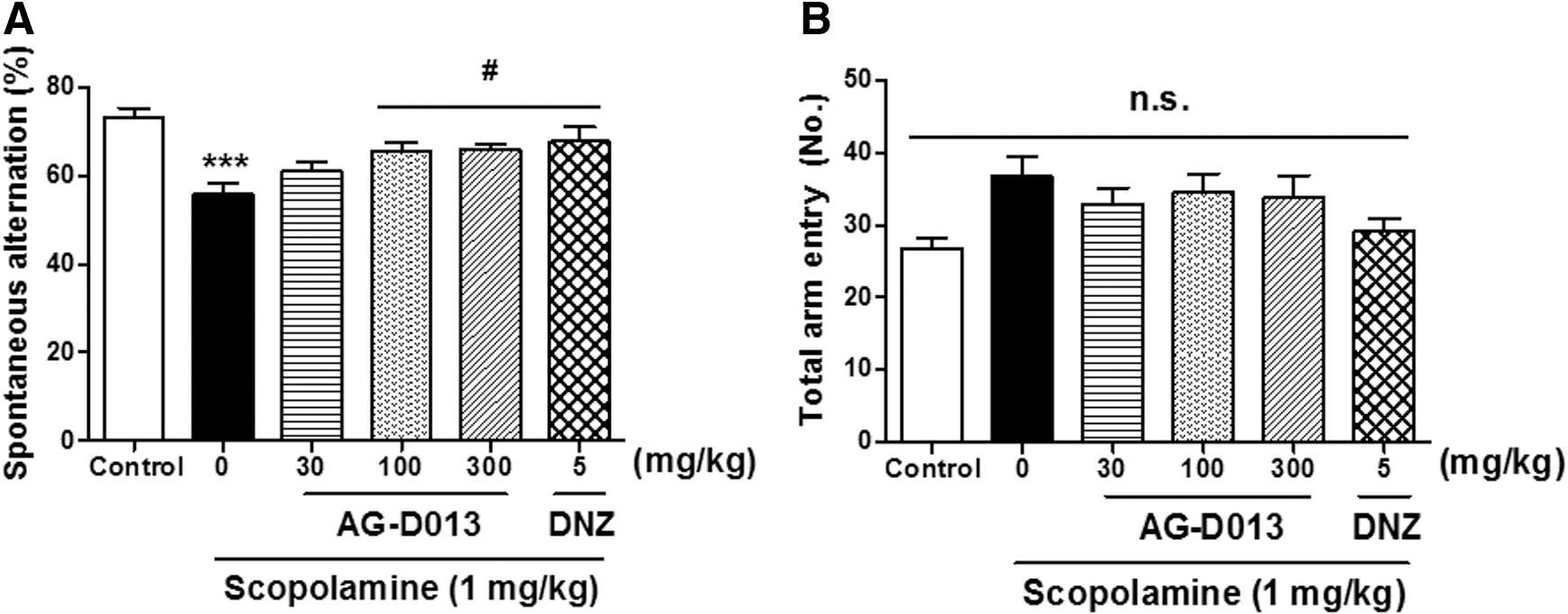
The effects of AG-D on scopolamine-caused memory impairment in the Y-M. AG-D (30, 100, or 300 mg/kg, p.o.), DNZ (5 mg/kg, p.o.), or equal volumes of vehicle solution (10% Tween 80 solution, p.o.) was used to treat the mice 1 h before the test. Memory impairment was caused by scopolamine (1 mg/kg, i.p.) 30 min before the Y-M. Spontaneous alternation
AG-D attenuated scopolamine-caused memory impairment in the MWM
To determine the effects of AG-D on spatial and long-term memory, we performed MWM. During the training trials, there were significant group effects of day [F(4, 262) = 77.87, P < .05] and treatment [F(5, 262) = 11.44] (Fig. 3A). The escape latencies of the scopolamine (1 mg/kg, i.p.)-treated group took longer than those of the control group during the training trials without first day and third day (P < .05). The escape latencies of both AG-D (30, 100, or 300 mg/kg, p.o.)- and donepezil (5 mg/kg, p.o.)-treated groups were significantly shorter than those of the group treated with scopolamine on the fourth and last day of training trials. There were notable group effects on swimming time in the target area in the probe trial [on day 6; F(5, 53) = 6.113, P < .05] (Fig. 3B). Compared to the control group, swimming time in the target area of the group was significantly shorter. In addition, the reduced swimming time caused by scopolamine in the target area tended to be increased by AG-D (30 or 100 mg/kg, p.o.), as observed in the group treated with donepezil (5 mg/kg, p.o.). However, the swimming velocities did not show any significant differences among all groups [F(5, 58) = 0.8206] (Fig. 3C). These results suggested that the administration of AG-D significantly alleviated the spatial and long-term memory disruption caused by cholinergic deficits.
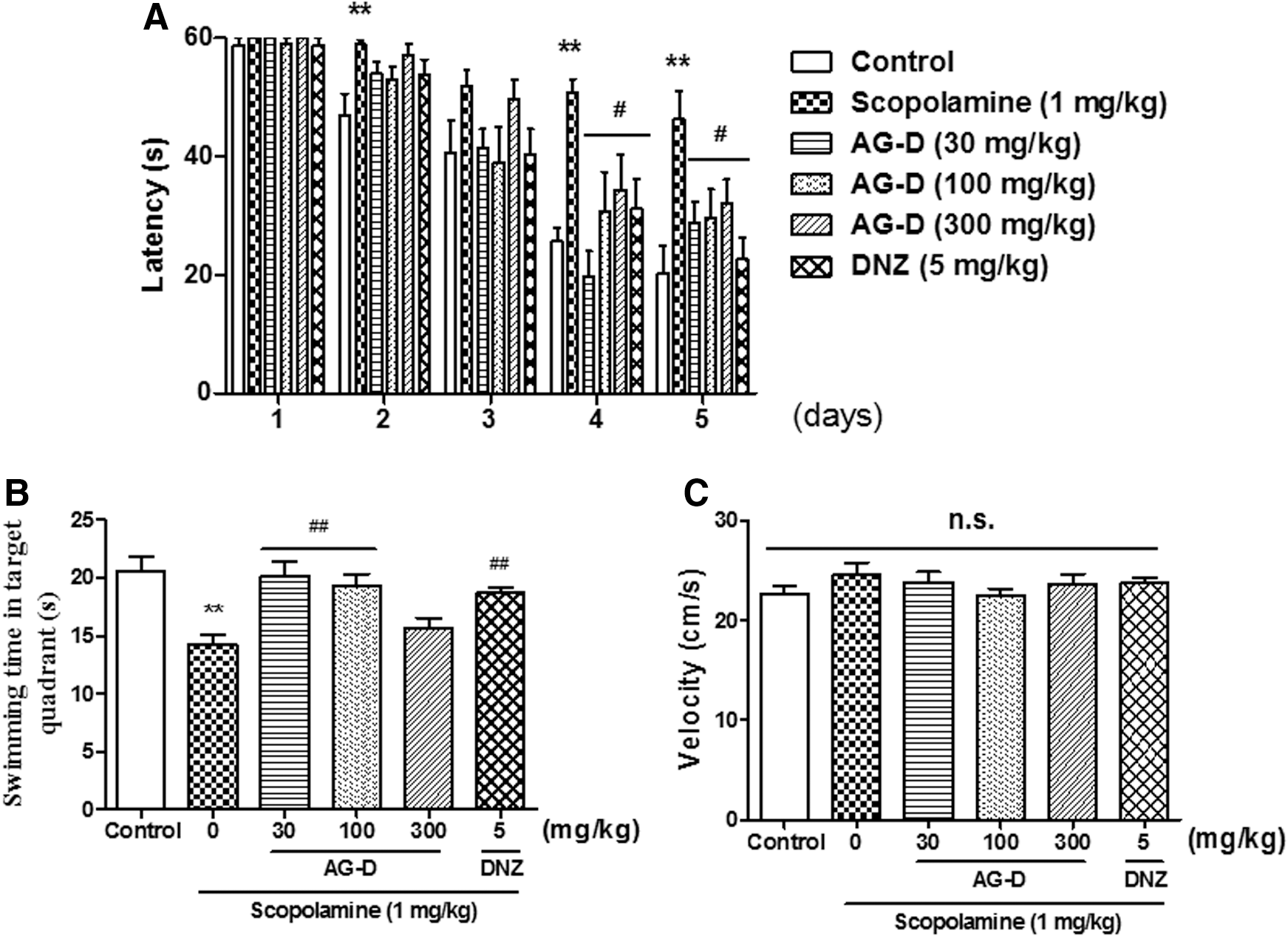
The effects of AG-D on scopolamine-caused memory impairment in the MWM. Escape latencies during the training trial days
AG-D inhibited AChE activity in an ex vivo assay
Because AG-D improved the cholinergic blockade-caused memory dysfunctions in the above behavioral tests, we investigated whether AG-D inhibits AChE activity. AChE activity was measured in an ex vivo condition. As shown in Figure 4, there were significant group effects on AChE inhibition [F(3, 19) = 7.463, P < .05]. AG-D (100 or 300 mg/kg) exhibited notable AChE inhibitory activity, as observed in the group treated with donepezil (P < .05). These results indicated that AG-D exerted inhibitory activity in AChE.

The inhibitory effect of AG-D on AChE activity in the mouse whole brain, except the cerebellum. AG-D (100 or 300 mg/kg, p.o.), DNZ (5 mg/kg, p.o.), or the equal volume of vehicle solution (10% Tween 80 solution, p.o.) was used to treat the mice 1 h before sacrifice. Whole brain, except cerebellum, was isolated and rapidly homogenized. Processed enzyme source was loaded onto a 96-well plate and incubated with reaction mixture for 10 min. The fluorescence was measured at 412 nm in a UV spectrophotometer. Data express the mean ± SEM (n = 5/group; *P < .05, compared with the control group treated with vehicle solution). AChE, acetylcholinesterase; UV, ultraviolet.
AG-D increased the phosphorylation of ERK, CREB, PI3K, Akt, and GSK-3β and the BDNF expression level in the hippocampus
To identify which signaling molecule(s) is related to the effects of AG-D on memory function, we conducted WB (Fig. 5). The levels of phosphorylated ERK or CREB were significantly increased in the hippocampal tissues after the administration of AG-D (100 or 300 mg/kg, p.o.) or donepezil (5 mg/kg, p.o.) [pERK, F(3, 17) = 6.995, P < .05 and pCREB, F(3, 19) = 5.448, P < .05]. In addition, the phosphorylation levels of PI3K and its downstream signaling molecules, including pAkt and pGSK-3β, were also significantly increased in the hippocampi caused by the administration of AG-D (100 or 300 mg/kg, p.o.) or donepezil [5 mg/kg, p.o.; pPI3K, F(3, 19) = 4.491, P < .05; pAkt, F(3, 18) = 8.717, P < .05; and pGSK-3β, F (3, 19) = 16.73, P < .05]. The levels of the above signaling molecules were significantly increased compared to those in the control group treated with the vehicle solution. In our previous studies, 28,29 ERK-CREB or PI3K-Akt-GSK-3β signaling pathway mediates the increase of BDNF expression levels at 9 and 12 h after each treatment. In that reason, we observed temporal profiles of BDNF expressions in the hippocampal tissues (Fig. 6). There were notable group effects of time [F(3, 47) = 6.426, P < .05] and treatment [F(2, 47) = 16.9, P < .05]. Compared to the control groups, the BDNF expression levels were evidently increased at 9 and 12 h postadministration of AG-D (100 or 300 mg/kg, p.o.). These results indicated that the administration of AG-D increased phosphorylation of ERK-CREB and PI3K-Akt-GSK-3β signaling molecules, and it might mediate the increase of BDNF expression at specific time points 9 and 12 h postadministration.
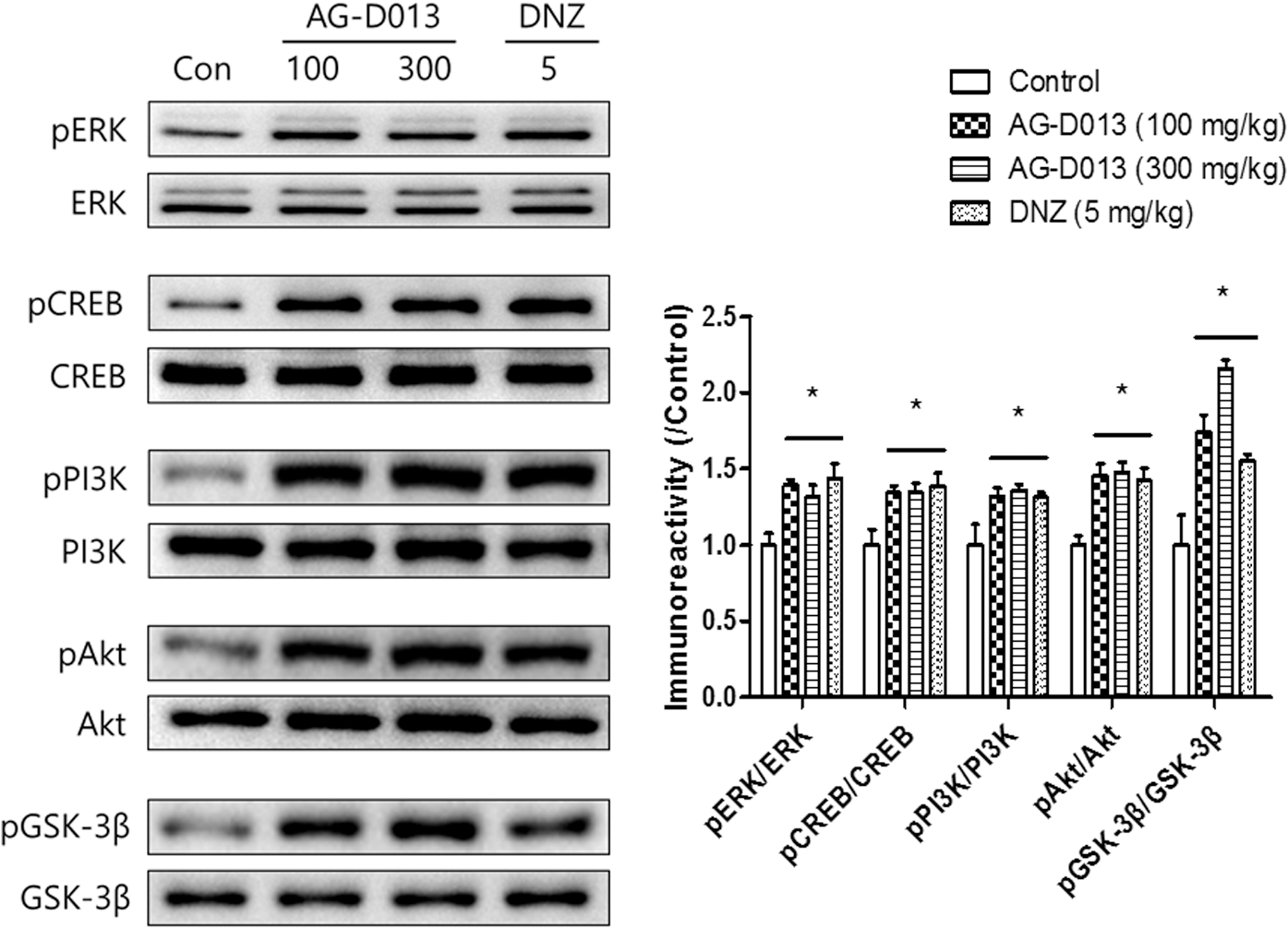
The effects of AG-D on the phosphorylation levels of ERK, CREB, PI3K, Akt, and GSK-3β in the hippocampi. One hour after the administration of AG-D (100 or 300 mg/kg, p.o.), DNZ (5 mg/kg, p.o.), or equal volume of the vehicle solution (Con, 10% Tween 80 solution, p.o.) mice were sacrificed. The immunoblotting of ERK, CREB, PI3K, Akt, GSK-3β, and their phosphorylation levels in the hippocampal tissues are represented in the left panel. The immunoreactivity levels of pERK/ERK, pCREB/CREB, pPI3K/PI3K, pAkt/Akt, and pGSK-3β/GSK-3β were standardized to those in the control group, and are presented in the right panel. Data are expressed as the mean ± SEM (n = 4–5/group; *P < .05, compared with the control group treated with vehicle solution). Akt, protein kinase B; CREB, cAMP response element-binding protein; ERK, extracellular signal-regulated kinase; GSK-3β, glycogen synthase kinase 3β; PI3K, phosphatidylinositol 3-kinase.
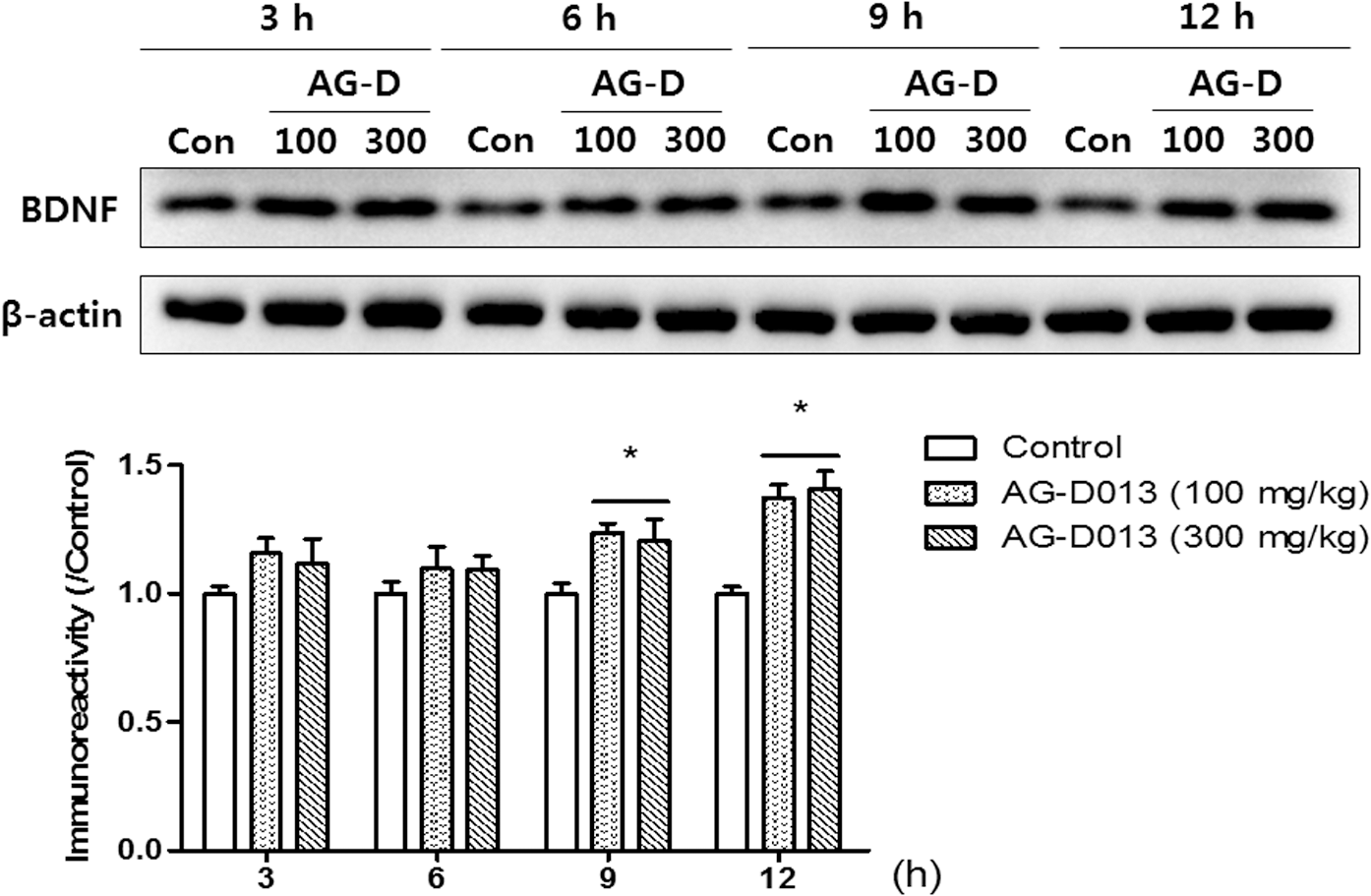
The temporal changes in the BDNF levels in the hippocampi after treatment with AG-D. Mice were sacrificed 3, 6, 9, and 12 h after the treatment of AG-D (100 or 300 mg/kg, p.o.) or equal volumes of vehicle (Con, 10% Tween 80 solution, p.o.). The immunoreactivity of BDNF and β-actin levels was measured in the hippocampal tissues. The immunoreactivity levels of BDNF/β-actin were standardized to the expression in the control groups, and are presented in the lower panel. Data are expressed as the mean ± SEM (n = 4–5/group; *P < .05, compared with the control group treated with vehicle solution). BDNF, brain-derived neurotrophic factor.
Discussion
In this study, we observed that AG-D ameliorated the cholinergic blockade-induced cognitive impairment in mice. The treatment with AG-D significantly reversed the memory impairment as assessed by PAT, Y-M, and MWM. AG-D also enhanced memory function in normal naive mice in the PAT. In addition, AG-D inhibited AChE activity in an ex vivo study. Moreover, we also found that the administration of AG-D increased the phosphorylation levels of ERK-CREB and PI3K-Akt-GSK-3β signaling pathways, and it mediated the increase of the expression level of BDNF in the hippocampi at specific time points.
First, we examined whether AG-D could reverse the working or long-term memory impairment induced by cholinergic blockade using several behavioral tests. The reduction in latency in the PAT and the reduction in the percentage of spontaneous alternation in the Y-M caused by scopolamine were increased by the treatment with AG-D (100 or 300 mg/kg), similar to that observed in the positive control group given donepezil. In addition, scopolamine decreased swimming time in the target area during the probe trial of the MWM and the administration of AG-D (30 or 100 mg/kg) significantly lengthened the swimming time in the target area. The total number of arm entries in the Y-M and the swimming velocity in the MWM were not different among all groups, indicating that locomotor activity was not affected by AG-D administration. All these results suggest that AG-D is effective for reversing working and long-term memory impairments associated with cholinergic dysfunction without any changes in behavioral activation.
Next, we investigated the mechanism of memory-ameliorating effect of AG-D. Cholinergic deficits are associated with AD, 30 and the synaptic levels of AChE in the brain are increased in AD patients. 31,32 These findings support the rationale for the prescription of AChE inhibitors for AD therapy. In this study, we conducted an AChE activity assay to examine the effects of AG-D on AChE activity. In the ex vivo study, AG-D significantly inhibited AChE activity, although the rate of AChE inhibition by AG-D was not higher than that by donepezil. These results suggest that AChE inhibition might be one of the mechanisms by which AG-D affects memory function.
Increased acetylcholine (ACh) levels will activate the ACh receptor, consequently activating the signaling pathway(s) associated with cognitive function. The ERK-CREB pathway is well acknowledged to play a key role in learning and memory processes 33 –35 and is activated by ACh receptor activation. 36 The activity of ERK is important for long-term potentiation (LTP) and memory consolidation. 37 In addition, ERK is needed for the maintenance of synaptic activities and the formation of several modi of memory. 27 CREB is a downstream signaling molecule of ERK and plays a crucial role in hippocampus-dependent memory. 38 –40 Therefore, we investigated whether AG-D changes the phosphorylation levels of ERK or CREB. In this study, the phosphorylation levels of ERK and CREB were evidently increased in the hippocampi by the administration of AG-D. In addition to CREB and ERK, BDNF is also important for memory formation, retention, and recall in the hippocampus. 41–42 Previously, we presented that upregulating BDNF expression levels at specific time windows, 9 and 12 h, is essential for memory consolidation and enhancement. 43 Similarly, the elevated expression levels of BDNF were observed 9–12 h after the treatment with AG-D. We found that AG-D inhibits AChE activity and activates the ERK-CREB-BDNF pathway. Until now, however, it has been unclear whether enhanced cholinergic neurotransmitter system activates the ERK-CREB-BDNF pathway. It has been reported that chronic donepezil treatment (15 months) has been shown to enhance serum BDNF levels in patients with mild cognitive impairment. 44 On the other hand, lesions in cholinergic innervations decreased BDNF messenger RNA (mRNA) expression in rat hippocampus. 45 Thus, our findings with others suggest that the cholinergic neurotransmitter system is closely associated with hippocampal BDNF expression. In addition, we also found that AG-D enhanced memory functioning in normal naive mice. These results mean that AG-D was able to ameliorate and enhance memory function, in part, by ERK-CREB-BDNF pathway activation and an increase in BDNF expression.
The PI3K-Akt pathway regulates synaptic plasticity and LTP. 46 PI3K is well known for its role in synaptic plasticity, consolidation of recognition memory, and LTP. 47 Akt is a downstream target of PI3K and is also involved in synapse formation and cell proliferation. 48,49 In addition, the activation of GSK-3β, a downstream of Akt, is also related to long-term memory 50 and enhances neurogenesis in the hippocampi, which results in cognitive enhancement. 51,52 In this study, the phosphorylation levels of PI3K, Akt, and GSK-3β were significantly increased in the hippocampi after the administration of AG-D. These results suggest that the ameliorating effects of AG-D on cognitive functions are due to, in part, its ability to activate the PI3K-Akt-GSK-3β signaling pathway. Furthermore, we investigated whether AG-D affects the phosphorylation levels of ERK-CREB and Akt-GSK-3β pathways under cholinergic blocking state. Although scopolamine is not a specific blocker of ERK and Akt, the administration of scopolamine significantly decreased the phosphorylation level of ERK and Akt. 53,54 Therefore, we conducted WB to determine the phosphorylation levels of ERK, CREB, Akt, and GSK-3β in the hippocampus after the administration of AG-D under scopolamine treatment (Supplementary Fig. S2). We found that AG-D significantly reversed the decreased phosphorylation level of ERK, CREB, Akt, and GSK-3β induced by scopolamine. These results also support the memory-ameliorating effects of AG-D.
A. glehni has antioxidant and anti-inflammatory activities and is used to treat insomnia and hypercholesterolemia in traditional and folk medicine. 11 As described in the introduction, sleep disturbance is a severe problem that is exacerbated by AD therapies. Therefore, AG-D would be a useful agent for the treatment of AD with less adverse effects. As of now, we do not know which compound(s) in AG-D exerts pharmacological effects on memory function. Caffeoylquinic acid derivatives, such as 3,5-DCQA, 5-O-caffeoylquinic acid, and 3-O-caffeoylquinic acid, are known bioactive compounds of A. glehni. 55 Such caffeoylquinic acid derivatives exhibit antioxidant and anti-inflammatory activities. 56,57 In addition, 3,5-DCQA and 3,4,5-tri-caffeoylquinic acid have been shown to inhibit cytotoxicity in human neuroblastoma SH-SY5Y cells caused by amyloid β-protein. 58 Recently, 3,5-DCQA was reported to decrease the activity of AChE. 59 Moreover, we also found that 3,5-DCQA ameliorated the scopolamine-caused memory impairment in mice as shown by the PAT. All of these observations suggest that 3,5-DCQA might play a role in the effects of AG-D, as an active compound. However, the amount of 3,5-DCQA in AG-D is not sufficient to be the only factor exerting these behavioral effects. Thus, AG-D likely exerts its ameliorating effects through the concerted effects of those caffeoylquinic acid derivatives.
Taken together, the above behavioral results suggest that AG-D is effective for reversing scopolamine-induced memory dysfunction, including long-term or working memory dysfunction, and enhances memory function. The results from the AChE activity assay and WB also suggest that the effects of AG-D on memory function are attributable to AChE inhibition and the activation of ERK-CREB-BDNF and PI3K-Akt-GSK-3β signaling pathways. This study suggests that AG-D is a promising agent for treating cognitive impairment induced by cholinergic dysfunction.
Footnotes
Acknowledgments
This research was supported by the High Value-added Food Technology Development Program (117077-03-1-HD040) and the Ministry for Food and Rural Affairs and Medical Research Center Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science and ICT (NRF-2017R1A5A2014768). We thank Chrislean Jun Botanas (College of Pharmacy, Shamyook University, Republic of Korea) for constructive discussions on the article and editorial input.
Author Disclosure Statement
The authors declare that there are no conflicts of interest.
Supplementary Material
Supplementary Data
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.