
Abstract
The occurrence of allergy-mediated inflammatory diseases such as asthma and atopic dermatitis have increased, but comprehensive treatment remains difficult. Previous studies have shown that Schisandra chinensis Baill has antioxidant, antidiabetic, and antitumorigenic effects. Cyanidin 3-rutinoside (CR) is the major anthocyanin pigment of S. chinensis. However, the biological effects of CR have been rarely studied to date. Therefore, the aim of this study was to investigate the regulatory effects of CR on phorbol-12-myristate-13-acetate (PMA)/A23187-induced allergic inflammation in vitro. CR inhibited the secretion of inflammatory cytokines such as interleukin-6 and tumor necrosis factor-α, and it also suppressed the phosphorylation of nuclear factor-kappa B. These results show that CR ameliorated PMA/A23187-induced allergic inflammation via the suppression of inflammatory cytokines in HMC-1 cells. Therefore, CR has potential as a therapeutic agent for allergic diseases.
Introduction
Allergic diseases are one of the most common chronic diseases that involve a hypersensitivity reaction of the immune system. 1 Allergic diseases occur in conjunction with several allergy-mediated disorders such as atopic dermatitis (AD), allergic rhinitis, allergic arthritis, and allergic eczema. 2 These allergic diseases represent a public health problem due to the effects of allergic diseases on quality of lifestyle and medical expenditure. 3 In addition, the occurrence of allergic diseases has increased because of changing environments, lifestyle, and food habits. 4 H1 receptor antagonists, selective β2 agonists, and corticosteroids have been used to treat allergic diseases. However, these treatments have been known to have negative side effects such as drowsiness, anorexia, nausea, deterioration of the immune system, and bone loss. 5,6 Therefore, a treatment with high efficacy and reduced side effects is required.
Among allergic diseases, AD is a highly pruritic chronic allergic skin disease. 2 Dry skin, skin hypersensitivity, thickening of skin, and eczematous skin lesions are clinical symptoms of AD. 7 Genetic factors, environmental factors, an imbalance in the immune system, and skin barrier defects cause AD. However, the main cause of AD remains unclear. The occurrence of AD is sometimes caused by allergic sensitization to environmental allergens. 8 AD is related to the expression of inflammatory cytokines, chemokines, and other immune factors from mast cells, keratinocytes, and other immune cells. 9 One of the allergic reactions of AD, caused by immunoglobulin E (IgE) antibodies against the antigen, has the potential to develop into a fatal immune disorder. 4 Moreover, the immune system disorder such as an imbalance between T-helper (Th)-1 cells and Th-2 cells influences the development of AD. 10 AD patients have high secretion levels of serum IgE and immune system related cytokines such as interleukin (IL)-4, IL-6, and IL-13. The affected keratinocytes and immune cells produce chemokines and proinflammatory cytokines that lead to the invasion of immune cells into skin lesions. 11 However, further study of the regulation of allergy-disease mechanism such as the regulation of inflammatory cytokine is very important.
Allergic responses from AD are initiated by mast cell infiltration and activation. The mast cell is exposed to antigen release of histamine and other cytokines such as IL-6, IL-8, and tumor necrosis factor (TNF)-α. 12 These cytokines are secreted by mast cells and are regulated by activated transcription factors, such as nuclear factor-kappa B (NF-κB). 13 Furthermore, inflammatory mediators, such as protease-activated receptor2 (PAR2), mitogen-activated protein kinases (MAPKs), and thymic stromal lymphopoietin (TSLP) are related to the intensity degree of allergic inflammation. 14
Schisandra chinensis Baill is a member of the family Schisandraceae. The fruit of S. chinensis has been traditionally used to treat asthenia, asthma, cough, and exhaustion in patients in Asian countries. 15 Recent studies have reported that S. chinensis inhibits hepatocarcinogenesis, 16 affects antioxidant activity, reduces blood pressure, 17 and has an anti-inflammatory effects on mouse edema. 18 The major components of S. chinensis include schisandrin, deoxyschisandrin, and gomisin A, which have been reported to have pharmacological effects. 19 However, the pharmacological effects of the anthocyanins family in S. chinensis are not well understood. Cyanidin 3-rutinoside (CR) is one of the most abundant constituents in fruits and vegetable. It has been reported that CR has pharmacological effects, including antioxidant, 20 antidiabetic, 21 and antitumorigenic effects. 22 However, the effect of CR on allergic inflammation has not been reported to date.
The aim of the present study was to investigate the anti-inflammatory effects of CR and the underlying mechanism in an allergic inflammation experimental model. We evaluated the effects of S. chinensis extract (SCE) on 2,4-dinitrofluorobenzen (DNFB)-induced AD-like dermatitis in vivo and the effects of CR on allergic inflammation in vitro via inflammatory cytokine secretion.
Materials and Methods
Reagents
CR was purchased from TOKIWA Phytochemical Co. (Sakura, Chiba, Japan). Compound 48/80, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium bromide (MTT), calcium ionophore A23187, and phorbol 12-myristate 13-acetate (PMA) were purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). Fetal bovine serum (FBS), Iscove's Modified Dulbecco's Medium (IMDM), and penicillin streptomycin were purchased from Gibco BRL (Grand Island, NY, USA). Phospho-JNK, -ERK, -p38, and NF-κB p65 were purchased from Cell Signaling Technology (Danvers, MA, USA). ERK, JNK, p38, Glyceraldehyde 3-phosphate dehydrogenase (GAPDH), and β-actin antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Anti-rabbit and mouse secondary antibodies were purchased from Jackson Immuno Research (West Grove, PA, USA).
Preparation of SCE
S. chinensis was purchased from Human Herb (Gyeongsan, Gyeongsangbuk-do, Korea). S. chinensis fruits (150 g) were added to distilled water (1.5 L) and extracted at 60°C for 8 h. The extract was filtered through a 0.22 μm syringe filter (HYUNDAI Micro, Seoul, Korea) and then lyophilized at −40°C on 5 mTorr. The yield of SCE was 28.7% (43.055 g).
Cell culture
The human mast cell line (HMC-1) was generously provided by HJ Ahn (Sangji University, Wonju, Korea). HMC-1 cells were cultured in IMDM contained with 10% FBS, 100 U/mL of penicillin, and 100 μg/mL of streptomycin and incubated in a humidified atmosphere of 5% CO2 at 37°C.
The cell viability of CR was measured by MTT assay. In brief, HMC-1 cells were seeded at 5 × 105 cells/well in 24-well plates and then incubated for 24 h. CR was treated with various concentrations (0.01–1000 μg/mL) for 24 h. The MTT solution was treated into each well of the plate, and the plate was incubated over 4 h at 37°C. Formazan crystal was dissolved with 600 μ of DMSO and the absorbance was read at 540 nm using a VersaMax microplate reader (Molecular Devices, CA, USA).
Enzyme-linked immunosorbent assay
HMC-1 cells (5 × 105 cells/well) were seeded into 24-well plates. HMC-1 cells were treated with various concentrations (0.1–100 μg/mL) of CR and, after 4 h, the HMC-1 cells were stimulated with PMA (5 ng/mL) and A23187 (10 ng/mL) for 18 h. For the enzyme-linked immunosorbent assay (ELISA) experiment, the IMDM containing HMC-1 cells was centrifuged at 15,520 g for 10 min, and then the supernatants were collected as the ELISA sample. ELISA kits (BD Pharmingen, San Diego, CA, USA) were used to determine the effects of CR on the secretion of inflammatory cytokines. In brief, 96-well plates were precoated with capture antibody and incubated at 4°C overnight. After that, plates were washed with 0.05% phosphate buffer saline-Tween20 (PBST) three times and blocked with 5% FBS in PBS was added and incubated at room temperature (25–30°C) (RT) for 1 h. After washing three times with 0.05% PBST, samples and standards were added into the 96-well plates and incubated for 3 h at RT. Plates were washed five times, and 100 μL of working detector (detection Ab+SAv-HRP) was added to each well and incubated at RT for 1 h. After washing five times, 100 μL of substrate solution was added to each well and the plates were incubated at RT for 15 min in the dark. Absorbance was measured at 450 nm using a VersaMax Microplate Reader.
Western blot analysis
The collected cells were washed with ice-cold PBS, lysed by lysis buffer (iNtRON Biotech, Seoul, Korea) for 1 h, and centrifuged at 15,520 g for 10 min at 4°C. The supernatants were collected and transferred to sterile tubes. Protein concentrations were quantified using a DC Protein Assay Kit (BIO-RAD Laboratories, California, USA). Lysates (25 μg of protein) were mixed with a 2 × sample buffer, boiled at 95°C for 5 min, subjected to 10% SDS-polyacrylamide gel electrophoresis, and then transferred to polyvinylidene difluoride (PVDF) membranes. The membranes were blocked with 5% skim milk in 0.05% PBST for 1 h 30 min and incubated with primary antibody at 4°C overnight. After washing with 0.1% PBST for 1 h, the membranes were incubated with secondary antibodies for 45 min at RT. After washing with 0.1% PBST for 2 h, the membranes were visualized using ECL Western Blotting Luminol Reagent (Santa Cruz Biotech, CA, USA). The protein bands were detected using a LAS-4000 image reader (Fujifilm Life Science, Tokyo, Japan).
Real-time polymerase chain reaction
Total RNA was isolated from HMC-1 cells using an easy-BLUE Reagent Kit (iNtRON Biotech). Total cellular RNA was used as a template for the first-strand cDNA synthesis using a 2 × RT premix cDNA Synthesis Kit (Bio FACT, Daejeon Korea) according to the manufacturer's directions. The transcription levels of genes were evaluated with an AriaMX real-time polymerase chain reaction (real-time PCR) system (Agilent Technologies, CA, USA). The relative mRNA gene expression was calculated using the comparative CT method with AriaMX Real-Time PCR Software (Agilent Technologies, CA, USA). The primer sequences used for real-time PCR are shown in Table 1.
Primers Utilized for Real-Time Polymerase Chain Reaction
The DNFB-induced AD-like dermatitis mice model
All experimental protocols (CBNU2017-0089) were approved by the Committee on the Care of Laboratory Animal Resources, Chonbuk National University and were conducted in accordance with the Guide for the Care and Use of Laboratory Animals. Male BALB/c mice (6 weeks old, 21–23 g) were purchased from SAMTAKO (Osan, Korea). The mice were housed in a laminar air-flow room maintained at a temperature of 25°C ± 2°C and humidity level of 55% ± 1% with a 12 h light–dark cycle.
The dorsal skin of the mice (n = 7) was shaved with depilatory creams before the experiment. The mice were treated with a vehicle or 100 μL of 0.15% DNFB in acetone/olive oil (3:1) to the dorsal skin twice per week for 5 weeks. The experimental mice were divided into four groups as follows: blank (nontreatment), DNFB (0.15% DNFB only), SCE 10 (0.15% DNFB+SCE 10 mg/kg), and SCE 100 (0.15% DNFB+SCE 100 mg/kg). The SCE groups were orally administered SCE 4 weeks after DNFB application.
Compound 48/80-induced anaphylactic shock
The 5 mg/kg of compound 48/80 intraperitoneally injected in the experimental mice (n = 8). SCE (10 and 100 mg/kg) was orally administered 1 h before injection of the compound 48/80. Survival rate was monitored for 1 h after the systemic anaphylactic shock was induced.
Liquid chromatography-mass spectrometry analysis of SCE and CR
The water extracts of S. chinensis and CR were dissolved in water and MeOH, respectively. High-performance liquid chromatography was performed on an Agilent 1100 system (Agilent Technologies, Waldbronn, Germany) with an Agilent 1100 series quaternary pump (G1311A), a photodiode array detector DAD (G1315D), and an Agilent 6410 Triple Quadrupole mass spectrometer (Agilent Technologies) coupled with an electrospray ionization (ESI) interface and an ion trap mass analyzer. The ESI source was operated in positive ionization mode.
The extract and CR was analyzed under the following conditions: column, TSK-gel ODS-80Ts (4.6 × 150 mm; Tosoh Co., Tokyo, Japan); mobile phase, 0.1% formic acid (solvent system A), and acetonitrile (CH3CN, solvent system B) in a gradient mode (B from 10% to 70% in 30 min); sample injection volume, 10 μL; flow rate, 0.5 mL/min; column temperature, 25°C, UV wavelength, 520 nm. High-purity nitrogen was used as the dry gas at a flow rate of 10 L/min, gas temperature of 300°C; and fragmentor voltage of 200 V. Nitrogen was also used as the nebulizer at 30 psi and capillary voltage of ±4000 V.
Statistical analysis
Results are shown as a summary of data from at least three experiments. All results are presented as the mean ± standard error of the mean (S.E.M). Results were analyzed using the Graph Pad Prism version 5.0 software program (Graph Pad software, Inc., La Jolla, CA). One-way analysis of variance with Tukey's post hoc test was used to determine statistically significant differences. P < .05 indicated significance.
Results
Effects of SCE on DNFB-induced AD-like dermatitis in vivo
To determine the effect of SCE on DNFB-induced AD-like skin lesions, these skin lesions in mice were observed 5 weeks into the experimental period. Symptoms in the DNFB group were higher than that in the blank group. However, the 0.15% DNFB-induced symptoms of AD-like dermatitis were alleviated by administration of the 100 mg/kg SCE treatment (Fig. 1A).

Effects of SCE on DNFB-induced AD-like dermatitis in vivo. Comparison of DNFB-induced AD-like dermatitis in BALB/c mice after oral administration of SCE (10 and 100 mg/kg)
In hematoxylin and eosin-stained skin tissue, the skin thickness was thicker than that of the blank group in the DNFB group. The thickened skin in the 0.15% DNFB application was alleviated by the oral administration of SCE (Fig. 1B). In the toluidine blue-stained tissues, infiltration of mast cells on skin tissue was higher in the 0.15% DNFB application than that in the blank group. The SCE orally administered group showed inhibition of the number of infiltrated mast cells into the dermis (Fig. 1B).
To investigate the effect of SCE on the systemic allergic reaction, the compound 48/80-induced anaphylactic shock mice model was used. SCE oral administration regulated the survivor rate and induced compound 48/80 in 30 min (Fig. 2). The serum IgE level was higher in the 0.15% DNFB treatment than that in the blank group. However, the serum IgE level was significantly reduced by oral administration of 100 mg/kg SCE (Fig. 1C). The histamine release in the serum was inhibited by the 0.15% DNFB treatment. However, the increased histamine release was significantly suppressed by SCE oral administration compared to that in the DNFB group (DNFB; 24.70 ± 6.78 ng/mL, SCE 10; 10.56 ± 0.74 ng/mL, and SCE 100; 8.65 ± 2.48 ng/mL) (Fig. 1D).
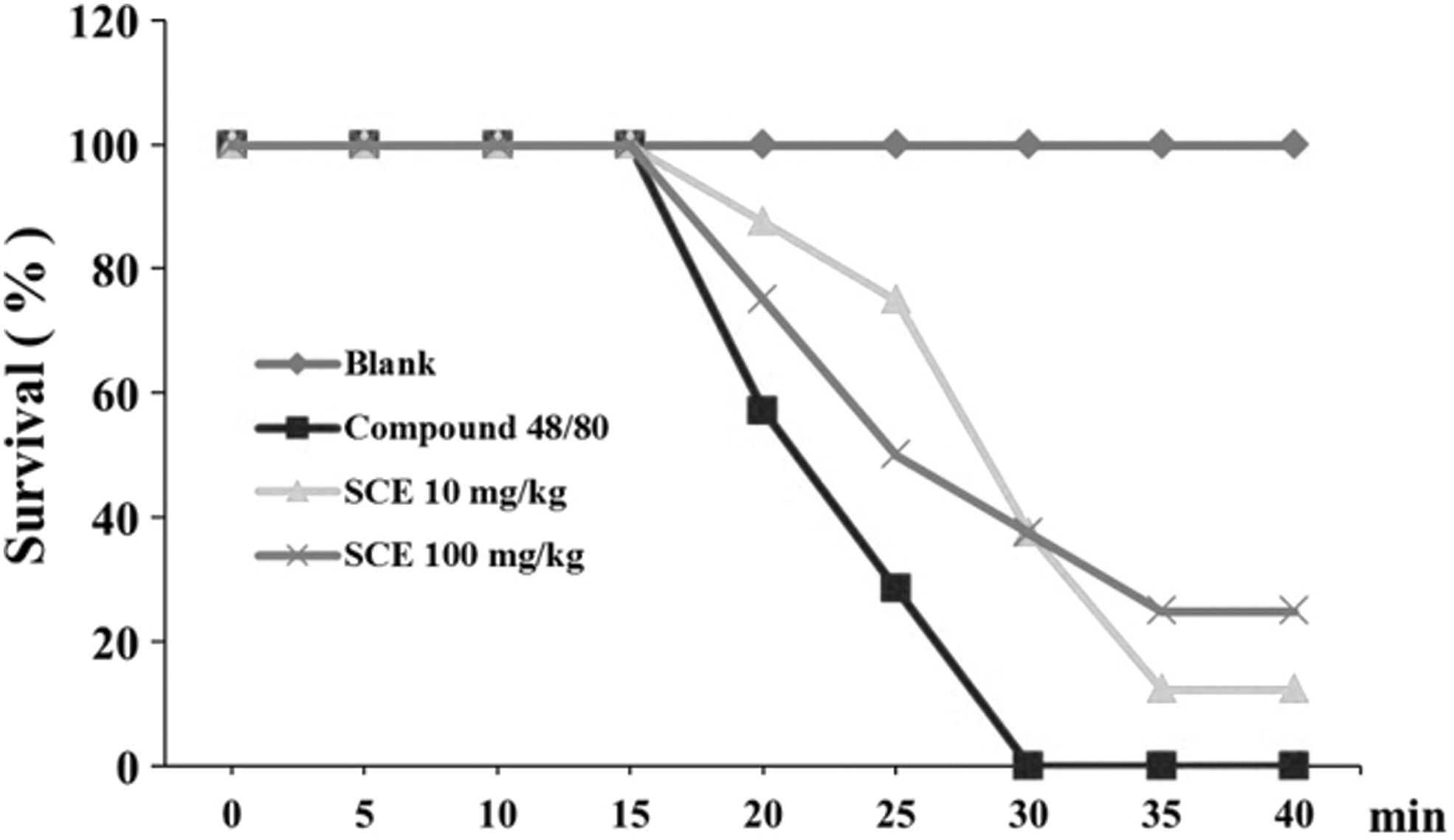
Inhibitory effect of SCE on compound 48/80-induced systemic anaphylactic shock. Four groups of mice (n = 7/group) were orally administered saline, SCE (10 mg/kg), SCE (100 mg/kg) at 1 h before injection of compound 48/80. The compound 48/80 was intraperitoneally given to mice. Survival rates (%) were monitored for 1 h after induction of anaphylactic shock.
Quantification of CR in SCE
When analyzed using liquid chromatography-mass spectrometry (LC/MS), the retention time of CR was 10.1 min. The CR in the water extract of the S. chinensis fruit was identified with a mass spectrum and retention time of the standard. The mass spectrum of CR showed a molecular ion peak at m/z 595 [M = H]+ and a fragment ion peak at m/z 287 similar to the results from a previous study. 23 Quantitative analysis of CR by the extracted ion chromatogram showed a concentration of 3.48 mg/g in the extract (Fig. 3).

Extracted ion chromatogram of SCE
Effects of CR on the secretion of inflammatory cytokines in PMA/A23187-stimulated HMC-1 cells
Mast cells play an important role in the allergic response via the secretion of immune mediators and cytokines. 24 Therefore, we determined whether CR regulated the secretion levels of inflammatory cytokines. The various concentrations of CR (0.1–100 μg/mL) showed noncytotoxicity in HMC-1 cells (Fig. 4A). The stimulation of PMA (50 nM) and A23187 (1 μM) significantly increased the secretion levels of IL-1β, IL-6, and TNF-α. The increased secretion levels of IL-1β, IL-6, and TNF-α were effectively suppressed by the CR treatments (Fig. 4B–D). In addition, the increased mRNA expression levels of IL-1β, IL-6, and TNF-α by PMA/A23187 stimulation were significantly decreased by CR treatments (Fig. 4E–G).
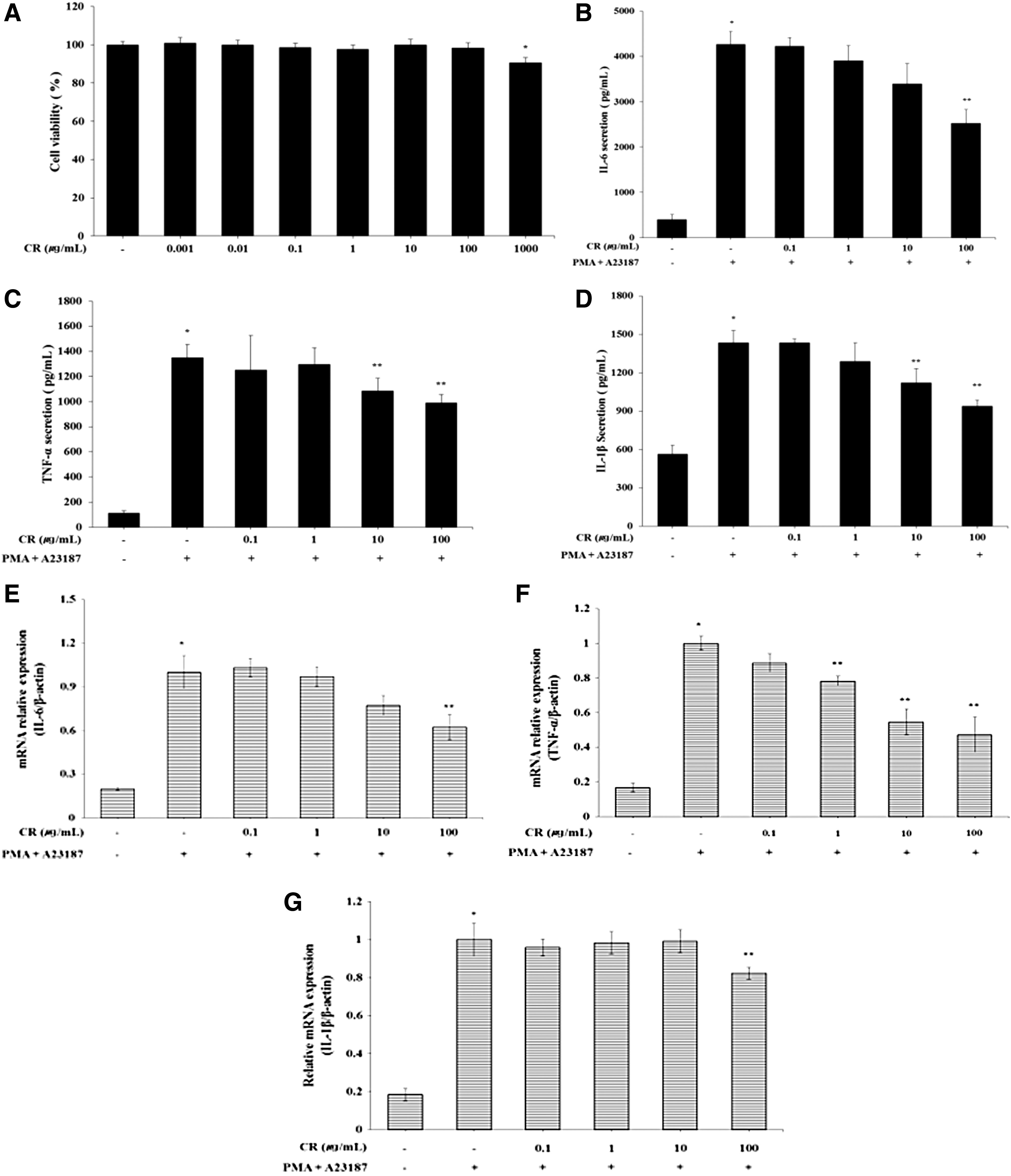
Effects of CR on the secretion of inflammatory cytokines and the phosphorylation of MAPKs and NF-κB in PMA/A23187-stimulated HMC-1 cells. The cytotoxicity of CR (0.001–1000 μg/mL) on HMC-1 was analyzed using MTT assay
Effects of CR on the phosphorylation of MAPKs and NF-κB in PMA/A23187-stimulated HMC-1 cells
The phosphorylation of MAPKs and NF-κB induced the secretion of inflammatory cytokines in mast cells. The regulation of the MAPKs and NF-κB signaling pathway is important in regulating mast cell-mediated inflammation. 25 The CR treatment considerably decreased PMA/A23187-induced phosphorylation of JNK, ERK, and p38 (Fig. 5A). In addition, CR inhibited the phosphorylation of NF-κB p65 (Fig. 5C).
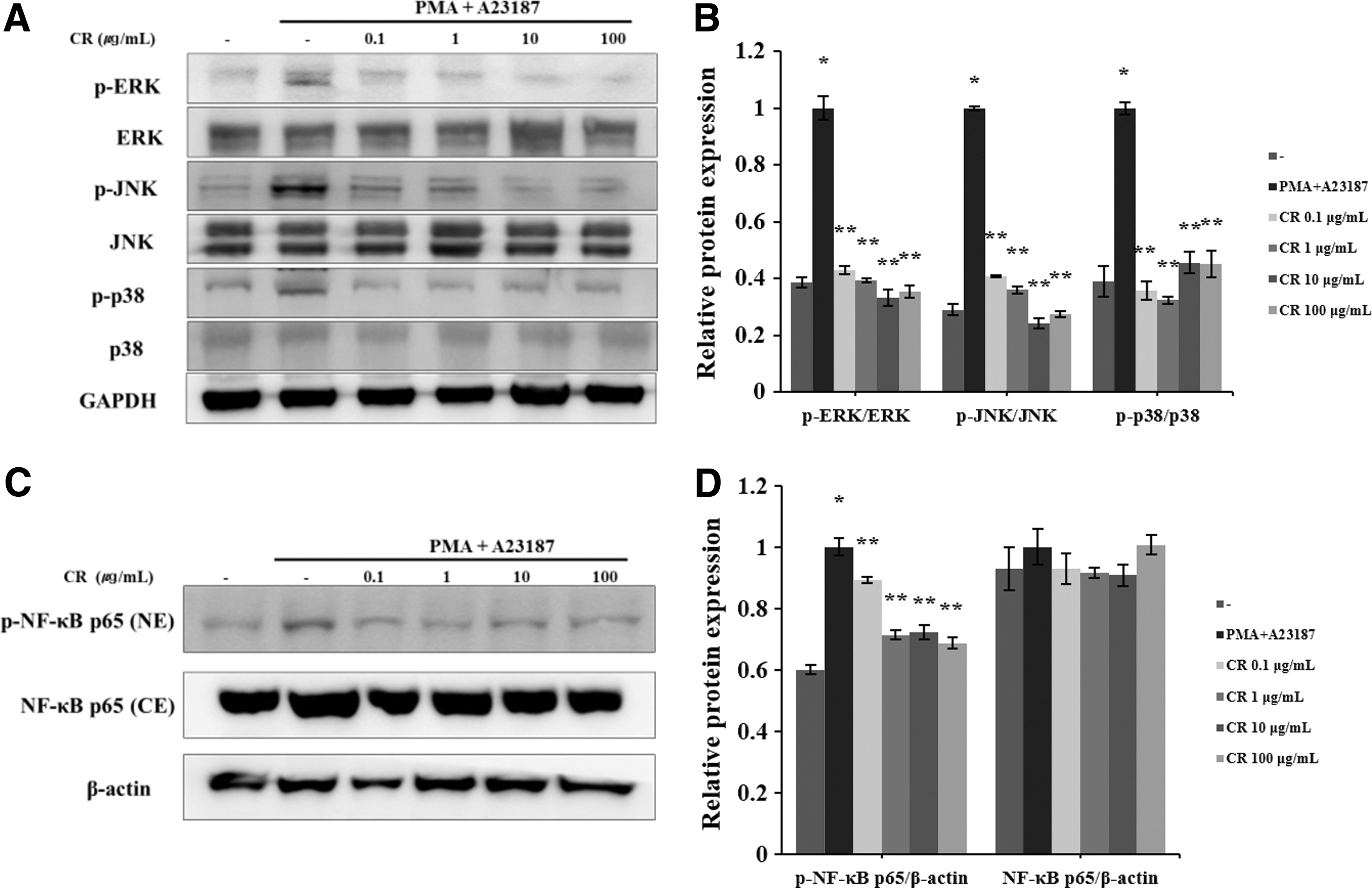
Effect of CR on the MAPK and NF-κB signaling pathway in HMC-1 cells. Phosphorylation of MAPKs and the NF-κB
Effects of CR on allergy mediators in HMC-1 cells
T lymphocytes, eosinophils, keratinocytes, chemokines, and cytokines are involved in the progression of atopic diseases. 26 In addition, the inhibition of caspase-1 is the method for treating patients with allergic inflammatory diseases. 27 The activation of caspase-1 was induced by PMA/A23187 stimulation. The activated caspase-1 was suppressed by CR treatment (Fig. 6). In addition, allergic inflammation mediators such as TSLP and histamine were regulated by various concentrations of CR (Fig. 7). The mRNA level of TSLP was increased by PMA/A23187 stimulation. The increased TSLP mRNA level was decreased by 100 μg/mL of CR treatment. PMA/A23187 stimulation increased histamine release (Blank; 0.98 ± 0.13 ng/mL, PMA/A23187; 4.73 ± 0.16 ng/mL). CR treatment downregulated increased histamine release dose dependently (PMA/A23187; 4.73 ± 0.16 ng/mL, CR 1 μg/mL; 3.78 ± 0.80 ng/mL, CR 10 μg/mL; 3.17 ± 0.40 ng/mL. CR 100 μg/mL; 2.06 ± 0.30 ng/mL).

Effects of CR on caspase-1 expression in PMA/A23187-stimulated HMC-1 cells. The expression level of caspase-1 was determined using western blot analysis

Effects of CR on allergic mediators in PMA/A23187-stimulated HMC-1 cells. The relative cytokine mRNA expression level of TSLP was determined using real-time PCR
Discussion
Allergic responses are classified into early and late phases, both of which result in chronic allergic inflammation. 28 During the early phase, the release of histamine and other mediators bind IgE to allergens in mast cells. 29 One characteristic of the late phase is the recruitment of leucocytes that results in upregulation of inflammatory cytokines. 30 In the present study, the authors investigated the effect of CR on allergic inflammation. AD, chronic allergic disease, involves local itching and dry skin. Th-2 cell-mediated cytokines, mast cells, and IgE are involved in the progression of AD. 26
Mast cells are related to allergic inflammation and are considered to play a major role in AD progression because an increase in mast cells has been detected in AD patients. 31 Activated mast cells produce various inflammatory mediators such as histamine, IgE, IL-1β, IL-6, TNF-α, and chemokines. 32 HMC-1 cells are the typical cell line for studying the pathway of allergic inflammation. After PMA and calcium ionophore A23187 stimulation in HMC-1, immediate hypersensitivity and multiple cytokines are produced. 33 Inflammatory cytokines such as IL-6, IL-1β, and TNF-α are secreted by the allergic condition and promote tissue fibrosis and influence chronic inflammatory diseases. 34 In the present study, the CR treatment regulated the inflammatory cytokine secretion in PMA/A23187 stimulated HMC-1 cells (Fig. 4).
In chronic allergic disease conditions, allergic inflammatory mediators such as NF-κB, TSLP, and MAPKs expressions are related to the severity of the allergic inflammation. 14 The activation of MAPKs is related to the allergic response via phosphorylation of NF-κB, which causes the secretion of inflammatory cytokines and chemokine. 35 TSLP, an epithelial-derived cytokine, sustains Th-2-type allergic inflammation. In addition, TSLP expression is influenced by PAR2 and NF-κB activation. 36 As shown in Figures 3 and 4, CR treatment of HMC-1 cells significantly suppressed MAPKs and NF-κB expression.
CR is a natural anthocyanin pigment that is present in high proportions in blackberry, raspberry, and S. chinensis. CR has been reported to possess biological activities, including antioxidant, antihyperglycemic, and antidiabetic effects. 37 CR is not the most abundant compound in SCE; however, it shows variation in quantity depending on the fruit color. 38 The color as well as biological activities could be changed by the composition of anthocyanins in quantity and type. In the present study, CR was detected in SCE by LC/MS analysis (Fig. 3).
In conclusion, CR may have potential as an effective candidate for the treatment of allergic diseases. Further, CR regulated allergic inflammation via the inhibition of inflammatory cytokine secretion and allergic mediators. However, the specific mechanistic pathway is still not clear and further studies are required.
Footnotes
Acknowledgments
This research was supported by the Korean Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry, and Fisheries through the Agri-Bioindustry Technology Development Program, funded by the Ministry of Agriculture, Food, and Rural Affairs (315033-03). This research was supported by “Research Base Construction Fund Support Program” funded by Chonbuk National University in 2017.
Author Disclosure Statement
No competing financial interests exist.