
Abstract
The fruits, leaves, and roots of Cudrania tricuspidata have been reported to contain large amounts of vitamin B, vitamin C, and flavonoids. They exhibit various physiological activities such as antitumor and anti-inflammatory effects. However, the hepatoprotective effects of C. tricuspidata extracts against oxidative stress-mediated liver injury have not yet been investigated. We thus examined whether C. tricuspidata leaf extracts (CTEs) protect against oxidative stress-mediated liver injury in vitro and in vivo and elucidated the underlying mechanism. The cytoprotective effects of CTE through the NF-E2-related factor 2 (Nrf2)/antioxidant response element (ARE) activation were presented and measured by biochemical analysis in HepG2 cells. To assess the protective effects of CTE in vivo, mice were administered with CTE (250 and 500 mg/kg; 5 days; p.o.) before a single dose of acetaminophen (APAP) (300 mg/kg; 24 h; i.p.). CTE increased ARE luciferase activity when compared with extracts of other parts of C. tricuspidata. CTE upregulated nuclear translocation of Nrf2 and its target gene expression. In addition, CTE inhibited the generation of reactive oxygen species (ROS) and cell death induced by arachidonic acid (AA) and iron (Fe) treatment in primary hepatocytes or HepG2 cells. The cytoprotective effects of CTE against oxidative stress might be due to kaempferol, the major flavonoid present in CTE. Kaempferol pretreatment blocked AA+Fe-induced ROS production and reversed glutathione depletion, which in turn led to decreased cell death. Furthermore, the protective effects of CTE against liver injury induced by excess APAP in mice or primary hepatocytes were observed. CTE could be a promising therapeutic candidate against oxidative stress-induced liver injury.
Introduction
Oxidative stress has been shown to play a pivotal role in pathologic changes observed in various liver diseases. 1,2 The redox state regulates inflammatory, metabolic, fibrogenic, and proliferative liver disorders. The liver is the principal organ in charge of xenobiotic metabolism and is potentially more exposed or susceptible to oxidative stress-associated molecules. 3 At present, no specific therapeutics is available for oxidative stress-related acute liver injury.
Cudrania tricuspidata is a deciduous thorny tree belonging to the family Moraceae, and it is mainly distributed in Asian countries, including Korea, China, and Japan. It has been extensively used as an important traditional oriental medicine or health supplement for the treatment of various ailments such as eczema, mumps, tuberculosis, contusions, insomnia, and acute arthritis. 4 In addition, several pharmacological activities of C. tricuspidata extract, including anticancer, 5 antioxidative, 6 antihyperglycemic, 7 and anti-inflammatory 8 effects, have been reported. However, the hepatoprotective effects of C. tricuspidata extract on oxidative stress-mediated acute liver damage remain unknown. Chemical studies have shown that the major constituents of C. tricuspidata leaf extract (CTE) are phenolic compounds, including xanthones and flavonoids such as kaempferol, 9 possessing important biological activities. 10
Previously, we established and reported an in vitro model using arachidonic acid (AA) and iron (Fe) treatment in HepG2 cells to induce cell death and mitochondrial dysfunction. 11 This model has been used to examine the effects and molecular mechanisms of candidate drugs for the treatment of oxidative stress-mediated acute hepatic disease. 12,13 In the present study, we adopted this model to investigate the effects of C. tricuspidata extracts and its major constituent kaempferol. In addition, we used an in vivo experimental animal model of acetaminophen (APAP)-induced acute liver injury in mice to confirm the hepatoprotective activities of C. tricuspidata extracts. APAP-associated liver toxicity is characterized by extensive oxidative stress and is often accompanied by lipid peroxidation. 14 The Fe chelation with deferoxamine protects against APAP-induced liver damage due to the chelation of Fe, which can catalyze reactive oxygen species (ROS) generation in hepatocytes. 15
Therefore, in the present study, we aimed to evaluate the biological activities of C. tricuspidata extracts on oxidative stress-associated liver injury. The activation of NF-E2-related factor 2 (Nrf2), a fundamental regulator of antioxidant and detoxification abilities, and the upregulation of its target genes by CTE were shown to protect in response to AA+Fe-induced oxidative injury and to exert cytoprotective effects in hepatocytes. Biochemical and histopathological analyses showed the protective effects of CTE against APAP-mediated oxidative stress, inflammatory responses, and liver injury. Moreover, we determined the pharmacological efficacy of kaempferol, the major functional constituent of CTE. These results show that CTE protects against liver damage via the alleviation of oxidative stress and inflammatory responses, suggesting that CTE could be a promising therapeutic candidate for the prevention and treatment of acute liver disease.
Materials and Methods
Materials
Antibodies against Nrf2 (sc-722) and NADPH quinone oxidoreductase 1 (NQO1; sc-16464) were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Phospho-Nrf2 (NB100-80012) antibody was provided from Novus Biologicals (Littleton, CO, USA). Lamin A/C (no. 2032) antibody was obtained from Cell Signaling (Danvers, MA, USA). Glutamate cysteine ligase (GCL; ab41463) antibody was obtained from Abcam (Cambridge, MA, USA). Horseradish peroxidase-conjugated goat anti-rabbit (G21234) and anti-mouse (G21040) antibodies were purchased from Invitrogen (Carlsbad, CA, USA). Heme oxygenase (HO)-1 (ADI-SPA-896) antibody was provided by Enzo Life Sciences (Plymouth Meeting, PA, USA). 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), tert-butylhydroquinone (t-BHQ), 2′-7′-dichlorodihydrofluorescein diacetate (DCFH-DA), dimethyl sulfoxide (DMSO), kaempferol, APAP, and β-actin antibody were purchased from Sigma Chemicals (St. Louis, MO, USA). PD98059, SB203580, and SP600125 were obtained from Calbiochem (Billerica, MA, USA).
Preparation of CTE
The fruits, leaves, branches, and roots used for C. tricuspidata extract were provided by Lifetree Biotech (Suwon, Gyeonggi-do, Korea).
Cell culture
HepG2 cells were provided by American Type Culture Collection (ATCC, Manassas, VA, USA). The cells were seeded into 6-well plates at a density of 1 × 106 cells/well, and the wells with 70–80% confluency were used. The cells were incubated in Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS; Atlas, Fort Collins, USA), 50 units/mL penicillin, and 50 mg/mL streptomycin at 37°C in a humidified incubator with 95% air/5% CO2 atmosphere. Cells are treated with CTE and maintained for the indicated time at 37°C. The cells were then rinsed twice with ice-cold 1 × phosphate-buffered saline (PBS).
Primary hepatocyte isolation
Primary hepatocytes were isolated from ICR mice and cultured as reported previously. 16 Briefly, mice were anesthetized with Zoletil (Virbac, France), and portal vein cannulation was performed. The liver was perfused in situ with Ca2+-free Hank's balanced salt solution (HBSS) at 37°C for 5 min, followed by perfusion of 0.05% collagenase with HBSS and Ca2+ at 10 mL/min for 20 min. After that, the livers were minced using sterile scissors and reconstituted in sterilized PBS. The cell suspension was filtered via a cell strainer and centrifuged at 50 g for 5 min to pellet only hepatocytes. Viability of isolated hepatocytes evaluated by trypan blue dye is usually 80–90%. Isolated hepatocytes were seeded in collagen-coated plates with DMEM containing 50 units/mL of penicillin/streptomycin mix with 10% FBS.
AA and Fe-nitrilotriacetate treatment
Cells were plated for 24 h and then incubated in minimum essential medium (MEM) for 12 h. Cells were treated with 10 μM AA and C. tricuspidata extracts or kaempferol for 15 h, followed by exposure to 5 μM Fe-nitrilotriacetate (Fe) for the indicated time. After that, cells stained with 0.2 mg/mL MTT or 10 μM DCFH-DA were subjected to measure cytotoxicity or ROS production, respectively.
MTT assay
To determine cytotoxicity, HepG2 cells were seeded into 24-well plates at a density of 1 × 105 cells/well and exposed to 100, 200, and 400 μM C. tricuspidata extracts for 24 h. For the experiment with primary hepatocyte, we treated cells with APAP (10 mM) or AA+Fe for the indicated time period after pretreatment with CTE for 30 min. After incubation, the cells were stained with MTT (final concentration of 0.2 mg/mL) for 4 h as previously described. 17 The media were then removed and formazan crystals were dissolved in 300 μL DMSO. The absorbance was measured at 550 nm with a microplate reader (Spectra MAX; Molecular Devices, Sunnyvale, CA, USA). Relative cell viability was expressed as the percentages of relative to the vehicle-treated control [i.e., viability (% control) = 100 × (absorbance of CTE-treated sample)/(absorbance of vehicle-treated control sample)].
Luciferase assay
To provide rapid and reproducible results for antioxidant response element (ARE) luciferase activity, we constructed a HepG2 cell line stably transfected with NQO1-ARE firefly luciferase gene, which contains three tandem repeats of ARE and the 5′-upstream region of NQO1 as previously described. 17 The luciferase activity was measured with the Luciferase Assay Reagent II (Promega, Madison, WI, USA).
Measurement of ROS production
Cells were stained with 10 μM DCFH-DA for 30 min at 37°C. Cellular ROS generation was measured by dichlorofluorescein staining. Fluorescence intensity was determined with a fluorescence microplate reader (Gemini; Molecular Devices) at excitation/emission wavelengths of 485/530 nm or by flow cytometry (Beckman).
Immunoblot analysis
Whole-cell lysates or subcellular fractions were prepared as previously described. 18 Sodium dodecyl sulfate/polyacrylamide gel electrophoresis and Western blotting were carried out according to previously published procedures. 18
Determination of glutathione content
The cellular glutathione (GSH) levels were measured using a commercial GSH assay kit (BIOXYTECH GSH-400; Oxis International) as previously reported. 19 The cells were cultured in 6-well plates and subjected to chemical exposure. The cells were scraped and then lysed in 5% metaphosphoric acid to precipitate the cellular proteins. After centrifugation (10,000 g, 15 min), the supernatants were used for estimation of GSH concentration. The absorbance was determined at a wavelength of 400 nm with a microplate reader (Spectra MAX; Molecular Devices).
Animals
The protocols for animal research were approved by the Institutional Animal Care and Use Committee of Chosun University (CIACUC2017-A0029). Male ICR mice (7 weeks old) were purchased from Orient Bio (Seongnam, Korea) and acclimatized to the environment for 1 week. The mice were housed at an ambient temperature of 20°C ± 2°C with 12 h light/dark cycles in relative humidity of 50% ± 5%, with food (Purina, Korea) and water available ad libitum. The mice were administered with CTE (250 and 500 mg/kg; 5 days; p.o.) before being given a single dose of APAP (300 mg/kg; 24 h; i.p.) or vehicle.
Histopathology
Liver samples were harvested and fixed in 10% neutral buffered formalin, embedded in paraffin, sectioned into 3–4 μm slices, and stained with hematoxylin and eosin (H&E) for general histopathological image analysis as previously published. 20 For immunohistochemical analysis, an experienced histopathologist was blinded to the group distribution. Immunohistochemical staining was carried out with 4-hydroxynonenal (4-HNE; ab46545; diluted 1:100; Abcam, Cambridge, United Kingdom) or nitrotyrosine (NT; 06-284; diluted 1:200; Millipore, Temecula, CA, USA) antibodies as previously published. 20 Briefly, the tissue sections were deparaffinized in xylene, hydrated in graded ethanols, and incubated with 10 mM citrate buffer (pH 6.0). The tissue was placed in hydrogen peroxide to inactivate endogenous peroxidase and blocked with normal horse serum (Vector Labs, Inc., Burlingame, CA, USA). Inactivated tissue sections were incubated with the primary antibodies overnight at 4°C and then stained by the method using avidin/biotin interaction (Vector Labs, Inc.). If the cells were occupied by >10% of immunoreactives, the relative density of each antiserum compared with intact cells, they were regarded as positive. The number of 4-HNE- and NT-positive cells per mm2 was calculated through a digital image analysis at 400 × magnification, and the sections were processed by light microscopy (Nikon, Tokyo, Japan).
Blood chemistry
Serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels were measured using commercially available kits supplied by Asan Pharmaceutical (Seoul, Korea), as previously published. 20
Statistical analysis
One-way analysis of variance was used to assess the statistical significance of differences between groups. The Newman–Keuls test was applied to determine the significance of differences between the several groups. All data are expressed as mean ± standard error.
Results
Nrf2 activation by C. tricuspidata extracts
First, we examined the cell viability and cytotoxic effect of extracts of C. tricuspidata fruits, leaves, branches, and roots using MTT assay in human hepatocyte-derived HepG2 cells. No significant toxicities were detected in cells treated with extracts from the different parts of C. tricuspidata at concentrations up to 400 μg/mL (Fig. 1A). Nrf2, a redox-sensitive transcription factor, regulates cellular responses to oxidative stress by binding to AREs. 21 We compared the relative ARE luciferase activity to determine the antioxidant efficacy of extracts of C. tricuspidata fruits, leaves, branches, and roots in the HepG2 cell line stably expressing NQO1-ARE. CTE showed significant dose-dependent increases in ARE luciferase activity compared with extracts from other parts (Fig. 1B). Tert-Butylhydroquinone (t-BHQ), a well-known Nrf2 activator, was adopted as the positive control.
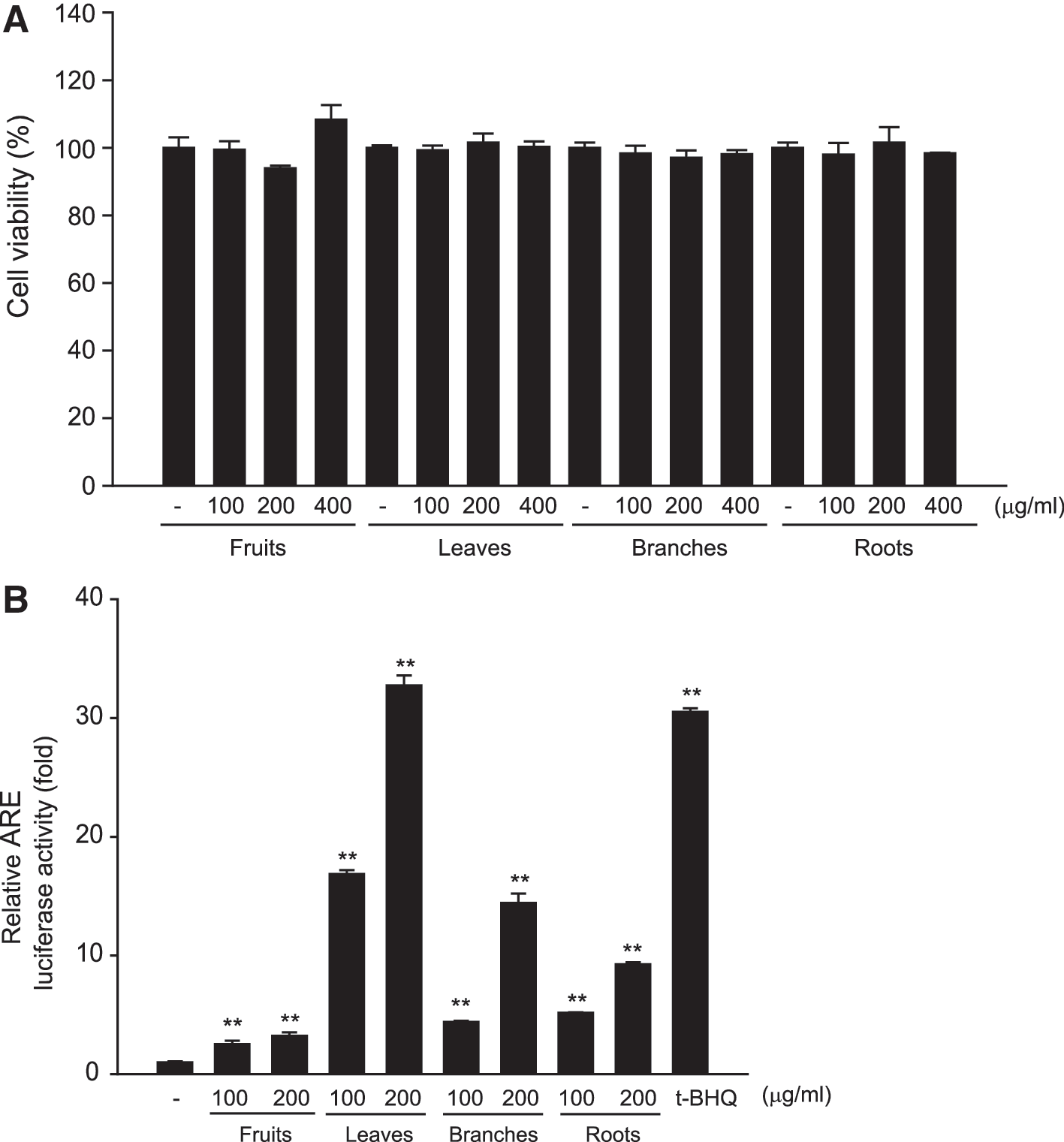
Effects of Cudrania tricuspidata extracts on cytotoxicity and ARE luciferase activity.
Nrf2 target gene induction by CTE
The Nrf2-ARE signaling pathway is associated with Nrf2 phosphorylation and subsequent Nrf2 release from Keap1, thereby allowing Nrf2 migration into the nucleus and binding to ARE in the promoter region of antioxidant defense genes. 22 To investigate the effect of CTE on Nrf2 activation, cells were incubated with CTE for different durations and nuclear accumulation of Nrf2 was observed. Treatment with CTE augmented the level of nuclear Nrf2 at 6 h. However, treatment with CTE did not change the total level of Nrf2 (Fig. 2A). Next, we investigated whether nuclear Nrf2 accumulation leads to its target gene expression, including GCL, NQO1, and HO-1. GCL is an enzyme involved in pathway of GSH biosynthesis, and its expression is primarily modulated by the Nrf2-ARE pathway. In addition, NQO1 and HO-1 are well-studied Nrf2-regulated genes and protect against oxidative stress. As expected, the protein levels of GCL, NQO1, and HO-1 were upregulated by CTE treatment (Fig. 2B). Nrf2 phosphorylation was also increased by CTE treatment (Fig. 2C). To examine the signaling pathway associated with the regulatory mechanism of Nrf2 phosphorylation by CTE, we treated the cells with mitogen-activated protein kinase (MAPK) inhibitors (Fig. 2D). HepG2 cells treated with MAPK inhibitors showed that the extracellular signal-regulated kinases (ERK) inhibitor PD98059 specifically reduced HO-1 expression before exposure to CTE, indicating that ERK-mediated phosphorylation is required for Nrf2 activation and upregulation of antioxidant defense gene expression by CTE.

Effect of CTE on Nrf2 activation.
Inhibition of ROS production by CTE
ROS serve as cellular signaling molecules in the maintenance of normal physiological functions. However, excessive production of ROS results in oxidative stress and increases the sensitivity of cells to ROS-mediated cytotoxicity. Therefore, we examined the effect of CTE on AA+Fe-induced ROS generation. HepG2 cells treated with CTE reduced AA+Fe-induced ROS generation, which was comparable with that obtained with Trolox, a water-soluble analog of α-tocopherol used as positive control (Fig. 3A). As rhodamine 123 (Rh123) is used as an indicator of mitochondrial damage, we used Rh123 as a fluorescent probe for changes in mitochondrial membrane potential (MMP) of cells. The subpopulation of Rh123-negative cells was increased by AA+Fe treatment. However, CTE inhibited the MMP transition induced by AA+Fe treatment (Fig. 3B). Next, we studied the effect of CTE pretreatment on the cytotoxic effects induced by AA+Fe. CTE pretreatment markedly protected against AA+Fe-mediated cell death in HepG2 cells (Fig. 3C). The protective effect of CTE on AA+Fe-induced ROS generation and cell death was confirmed in mouse primary hepatocytes (Fig. 3D, E). These data indicate that the attenuation of ROS production and increase in cell viability are due to the antioxidant effects of CTE.
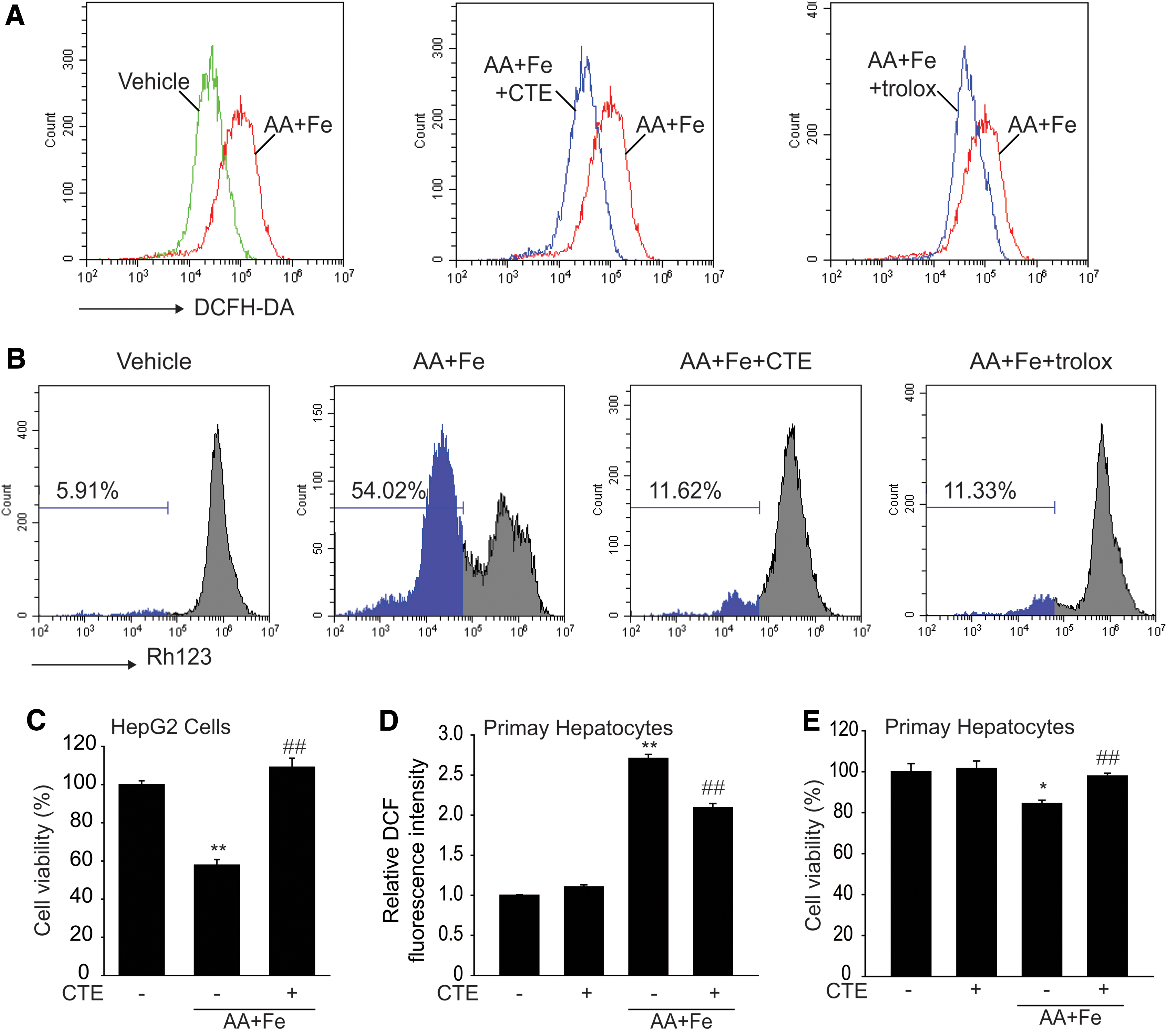
Inhibition of AA+Fe-induced oxidative stress by CTE.
Cytoprotective effect of kaempferol against oxidative stress
Kaempferol is the major functional constituent of CTE. 9 Flavonoids are shown to possess anti-inflammatory, anticancer, and antioxidant activities. First, we examined the cytotoxicity of kaempferol. No significant toxicities were observed up to 200 μM in HepG2 cells (Fig. 4A). ARE luciferase activity was significantly increased by kaempferol in the HepG2 cell line stably expressing NQO1-ARE luciferase plasmid (Fig. 4B). Next, we investigated the cytoprotective efficacy of kaempferol on AA+Fe-induced cell injury. Treatment with AA+Fe induced cell death, which was significantly inhibited when the cells were pretreated with kaempferol (Fig. 4C). Kaempferol also reduced ROS production induced by AA+Fe (Fig. 4D). In addition, kaempferol restored GSH depletion induced by AA+Fe treatment (Fig. 4E). Our results demonstrate that CTE and its primary flavonoids inhibit oxidative stress-mediated hepatocyte injury.
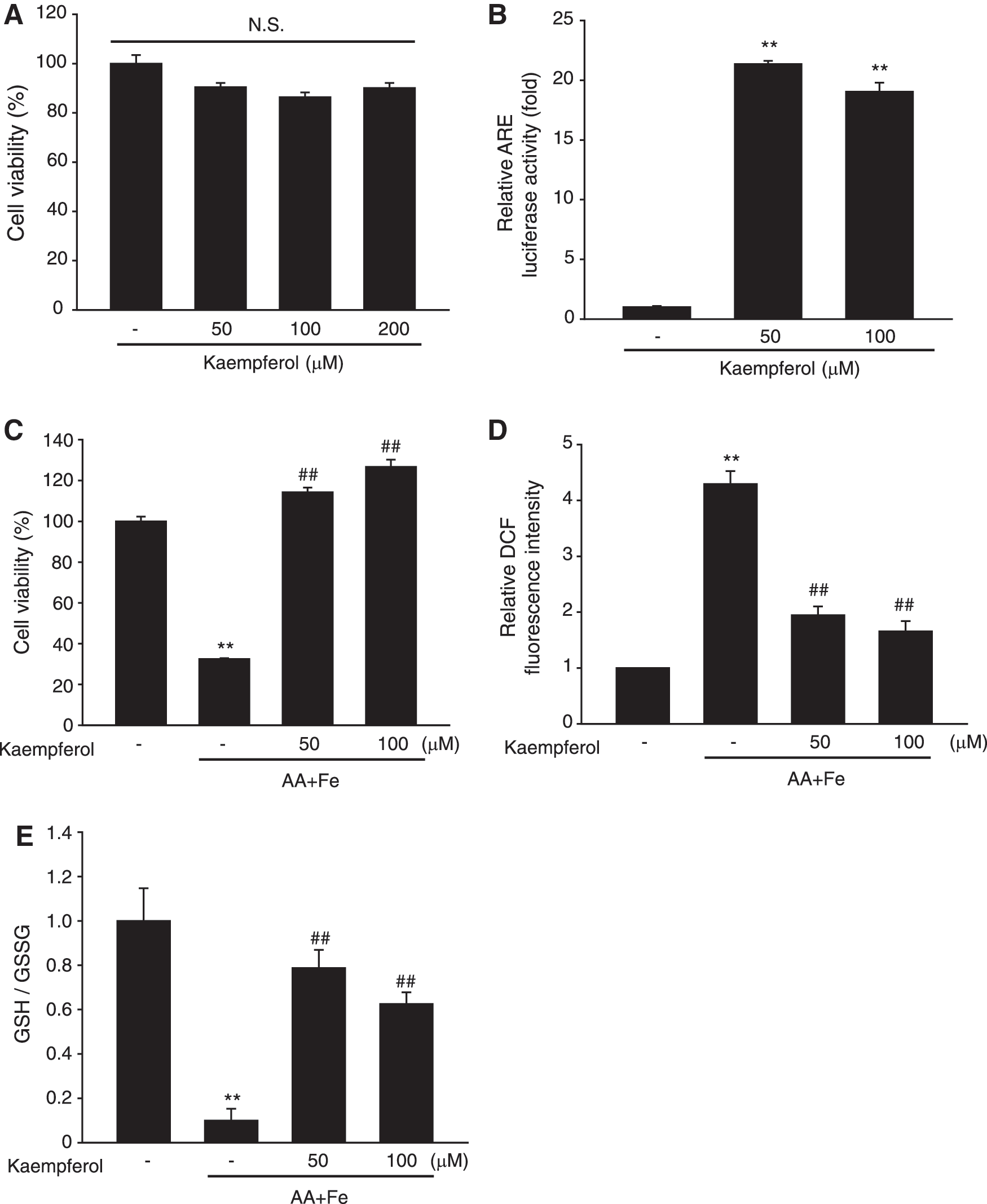
Cytoprotective effects of kaempferol on AA+Fe-induced oxidative damage.
Inhibition of APAP-induced acute liver injury by CTE in vivo
To assess the protective effects of CTE in vivo, we used an animal model of acute liver injury induced by APAP treatment. In mice, the commonly used serum markers for liver damage such as ALT and AST were significantly increased at 24 h after APAP treatment (300 mg/kg); however, these levels were significantly reduced by CTE treatment at a concentration of 250 or 500 mg/kg (Fig. 5A). Similar to the results of blood biochemistry, H&E staining showed a significant protection against the development of degenerative changes, cellular inflammatory infiltration, and abnormal hepatocytes in the liver tissue (Fig. 5B and Table 1). These data indicated that CTE protected against acute liver injury induced by APAP.
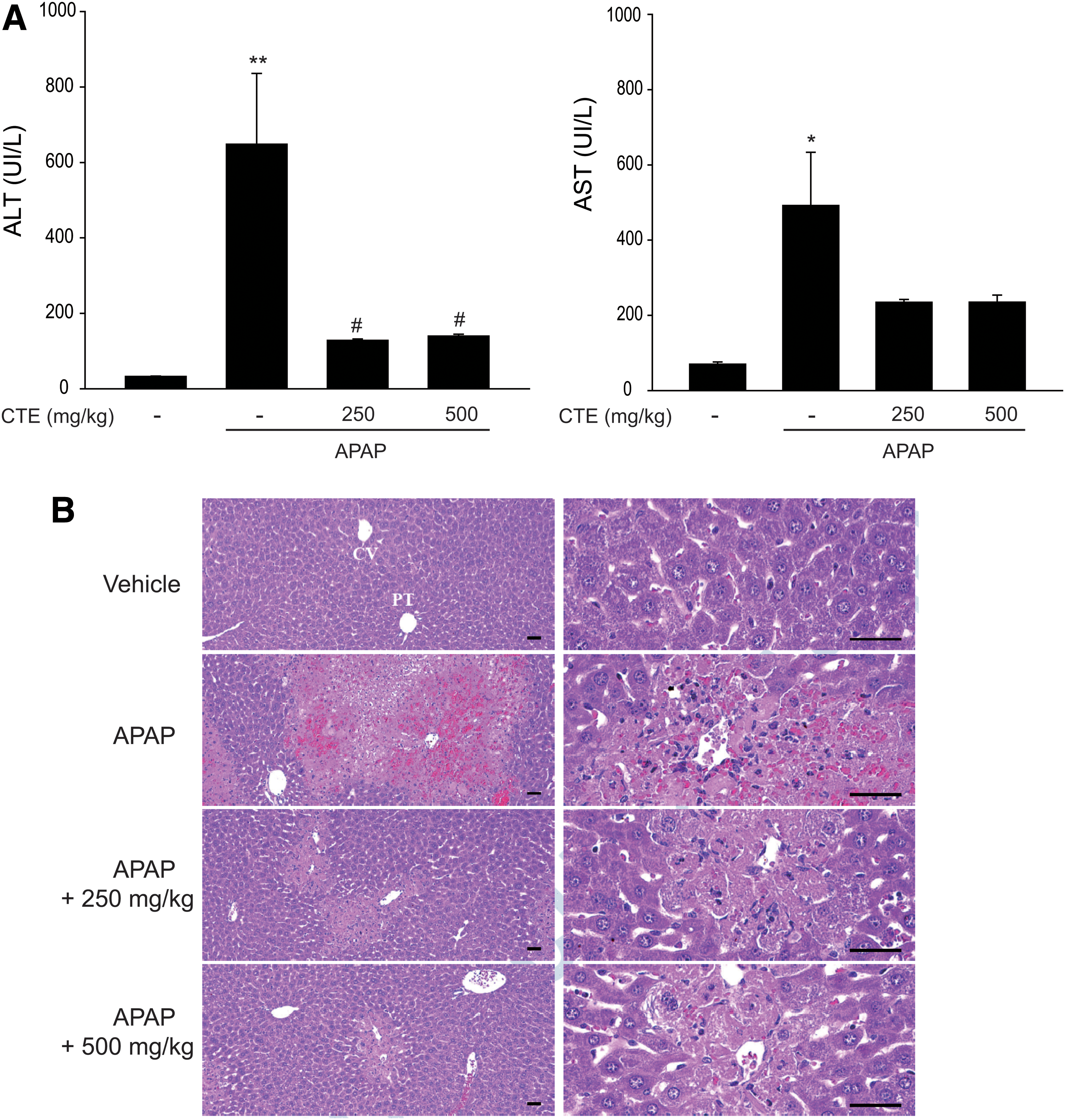
Inhibition of APAP-induced liver injury by CTE.
General Histomorphometric Analysis of Hepatic Tissues
Values are expressed as mean ± SD of six hepatic histological fields.
P < .01 and * P < .05 compared with vehicle-treated mice.
P < .01 and # P < .05 compared with APAP-treated mice.
APAP, acetaminophen; CTE, Cudrania tricuspidata leaf extract; SD, standard deviation.
Inhibition of APAP-mediated oxidative stress and apoptosis by CTE
NT is well known as an indicator of cell damage, inflammation, and nitric oxide production. Tyrosine nitration is mediated by reactive nitrogen species, including peroxynitrite formed by generation of nitric oxide and superoxide through xanthine oxidase. 23 The number of NT-immunoreactive cells was increased in mice treated with a single administration of APAP; however, treatment with CTE significantly inhibited this increase (Fig. 6A left). 4-HNE, a major α, β-unsaturated aldehyde of n-6 polyunsaturated fatty acid oxidation, has been used as a biomarker of lipid peroxidation, which is associated with various pathologies such as neurodegenerative diseases, metabolic disorders, and cancers. 24 4-HNE-immunoreactive cells were found in APAP-administered mice, and these cells were markedly inhibited by CTE treatment (Fig. 6A right). Consistently, APAP-administered mice exhibited a marked increase in ROS levels compared with vehicle-treated mice, while CTE treatment reduced ROS levels in the liver tissues (Fig. 6B).
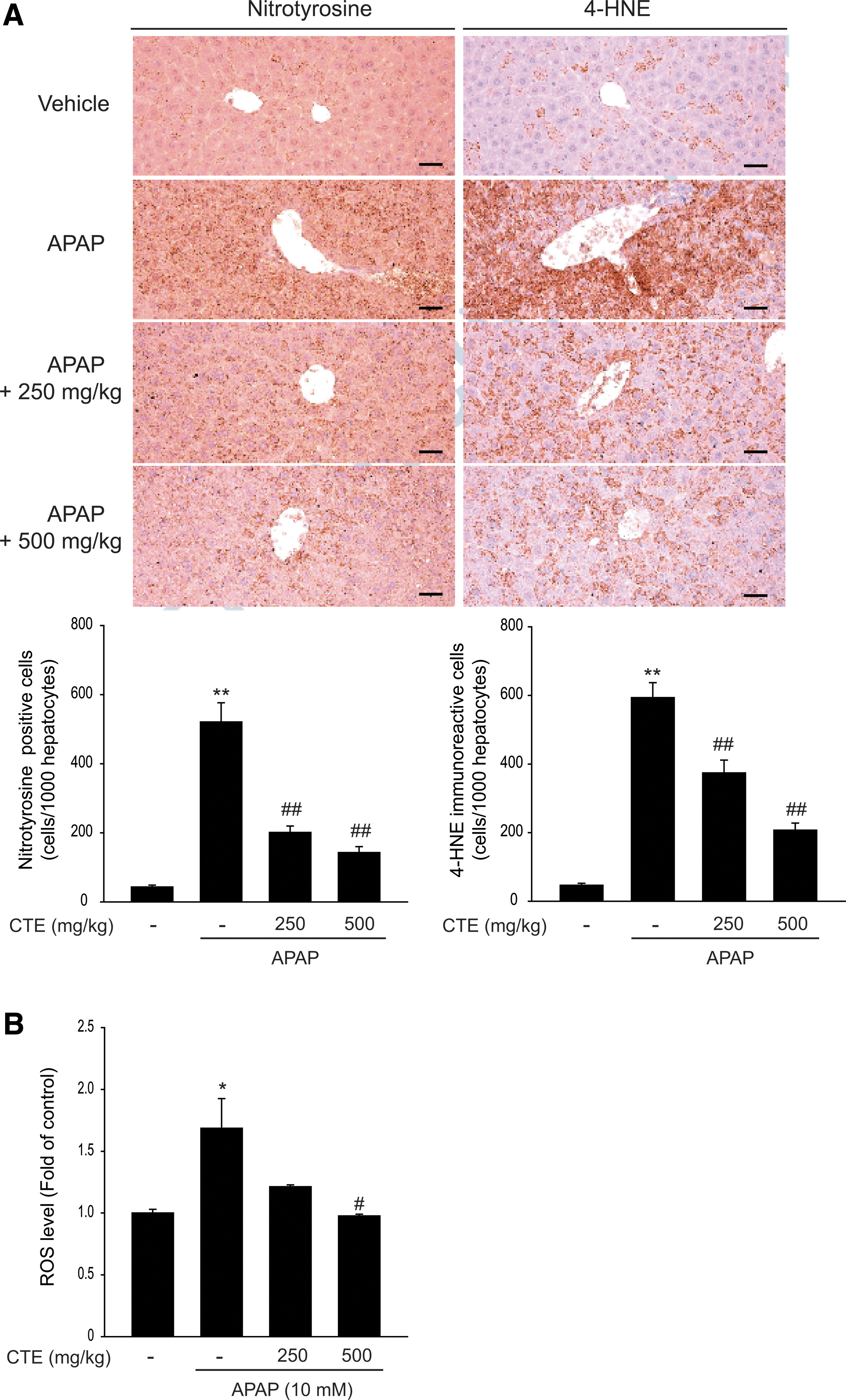
Inhibition of APAP-induced oxidative stress by CTE.
Furthermore, CTE markedly reduced the increase in caspase-3 cleavage and poly (adenosine diphosphate-ribose) polymerase cleavage, which are considered to be hallmarks of apoptosis, induced by a single challenge of APAP in hepatic tissues (Fig. 7A). Next, we isolated primary hepatocytes from mice to investigate the cytoprotective effect of CTE against APAP. Consistently, treatment with CTE significantly reversed APAP-induced cell death in primary hepatocytes (Fig. 7B). These results indicate that CTE exhibited a beneficial effect on APAP-mediated acute liver injury through its antioxidant capacity and inhibition of apoptosis (Fig. 7C).

Inhibition of APAP-induced apoptosis by CTE.
Discussion
It is well established that the progression of acute liver injury is associated with increased oxidative stress and decreased antioxidant capacity. 25,26 Hence, the role of Nrf2 activation in the pathophysiology of liver diseases has been comprehensively studied using in vitro and in vivo experimental animal models. 21 Comparison of Nrf2 activity in cells treated with extracts of C. tricuspidata fruits, leaves, branches, and roots by luciferase assay showed that the leaves had a stronger Nrf2 activity than other parts, which was similar to that observed in t-BHQ-treated cells. These findings were in agreement with a previous study in which the total polyphenol levels and antioxidant effects were highest in leaves of C. tricuspidata. 27 CTE increased phosphorylation and nuclear translocation of Nrf2, resulting in the induction of its target gene expression, namely GCL, NQO1, and HO-1. HO-1 upregulation by CTE was specifically antagonized by the ERK inhibitor, but not by other MAPK inhibitors. These results suggested a role for ERK in Nrf2 activation and its target gene induction by CTE. A relationship between ERK activation and Nrf2-mediated antioxidant gene expression has been reported. 28
Furthermore, CTE reduced ROS production, mitochondrial permeability transition, and cell death induced by AA+Fe. ROS activate cytosolic phospholipase A2, and consequently release AA from cell membranes. 29 AA accumulation induces oxidative stress and inflammation, resulting in apoptosis through mitochondrial impairment in cells. 30 In the presence of AA, Fe synergistically induces cell death by decreasing the MMP. 31 Thus, the combination of AA and Fe is considered a toxicant, inducing cell damage via oxidative stress. It is adopted to assess the efficacy of candidate drugs and probe the underlying molecular and cellular mechanisms that protect cells from oxidative stress-induced apoptosis. 12,32 Our present study showed that CTE inhibited AA+Fe-induced ROS generation and MMP transition. Moreover, CTE improved cell viability, which was decreased by AA+Fe in HepG2 cells or primary hepatocytes. These observations indicated that the inhibition of ROS production and MMP transition by CTE could protect cells against oxidative stress-mediated cell death.
Although APAP is the most commonly used nonprescription analgesic, an excessively large amount of APAP may cause severe liver damage and failure. Oxidative stress is a major factor that mediates hepatic tissue damage by APAP. 14 Thus, overdose of APAP in experimental animals is wildly used to investigate the hepatoprotective potential of candidate drugs. The major underlying mechanisms of APAP-induced hepatotoxicity in rodent models have been confirmed in patients with APAP-induced liver injury 33 and in human hepatocytes. 33,34 APAP-mediated hepatotoxicity is caused by the formation of the reactive toxic metabolite N-acetyl-p-benzoquinone imine by cytochrome P450 enzymes, resulting in hepatic GSH depletion and thereby inducing oxidative stress. APAP-induced hepatotoxicity is aggravated in Nrf2-deficinet mice, 35,36 and Nrf2 activation is regarded a promising therapeutic strategy for APAP-mediated liver injury. CTE inhibited APAP-induced ALT and AST levels, lipid peroxidation, ROS production, and apoptosis markers. These results support the notion that the hepatoprotective effect of CTE might be due to Nrf2 activation.
It was reported previously that the flavonoid kaempferol is the major constituent of CTE, and the constituents of CTE exhibited antioxidant efficacy. 27 Kaempferol in water extracts from C. tricuspidata leaves improved inflammation and insulin sensitivity in endoplasmic reticulum stress-induced hepatocytes. 37 However, the effect of kaempferol on AA+Fe-induced hepatocyte damage has not yet been investigated. We succeeded in demonstrating that kaempferol inhibited ROS formation, MMP transition, and cell death in AA+Fe-treated hepatocytes. These results indicate that the hepatoprotective effect of CTE is mediated, at least in part, by the antioxidant effect of kaempferol. Kaempferol has been shown to exert hepatoprotective effects against drug- or alcohol-mediated liver injury. 38
Collectively, the present study clearly shows that CTE and its major constituent kaempferol inhibited mitochondrial dysfunction and cell death caused by AA+Fe. Furthermore, CTE inhibited APAP-induced ROS formation and liver injury. Our present data strongly suggest that CTE may be a potential therapeutic candidate to successfully prevent or treat oxidative stress-induced liver injury.
Footnotes
Acknowledgments
This work was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through the Agri-Bio Industry Technology Development Program, funded by the Ministry of Agriculture, Food and Rural Affairs (MAFRA; 316007-5).
Authors' Contributions
S.H.K., D.S.O., S.S.C., and C.-S.N.: conceived and designed the experiments; S.S.C., J.H.Y., K.H.S., G.U.J., J.H.E., and J.B.K.: performed the in vivo and in vitro evaluation; S.K.K. and I.J.C.: performed histological analysis. S.H.K., S.M.S., E.Y.P., and J.S.P.: analyzed the data and wrote the article.
Author Disclosure Statement
No competing financial interests exist.