
Abstract
Senile osteoporosis increases the risk of skeletal fractures with age. Cheonggukjang (CGJ), a traditional Korean dry fermented soybean product, has numerous therapeutic effects; however, its effects on bone mineral density (BMD) and bone metabolism in senile osteoporosis are unclear. In this study, we treated the senescence-accelerated mouse prone 6 (SAMP6) model of senile osteoporosis with CGJ to determine its potential for ameliorating and preventing osteoporosis progression. High-performance liquid chromatography analysis for isoflavone profiles revealed that short-term fermentation significantly increased the isoflavone aglycone content in soybeans. Thereafter, we fed 6-week-old SAMP6 mice with experimental diets containing 5% or 10% CGJ for 15 weeks. Microcomputed tomography revealed that CGJ supplementation effectively increased the BMD and relative bone length. In vitro, CGJ increased the osteopontin reactivity and upregulated the expression of Alp, Col1a1, Fak, Bmp2/4, Smad1/5/8, and Runx2 in osteoblasts, and decreased Cathepsin K reactivity and downregulated Rankl and Nfatc1 expression in osteoclasts. In addition, CGJ increased the osteoprotegerin/Rankl ratio. Collectively, these results demonstrate that CGJ can ameliorate the detrimental effects of senile osteoporosis by improving osteogenesis and decreasing osteoclast activity.
Introduction
Osteoporosis is a skeletal disease characterized by a reduction in bone mass and an increased risk of bone fracture. 1,2 Bone remodeling occurs upon achieving a balance between bone resorption and formation, which is required for maintaining skeletal integrity and strength. 3 However, the bone remodeling process gradually decelerates with age, and the resulting imbalance in the bone turnover results in bone loss. 4,5 This increase in bone turnover results in deterioration of the bone microarchitecture and a reduction in bone mineral density (BMD), leading to a condition known as senile osteoporosis, which is characterized by an increased susceptibility for low-trauma fracture in elderly individuals. 6,7
Food and Drug Administration (FDA)-approved osteoporosis treatments for men include antiresorptives such as bisphosphonates (alendronate, risedronate, and zoledronic acid: inhibitor of farnesyl pyrophosphate synthase) and denosumab (inhibitor of receptor activator of nuclear factor kappa-B ligand [RANKL]). 9 Although these drugs are widely used because of their efficacy, treatment of osteoporosis through hormone therapy with antirestorative agents is controversial owing to its potential adverse effects. 10 Thus, extensive research effort has been devoted to establishing alternative treatment strategies for osteoporosis; however, most of these studies primarily focused on postmenopausal osteoporosis, and there are few reports on potential treatment and/or preventive options for senile osteoporosis.
Cheonggukjang (CGJ) is one of the several types of Korean traditional fermented soybean food products, which is a dried fermented soybean product characterized by a short fermentation time of only 3 days. 11,12 Traditionally, CGJ is made from boiled soybean (BS) and airborne microbes or microflora present in rice straw. 13,14 Hence, CGJ contains various enzymes, microorganisms, and bioactive compounds that are absent in unfermented soybeans. 15
During CGJ fermentation, many proteins are degraded into readily absorbable small peptides and amino acids, and the total flavonol and phenolic acid contents are increased. 11,16,17 The most significant changes from the soybean fermentation, including CGJ, is that the isoflavone glycosides are converted to aglycones by hydrolysis. Shiguemoto et al. and Yoo et al. have reported that consumption of fermented soybean products was shown to effectively ameliorate bone-related disorders owing to the increased isoflavone aglycone content. 18,19 In a previous study, we have revealed that isoflavone metabolites in CGJ have beneficial effects on bone formation. 12,20 Furthermore, Setchell and Lydeking-Olsen have reported that soybean isoflavone inhibited bone loss in both female and male osteoporotic animal models. 21 Several studies have reported the antiosteoporotic effect of CGJ using the ovariectomized (OVX) model. 22 –24 However, the specific effect of CGJ on bone metabolism in male senile osteoporosis is unknown.
OVX animals are most commonly used to study bone metabolism in osteoporosis. This model is followed by accelerated bone loss through estrogen deficiency, paralleling the situation in humans after menopause. This model, however, allows only the analysis of female animals and, thus cannot be used for the study of gender differences in fracture healing of osteoporotic bone.
Senescence-accelerated mouse prone 6 (SAMP6) mice is the first rodent model of senile osteoporosis, which develops spontaneous fractures at older ages. SAMP6 displays a bone phenotype consistent with that of human senile osteoporosis, including reduced trabecular bone volume, decreased bone strength, and reduced BMD. Contrary to OVX model, SAMP6 mice model may be used for gender studies, because osteoporosis occurs in both male and female animals. 25,26 Therefore, in this study, we prepared CGJ by the short-term dry fermentation of soybean and evaluated its effects on male senile osteoporosis in SAMP6 mice. As a control, we used the senescence-accelerated mouse resistance 1 (SAMR1) mouse model, which is resistant to osteoporosis with aging. 6,27,28 In addition, the effect of CGJ on the expression of genes related to bone formation and bone resorption were evaluated in an effort to reveal the underlying mechanism. Together, this work may provide a theoretical basis for the development of CGJ as a potential antiosteoporotic food.
Materials and Methods
Preparation of the CGJ sample
CGJ samples were obtained from Hamssine Traditional Soybean Food (Jeonju-si, Jeollabuk-do, Korea). In brief, soybeans (Glycine max (L.) Merr.) were washed and then soaked overnight in water. The next day, the soybeans were steamed for 1 h at 110°C, cooled to 40°C, and used as the BS sample. The BS was fermented with naturally occurring airborne microorganisms such as Bacillus subtilis for 3 days, and ultimately used as the CGJ samples. The prepared samples were lyophilized and stored in a −80°C freezer until use.
Isoflavone analysis
The isoflavones were detected and quantified by high-performance liquid chromatography (HPLC) coupled with an ultraviolet-visible detector (Jasco, Tokyo, Japan) and YMC-Pack ODS column (250 × 4.6 mm, 5 μm; Yamamura Chemical, Kyoto, Japan) using daidzin, glycitin, genistin, malonyl-daidzin, malonyl-glycitin, malonyl-genistin, acetyl-daidzin, acetyl-glycitin, acetyl-genistin, daidzein, glycitein, and genistein standards (Nacalai, San Diego, CA, USA). BS and CGJ were extracted using 0.2% 0.1 N hydrochloric acid in HPLC grade acetonitrile (JT Baker, Inc., Phillipsburg, NJ, USA) and mixed for 2 h at room temperature. The isoflavones were detected at 245 nm. Mobile phases comprised 0.1% glacial acetic acid (JT Baker, Inc.) in water (A) and 0.1% glacial acetic acid in acetonitrile (B) as a gradient system for 65 min. The flow rate was 1.0 mL/min. After injection of 20 μL of the sample, solvent B was increased from 15% to 35% over 50 min, and then held at 35% for 10 min. HPLC grade water was obtained through a Sartorius water purification system (0.22 μm pore size; Sartorius Stedim Biotech GmbH, Goettingen, Germany). All other chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Animal experimentation
Six-week-old male SAMP6 (n = 33) and SAMR1 (n = 11) mice were purchased from Japan SLC, Inc. (Hamamatsu, Japan). The mice were housed individually in plastic cages in a temperature-controlled room (23 ± 1°C), with a 12:12-h light/dark cycle, and ad libitum access to food and water. After 1 week of acclimatization with a normal chow diet (2018S, 6.2% fat, 44.2% carbohydrate, 18.6% protein, 18.2% fiber; Harlan Teklad, Madison, WI, USA), the mice were divided into four experimental groups (n = 11 in each group) as follows: group 1, SAMR1; group 2, SAMP6 control; group 3, SAMP6 fed 5% CGJ (w/w); and group 4, SAMP6 fed 10% CGJ (w/w). The experimental diets were based on the AIN-93G (Harlan Teklad). Body weight was recorded once a week and food intake was recorded every 2–3 days. For urine collection, the mice were housed for 16 h in metabolic cages (one mouse per cage) for 14 weeks. At the end of the experimental period, blood samples were collected from the orbital venous congestion under anesthesia through intraperitoneally administered tribromoethanol (avertin; 250 mg/kg body weight; Sigma Aldrich). The tibia and femur were dissected and the relative bone length was measured by dividing the length of bones by the body weight. The Korea Food Research Institute Animal Care and Use Committee (KFRI-M-17007) approved the experimental protocol.
Microcomputed tomography analysis
Isolated left femurs of the mice were dissected, and the distal metaphysis was scanned through a microcomputed tomography (μ-CT) scanning process using the SkyScan 1076 μ-CT scanner (Bruker μ-CT, Seeshaupt, Germany) at ∼18 μm/μm3 resolution. Parameters of the trabecular bone such as trabecular BMD (mg/cm3), trabecular volume (mm3), bone surface per total volume (BS/TV, mm2/mm3), bone volume per total tissue volume (BV/TV, %), and bone surface per bone volume (BS/BV, mm2/mm3) were also analyzed and quantified.
Serum and urine biochemical analysis
Serum calcium (Ca) was measured with a Ca kit (Cayman, MI, USA). Bone-specific alkaline phosphatase (Bone ALP), calcitonin, tartrate-resistant acid phosphatase type 5 (TRACP5b), and deoxypyridinoline (DPD) were determined using commercial kits (Kamiya, Seattle, WA, USA). C-terminal telopeptide of type I collagen (CTX-1) levels were measured with a test kit (My BioSource, San Diego, CA, USA). RANKL was determined using a commercially available test kit (Abcam, Cambridge, MA, USA). Serum magnesium (Mg), phosphorus (P), aspartate aminotransferase (AST), alanine aminotransferase (ALT), and urine Ca and creatinine (Cr) levels were determined at Green Cross Reference Lab (Youngin-Si, Gyeonggi-do, Korea).
Immunohistochemistry
Isolated right femurs were fixed in 10% phosphate-buffered formalin for 3 days at 4°C. The femur was then decalcified in a 0.25 M ethylenediaminetetraacetic acid (solution for 2 weeks and then embedded in paraffin. A 5 μm thick femur section was prepared and incubated with the polyclonal rabbit-immunoglobulin G (IgG) anti-osteopontin (OPN) antibody (dilution, 1:200; Abcam no. ab8448; Abcam, Cambridge, United Kingdom) and polyclonal rabbit-IgG anti-cathepsin K (CTK) antibody (dilution, 1:200; Abcam no. ab19027). The slides were then incubated with hematoxylin as a counterstain. The stained areas were viewed using a light microscope with a magnifying power of × 200.
Quantitative real-time polymerase chain reaction analysis
Total RNA from the tibia was isolated using TRIzol® reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) and a NucleoSpin® RNA II kit (Macherey-Nagel GmbH & Co., Duren, Germany) according to the manufacturer instructions. To quantify mRNA expression levels, cDNA was prepared as described previously, 29 and quantitative real-time polymerase chain reaction (qRT-PCR) was performed using the SYBR® Green PCR Master Mix (Applied Biosystems, Grand Island, NY, USA). The expression level of each mRNA was normalized to that of β-actin. The primer sequences used for qPCR are given in Table 1.
Primer Sequences for Quantitative Real-Time Polymerase Chain Reaction
Alp, alkaline phosphatase; Bmp2, bone morphogenetic protein 2; Bmp4, bone morphogenetic protein 4; Col1a1, collagen type 1 alpha 1; Ctk, cathepsin K; Fak, focal adhesion kinase; Igf-1, insulin-like growth factor-1; Nfatc1, nuclear factor of activated T-cells 1; Opg, osteoprotegerin; Opn, osteopontin; Rankl, receptor activator of nuclear factor kappa-B ligand; Runx2, runt-related transcription factor 2; Smad1, Smad protein 1; Smad5, Smad protein 5; Smad8, Smad protein 8.
Statistical analysis
Data were expressed as mean ± standard error of the mean. Statistical analysis was performed using one-way analysis of variance (ANOVA) to compare the quantitative data among groups with GraphPad Prism 7 (San Diego, CA, USA). The Bonferroni post hoc test was used when the ANOVA was significant (P < .05).
Results
Quantification of isoflavones
The isoflavone profile and contents of the BS and CGJ samples were determined by HPLC, as described previously. 30 To compare isoflavone profiles in BS and CGJ, we identified 12 isoflavone peaks (Fig. 1A), only seven of which were used for comparison: daidzin, glycitin, genistin, daidzin, glycitein, acetyl-genistin, and genistein (Fig. 1B, C). After short-term fermentation, the aglycone-type isoflavones such as daidzein, glycitein, and genistein were elevated; thereafter, the glycoside-, malonyl-, and acetyl-type isoflavones were decreased (Table 2). These data confirmed that CGJ is a very important source of isoflavone aglycones such as daidzein and genistein despite the short fermentation period of soybean.

Typical chromatogram of
Isoflavone Contents of BS and CGJ a
Each value represents the mean ± standard deviation of three independent replications.
BS, boiled soybean; CGJ, Cheonggukjang; ND, not detected.
Effect of CGJ on body weight, food intake, and bone length
Changes in the body weights of the mice during the 15-week experimental period are given in Figure 2A. The initial average body weights were significantly lower in the SAMR1 mice compared with those of the SAMP6 mice (P < .01), and this difference between the mouse strains remained consistent throughout the experimental period. However, there was no significant change in the body weight or food intake of the SAMP6 mice after 15 weeks of CGJ treatment (Fig. 2B, C). Although the relative tibia and femur lengths of the SAMP6 control group were significantly lower than those of the SAMR1 mice, CGJ treatment significantly increased these lengths in a dose-dependent manner. Treatment with 10% CGJ ameliorated the reduction of relative bone lengths of the SAMP6 model, reaching the same level as that of the SAMR1 mice (Fig. 2D, E). Exogenous insulin-like growth factor 1 (IGF-1) can promote longitudinal femur growth and bone formation through direct effects on osteoblasts. 31,32 As given in Figure 2F, the Igf-1 mRNA expression was significantly upregulated upon CGJ supplementation compared with the values reported for the SAMP6 control. These data indicate that CGJ improved bone length and upregulated Igf-1 expression in SAMP6, without influencing body weight.

Effects of CGJ supplement on body weight and bone length.
Effect of CGJ on bone morphometric parameters
To ensure that the CGJ treatment ameliorated the trabecular microarchitecture, we assessed trabecular parameters through μ-CT (Fig. 3A). As given Figure 3A, two- and three-dimensional μ-CT scans showed that the SAMP6 control exhibited significant loss of the trabecular bones compared with the SAMR1 mice. However, the CGJ treatment dramatically attenuated this trabecular bone loss in SAMP6 mice. As expected, the SAMP6 control mice also exhibited significant loss of trabecular BMD and bone volume, BS/TV, and BS/BV compared with the SAMR1 mice, which was dramatically attenuated after CGJ supplementation, as evident from the microarchitectural parameters, compared with the SAMP6 control (Fig. 3B, C). These data suggest that treatment with CGJ could improve senile osteoporosis through recovery of the trabecular microarchitecture.

Effects of CGJ supplement on senile osteoporosis.
Effect of CGJ on serum and urine bone turnover markers
Serum levels of Ca, b-ALP, OCN, CTX-1, RANKL, and TRACP5b were all significantly increased upon CGJ treatment compared with those in the SAMP6 control (Table 3). However, there was no significant difference in the serum P, ALP, ALT, and calcitonin levels among the groups. The urine Ca/Cr ratio was significantly higher in the group administered 10% CGJ than in the SAMP6 control. The level of urinary DPD significantly decreased upon CGJ treatment compared with that of the SAMP6 control. These data suggest that CGJ recovered bone loss in SAMP6 by effectively increasing bone formation and reducing bone resorption concurrently.
Effect of Cheonggukjang Supplementation on Serum and Urine Bone Turnover Parameters a
Results of each parameter are presented as mean ± standard error of the mean of 11 mice per group.
P < .05, ** P < .01, *** P < .001, and NS compared with the SAMP6 control.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; Bone ALP, bone alkaline phosphatase; Ca, calcium; Ca/Cr, calcium/creatinine; CTX-1, C-terminal telopeptide of type I collagen; DPD, deoxypyridinoline; Mg, magnesium; NS, not significant; OCN, osteocalcin; P, phosphorous; RANKL, receptor activator of nuclear factor kappa-B ligand; SAMP6, senescence-accelerated mouse prone 6; SAMR1, senescence-accelerated mouse-resistance 1; TRACP5b, tartrate-resistant acid phosphatase 5b.
Effect of CGJ on bone formation
To confirm the effects of CGJ on promoting osteogenesis, we performed immunohistochemical staining and detected the mRNA expression levels of osteogenic genes in the femur. As given in Figure 4A, the OPN reactivity in SAMP6 control mice was the lowest among all groups, which was effectively increased by CGJ treatment. Opn mRNA expression was significantly upregulated in the 10% CGJ treatment compared with that of the SAMP6 control group (Fig. 4B). Bone formation-associated genes such as Alp, Col1a1, and Fak were also significantly upregulated in the 10% CGJ treatment compared with those in the SAMP6 control (Fig. 4C). In addition, the expression levels of BMP signaling genes such as Bmp2 and 4, Smad1, 5, and 8, and Runx2 were significantly increased in the 10% CGJ treatment group compared with the SAMP6 control (Fig. 4D).
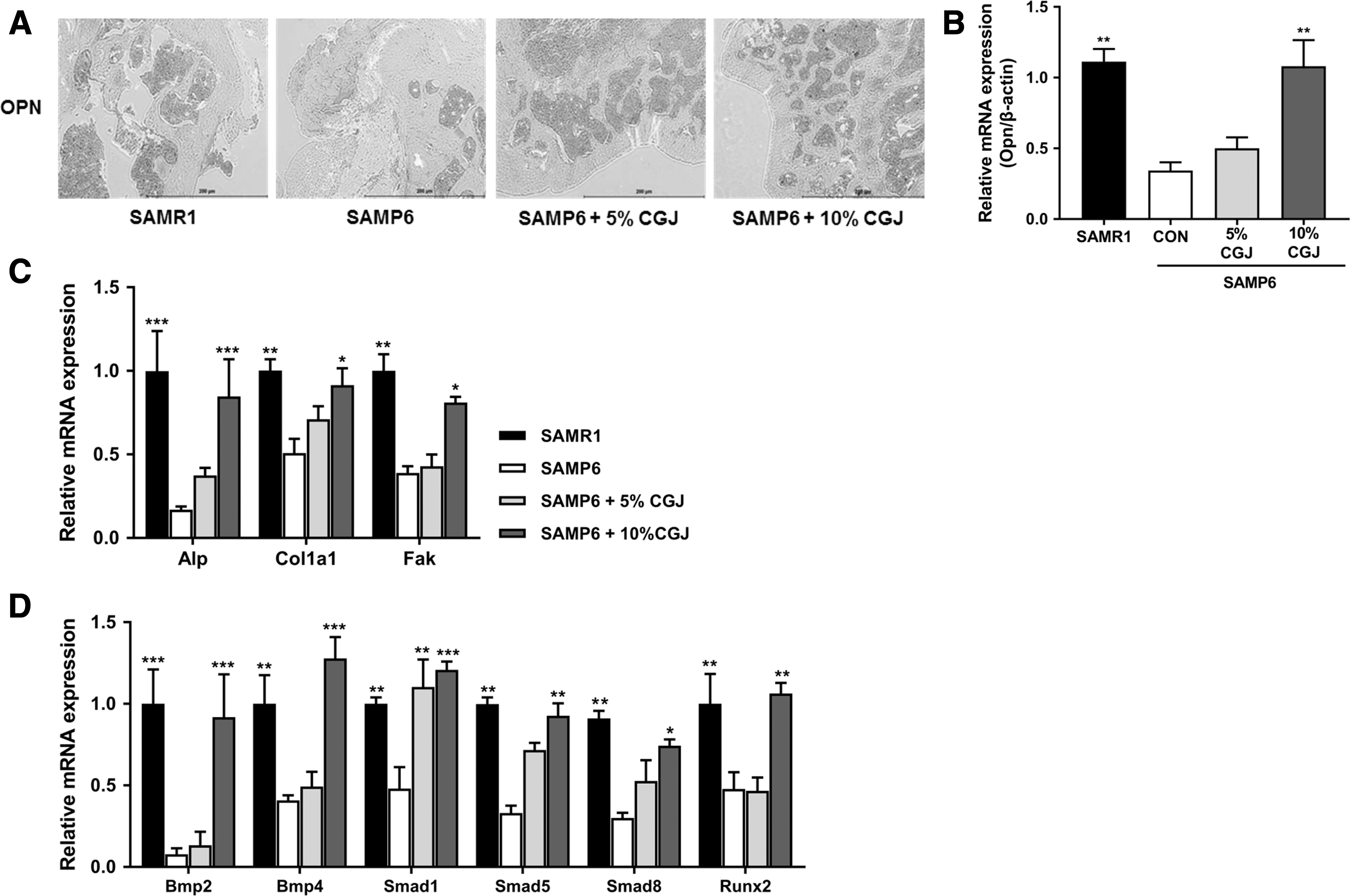
Effects of CGJ supplement on bone formation markers.
Effect of CGJ on bone resorption
To assess the effects of CGJ on bone resorption, we performed immunohistochemical staining and examined the mRNA expression in the femur. As given in Figure 5A, CTK reactivity was the highest in the SAMP6 control group, which was effectively decreased by CGJ treatment. Consistently, Ctk mRNA expression was significantly downregulated in the CGJ treatment groups compared with that of the SAMP6 control (Fig. 5B). Furthermore, the Opg mRNA expression was significantly upregulated in the 10% CGJ treatment group compared with that of the SAMP6 control. In contrast, Rankl and Nfatc1 mRNA expression was significantly downregulated in the CGJ treatment group compared with the SAMP6 control (Fig. 5C). Moreover, the ratio of Opg to Rankl relative gene expression levels was significantly higher in the CGJ treatment group than in the SAMP6 control (Fig. 5D).
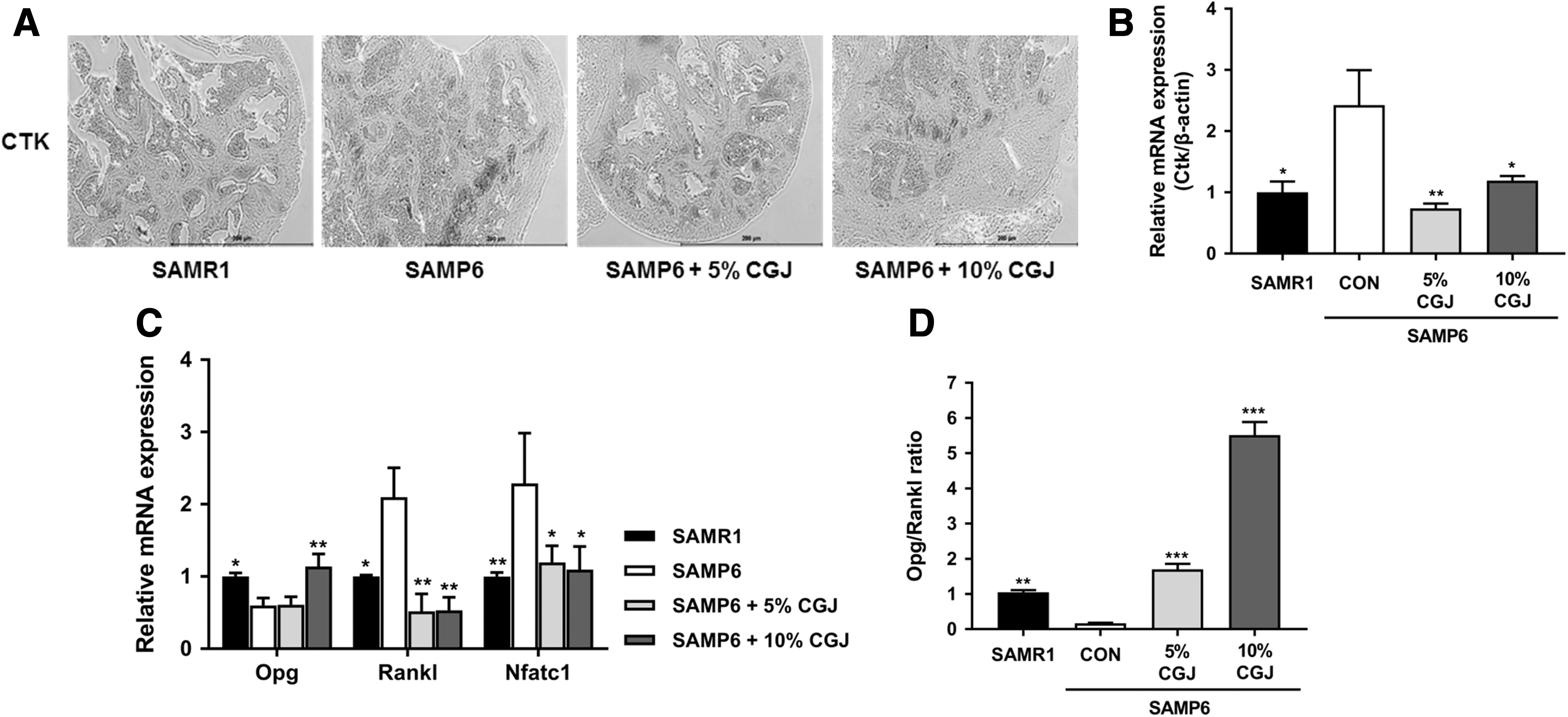
Effects of CGJ supplement on the osteoclastogenesis-induced bone resorption markers.
Discussion
Aging is associated with sustained bone loss, 1,4 which occurs as a result of two key processes: periosteal alignment outside the bone and bone resorption inside the bone. 33 Although periosteal alignment has a greater influence in women, both types of age-related bone loss result from changes in hormones, and cell number and function; hence, both men and women show similar age-related bone resorption patterns. 34 We previously reported that CGJ has high isoflavone bioavailability in OVX mice, and the constituent isoflavone metabolites stimulated ALP activity in MC3T3-E1 cells and attenuated RANKL-induced bone resorption activity in RAW264.7 cells. 12 In this study, we confirmed that CGJ contains abundant aglycone-type isoflavones, and provided evidence that CGJ may be an effective fermented soybean food for promoting bone health and senile osteoporosis prevention.
Fermentation is an excellent processing method for improving the nutritional and functional properties of soybeans by increasing the content of small bioactive compounds. 35 Isoflavones are present in soybeans and most soy-based foods primarily in the glycoside form cannot be absorbed by the body. 30 Once ingested, glycosides must be hydrolyzed to the absorbable aglycone form by β-glucosidase produced by the intestinal microflora. 36,37 Thus, isoflavone aglycones are absorbed faster and in higher amounts than their corresponding glycoside forms. 38 Isoflavone aglycone exists in some fermented soybean foods such as miso, natto, and tempeh, produced by microbial fermentation; however, the rate of production is not high. 30 Fujioka et al. reported that male mice fed with daidzein or genistein had a higher BMD than control mice and the plasma concentration of equol was increased after daidzein feeding in male and female mice. 39 Gao and Yamaguchi reported that daidzein and genistein induced an increase in ALP activity and Ca content in rat femoral bone, suggesting that isoflavone may have a bone anabolic effect. 40 We found that the isoflavone and aglycone contents in CGJ were 38.70 and 36.61 mg/100 g dry weight, respectively, and the rate of aglycone production was ∼94.5%. Especially, the contents of daidzein and genistein were increased the highest according to fermentation. Considering the average daily diet of the mice (4.5 g/day), the average daily isoflavone intake during the 15 weeks of feeding was ∼87–174 μg, suggesting the optimal amount of isoflavones in CGJ for ameliorating senile osteoporosis in SAMP6 mice.
Bone remodeling is regulated by two main activities: osteoclast-mediated bone resorption and osteoblast-mediated bone formation. 41 We found that CGJ supplementation significantly upregulated Igf-1 mRNA expression. IGF-1 plays a role in the commitment of mesenchymal stem cells to osteoprogenitor cells. 42 Furthermore, CGJ supplementation effectively upregulated Bmp2/4 and Smad1/5/8 mRNA expression. BMPs play important roles in bone metabolism by inducing bone formation, and some BMPs are clinically used in orthopedic treatment. 43 In osteoblast differentiation, BMP2, COL1A1, and OCN are involved in the formation, metabolism, and regeneration of bones. 44 BMP2, an important growth factor, regulates osteoblast differentiation by stimulating osteoblast-related transcriptional factors, including RUNX2. 45 In addition, SMAD1/5/8 specifically mediates BMP2 responses such as the osteoblastic differentiation of precursor cell lines. 46 Therefore, our results demonstrated that CGJ effectively improved osteoblast-induced bone formation through regulation of the BMP-SMAD-RUNX2 pathway.
Bone resorption results from osteoclast activity. 47 Osteoclasts constitute a differentiation product of osteoclast precursors, which is regulated by certain transcription factors and signaling pathways activated upon RANKL/RANK interactions. 48 CGJ supplementation downregulated the osteoclast-related bone resorption marker Ctk, which plays a critical role in the degradation of type 1 collagen during osteoclast-mediated bone resorption. 49 In addition, CGJ supplementation effectively downregulated Rankl and Nfatc1 mRNA expression. The expression NFATC1 is considered to be critical for the completion of osteoclast precursor differentiation by RANKL. 48 Furthermore, osteoprotegerin (OPG) is critical for suppressing the number of osteoclasts so as to prevent bone resorption. 38 CGJ supplementation not only upregulated Opg mRNA expression but also increased the Opg/Rankl ratio. These data confirmed that CGJ negatively regulates osteoclast-related gene expression, resulting in decreased bone resorption in SAMP6 mice.
In conclusion, we showed that CGJ, a Korean traditional short-term fermented soybean food, has abundant isoflavone aglycone content, and its supplementation in SAMP6 mice could effectively promote osteogenesis and inhibit osteoclastogenesis through the BMP2/SMADs/OPG pathway (Fig. 6). These results suggest that consumption of CGJ is a potentially promising alternative and complementary therapeutic food to prevent senile osteoporosis.

Proposed mechanism underlying the amelioration of bone turnover in senile osteoporosis upon CGJ supplementation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by Main Research Program of the Korea Food Research Institute (