
Abstract
This study researched the effects of Lactobacillus plantarum PMO 08 alone and combined with chia seeds on metabolic syndrome and parameters related to microbiota modulation and intestinal barrier integrity in obese mice fed high-fat diets (HFDs; 45% kcal fat). Male C57BL/6J mice were acclimated for a period of 2 weeks and then randomly separated into five groups depending on whether they received a normal diet (ND group), an HFD (HFD group), an HFD with L. plantarum (PMO group), an HFD with L. plantarum combined with chia seeds (PMOChia group), or an HFD with chia seeds (Chia group). Serum lipid profiles and related markers (cholesterol metabolism-related gene expression) were measured. Intestinal barrier integrity was assessed by measuring occludin mRNA expression of tight junction proteins. Mucosal bacteria were checked with quantitative reverse transcript polymerase chain reaction (qRT-PCR). After 16 weeks of feeding, the PMO group showed significantly lower serum total cholesterol, low-density lipoprotein cholesterol levels, atherogenic index, and cardiac risk factors compared to the HFD group. Moreover, the hepatic mRNA expression of SREBP2 (sterol regulatory element binding protein 2), a protein related to cholesterol metabolism, was significantly downregulated in the PMO group. We also found a positive synergistic effect in the PMOChia group, as manifested by the hepatic mRNA expression of hepatic CYP7A1 (cholesterol 7α-hydroxylase), strengthening of the gut barrier function, and the promotion of more L. plantarum in the colonic mucosa than in either the HFD or PMO group. In conclusion, our results indicate that PMO 08 may protect against metabolic syndrome by exerting effects on the regulation of lipid metabolism. Although the effects of chia seeds alone remain uncertain based on this experiment, its combination with PMO 08 was demonstrated to improve multiple beneficial effects of PMO 08 in obese mice fed HFD, which is a promising possibility for future research.
Introduction
Growing evidence suggests that dysbiosis of the gut microbiota is associated with obesity and related metabolic disorders. 1 –4 The mammalian gut is colonized with a varied range of microorganisms that play a significant role in host metabolism. Therefore, it has been shown that manipulation of microbiota in mice using probiotics has beneficial effects on obesity, diabetic mellitus, and metabolic syndrom. 5,6
Probiotics contain living microorganisms that are beneficial to the host when administered in adequate amounts. 7 They are beneficial for several common health issues, such as constipation, irritable bowel syndrome, diarrhea, allergic disease, weight loss, glycemic control, and lipid metabolism. 8,9 Recently, probiotics have been introduced as a natural therapeutic intervention with the potential for ameliorating metabolic syndrome. 10
Despite the fact that the molecular mechanism of probiotic actions has not been fully elucidated, many of their effects may prove helpful in treating obesity and related disorders. For example, Lactobacillus plantarum HAC01 inhibited the enlargement of mesenteric adipose tissue in high-fat diet (HFD)-fed obese mice 11 ; L. plantarum MTCC5690 improved gut integrity, decreased systemic levels of lipopolysaccharide (which exerts a proinflammatory effect), increased glucagon-like peptide-1 levels, reduced endoplasmic reticulum stress, and subsequently improved peripheral insulin sensitivity in an HFD-fed mouse model 12 ; and L. plantarum K21 inhibited fat mass accumulation, strengthened gut barrier function, and modulated the composition of gut bacteria. 13 Thus, it is crucial to demonstrate the functional efficacy of various probiotic strains, as each exerts distinct effects on parameters related to metabolic syndrome. The functional efficacy of L. plantarum PMO 08 in this aspect has not been investigated, and hence, this study was conducted to fill this research gap.
L. plantarum has a long history of safe use in fermented foods. Due to its high tolerance to acid and bile salt, L. plantarum strains show a high survival rate after human ingestion. 14 L. plantarum PMO 08 is a Korean native lactic acid bacteria isolated from kimchi, a traditional form of Korean fermented cabbage. It has excellent acid and bile tolerance, strong bile salt hydrolase (BSH) activity, and high antibacterial and intestinal attachment ability. 15 –17 Lactobacilli with BSH activity can survive to inhabit the lower small intestine, an important site for the enterohepatic cycle of cholesterol. Thus, BSH activity is acknowledged to be an important colonization factor and a crucial criterion for the selection of a probiotic with cholesterol-lowering properties. 18 Hence, considering the ability of L. plantarum PMO 08 to enzymatically deconjugate bile acids, we assessed its potential in vivo as a cholesterol-lowering probiotic.
The definition of prebiotic is “a selectively fermented ingredient that results in specific changes in the composition and/or activity of the gastrointestinal microbiota, therefore conferring benefits upon host health.” 19 Some of the beneficial effects of prebiotics are the promotion of good bacterial growth such as Lactobacillus, and/or Bifidobacterium, the fortification of gut barrier function, and lowering the risk of obesity. 20 –22 The most common prebiotics are galacto-oligosaccharides, fructo-oligosaccharides, inulin, β-glucan, and psyllium husk. 23 Up until now, many studies into prebiotic efficacy of dietary fiber have emphasized the effects of isolated individual fibers, and less had been conducted into the potential prebiotic benefits of consuming intact cereal fiber in daily diet. Nowadays, researchers are interested in cereal fiber such as oat, barley, and wheat bran.
Chia seeds (Salvia hispanica L.), which contain appreciable amounts of fermentable fiber, could be used as prebiotics. It belongs to the mint family Lamiaceae and it is native to southern Mexico and northern Guatemala. 24 Thousands of years ago, chia seeds were already a staple food and they were consumed by pre-Columbian people who lived primarily in Central America. Chia seeds contain ∼30–34 g/100 g dietary fiber, of which the insoluble dietary fiber accounts for ∼85–93%, while the soluble fraction is about 7–15%, and this fiber content is higher than oat, barley, and flaxseed. 24,25 Higher contents of fiber improve bowel movements, decrease serum cholesterol, and alleviate symptoms of diabetes mellitus by decelerating the digestion process and release of glucose. 26 Some studies have reported that consumption of chia seeds led to health benefits, such as weight control, reducing blood pressure, and improving insulin sensitivity. 27 –31
As mentioned above, since chia seeds contain a large amount of dietary fiber, it is thought that they may be a good source of enteric fermentation in the intestinal tract and may show improvement in metabolic disease management by changing the intestinal microbiota. 32 However, the impact of chia seed as a prebiotic compound on promoting lactobacilli or other beneficial microbiota has not yet been evaluated. Hence, this study explored how chia seeds contributed to preventing metabolic syndrome by regulating the gut microbiota balance.
In this study, we investigate whether diets with probiotic L. plantarum PMO 08 alone and L. plantarum PMO 08 combined with chia seeds exerted beneficial metabolic effects in obese mice on HFD. Also, to elucidate the mechanism of PMO 08 that combats dyslipidemia, we measured gene expression relevant to lipid metabolism and inflammation. In addition, the measurement of mucosal L. plantarum content and gut barrier integrity is included, as the health-promoting effect of L. plantarum PMO 08 as a probiotic has not yet been proven.
Materials and Methods
Preparation of lactobacilli and chia seeds
The L. plantarum PMO 08 and chia seeds used in this study were provided by the R&D Center of Pulmuone Co. Ltd. (Seoul, Korea). L. plantarum PMO 08 was grown in MRS broth (Difco, USA) for 18 h at 37°C, and subcultured two times before each experiment. Chia seeds were ground and preserved at −80°C until further processing.
Animals
Six-week-old male C57BL/6J mice were obtained from DooYeol Biotech (Seoul, South Korea). Mice were housed two or three per cage and maintained in a temperature (22°C ± 3°C)- and humidity (50% ± 10%)-controlled room with a 12-h light/12-h dark cycle. The experiments were carried out in accordance with the guidelines of the Institutional Animal Care and Use Committee (IACUC) of Seoul National University (Permission No. SNU-150813-2).
Experimental design
After 2 weeks of acclimatization, mice were randomly separated into 5 groups (n = 10 mice/group) and fed with the following diets for 16 weeks: the normal diet (ND) group was on a standard diet (10% of calories from fat), the HFD group was fed an HFD (45% of calories from fat) as a control group, the PMO group received this HFD supplemented with L. plantarum PMO 08, the PMOChia group was fed an HFD and 4% (w/w) chia seeds with L. plantarum PMO 08, and the Chia group was fed an HFD with 4% (w/w) chia seeds in their diet. The chemical composition of the experimental diets is shown in Table 1. All of the test materials were added to the feed, except for L. plantarum PMO 08, which was supplied in tap water (0.1 billion CFU/mL). The water was kept at room temperature (19–23°C) and replaced every 48 h. The mice were enabled to freely consume their preweighed feed and water in each cage.
Composition of the Experimental Diets
Chia seeds were powdered after being freeze-dried in intact form (chia seed nutritional values: total energy: 393 kcal/100 g, carbohydrates: 46.03 g/100 g, proteins: 21.09 g/100 g, fats: 20.08 g/100 g, dietary fiber: 37.23 g/100 g).
Chia, HFD +4% chia seed powder; HFD, high-fat diet (45% fat); ND, normal diet; PMO, HFD+Lactobacillus plantarum PMO 08 (supplied in tap water); PMOChia, HFD +4% chia seed powder+L. plantarum PMO 08 (supplied in tap water).
Body weight and feed intake measurements
The animals were supplemented for a period of 16 weeks. In the experimental period, body weight was recorded once every 2 weeks. The remaining diet feed was weighed every day for three consecutive days every week of the feeding period to measure food intake.
Blood and tissue collection
After the end of the experimental period, the mice were fasted for 16 h and sacrificed. Blood samples were collected by cardiac puncture and centrifuged for 20 min at 1000 g at 4°C. After blood collection, the liver, adipose tissue (epididymal fat), and colonic tissue were promptly removed, rinsed with phosphate-buffered saline, and weighed. Serum and tissues were quickly frozen in liquid nitrogen and preserved at −80°C in a deep freezer until further analysis.
Serum metabolic parameters, the atherogenic index, and the cardiac index
Serum total cholesterol, HDL cholesterol, and triglyceride levels were analyzed using a kit obtained from Asan Pharmaceutical Co. (Yongin, Republic of Korea). Low-density lipoprotein (LDL)-cholesterol was calculated by Freidewald et al.'s formula. 33 Fasting blood glucose levels were checked with an Accu-chek Active (Roche Diagnostics, Indianapolis, IN, USA). Serum insulin was determined with a mouse insulin ELISA kit (Shibayagi Co., Gunma, Japan). The homeostasis model assessment-estimated insulin resistance (HOMA-IR) index was calculated using the following formula: HOMA-IR = [fasting glucose (mg/dL) × fasting insulin (uU/mL)]/22.5. The atherogenic index (AI) was calculated using the following formula: AI = ([Total cholesterol] − [HDL cholesterol])/[HDL cholesterol]. The cardiac index was calculated by the following formula: CI = (Total cholesterol/HDL cholesterol).
Hepatic triglyceride and cholesterol
Hepatic total lipids were extracted according to the method of Folch et al. 34 50 mg of liver tissues were homogenized with chloroform: methanol (2:1, v/v) and then, KCl solution (0.88%) was added to the mixture. The extract was then centrifuged for 15 min at 1000 g at 4°C and evaporated to dryness under nitrogen. The residue was suspended in the isopropanol and kept in −80°C before analysis. Liver triglyceride and cholesterol contents were also analyzed using the same kits as those used for blood analysis (Asan Pharmaceutical Co.).
RNA isolation and real-time polymerase chain reaction
Individual tissue samples (50–100 mg) were homogenized with 5 mm stainless steel beads (Qiagen, USA). Then, total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, USA). Purified RNA (2 μg) was reverse transcribed with PrimeScript™ RT Master Mix (Takara Bio, Inc., Japan) and then cDNA was synthesized. The cDNA was amplified on a StepOne™ Real-Time PCR System (Applied Biosystems, USA) using the SYBR® Green PCR Master Mix (Applied Biosystems) and TaqMan® Fast Advanced Master Mix (Applied Biosystems). The mouse primer sequences (HMGCR, LDLR, SREBP2, Occludin, and GAPDH) are described in Table 2. Inventoried TaqMan primer sets (Applied Biosystems) were used for analysis of TNF-α (Mm00443258_m1), IL-1β (Mm01336189_m1), MCP1 (Mm00441242_m1), CYP7A1 (Mm00484152_m1), and GAPDH (Mm99999915_g1). The expression levels of the mRNAs were analyzed using the 2−ΔΔCt method and were normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as an endogenous control.
Mouse Primers for Reverse Transcription–Polymerase Chain Reaction
GAPDH, glyceraldehyde-3-phosphate dehydrogenase; HMGCR, 3-hydroxy-3-methyl-glutaryl-CoA reductase; LDLR, low-density lipoprotein receptor; RT-PCR, reverse transcription–polymerase chain reaction; SREBP2, sterol regulatory element binding protein 2.
Mucosal bacterial DNA extraction and real-time polymerase chain reaction
Colonic samples were homogenized with 5-mm stainless steel beads (Qiagen). Then, total DNA was extracted using the TRIzol reagent (Invitrogen). The purity and integrity of DNA preparations were checked with a Nanodrop (Thermo Scientific, USA) and agarose gel electrophoresis. Polymerase chain reaction (PCR) was performed on a StepOne Real-Time PCR System (Applied Biosystems) using the SYBR Green PCR Master Mix (Applied Biosystems). The bacterial primer sequences (L. plantarum and eubacteria) are described in Table 2. The mucosal expression levels of L. plantarum DNA were analyzed using the 2−ΔΔCt method and were normalized to the expression levels of eubacteria as an endogenous control.
Statistical analysis
Statistical analyses were performed using SPSS software (ver. 22.0; IBM Corp., USA). Data were expressed as mean ± standard error of the mean. Differences between the groups were assessed by one-way ANOVA (analysis variance) followed by Duncan's multiple range test. The statistical significance was set at P < .05.
Results
Body weight gain, organ weight, and feed intake
The body weight gain and relative organ weight of the experimental groups are shown in Table 3. After the 16-week feeding period, the HFD group showed significantly greater body weight gain and relative fat weight than the ND group (P < .05). The PMOChia group showed lower body weight gain (by 10%) and relative fat weight (by 14%) than the HFD group, and the other experimental diet groups showed similar body weight gain and relative fat weight to the HFD group. Also, the relative epididymal fat and liver weight were not significantly different among the groups consuming HFD. There were no significant differences in food intake among the experimental group over the 16-week feeding period (data not shown).
Effects of Experimental Diets on Body Weight Gain, Relative Liver Weight, and Relative Fat Weight in High-Fat Diet-Fed Mice
Data are presented as the mean ± SEM (n = 10).
Relative liver weight (%) = 100% × liver weight (g)/body weight (g).
Relative fat weight (%) = 100% × fat weight (g)/body weight (g).
Different letters within a variable are significantly different at P < .05.
SEM, standard error of the mean.
Serum metabolic parameters, the AI, and the cardiac risk factor
Table 4 shows serum lipid composition. The serum total cholesterol and LDL cholesterol levels in the HFD group were significantly higher than in the ND group (P < .05), suggesting that HFD feeding led to the onset of dyslipidemia. The PMO and Chia groups showed significantly lower plasma LDL cholesterol levels compared to the HFD control group (P < .05). Moreover, total cholesterol levels were significantly lower in the PMO group compared to the HFD group (by 21%). Also, cardiac risk factor and the AI were significantly lower in the PMO group compared to the HFD group (Table 5). Fasting glucose and insulin concentrations in the experimental groups are presented in Table 6. Fasting blood glucose levels were increased in the HFD group than in the ND group (P < .05). However, no other experimental diet groups showed any positive effects in this experiment. Furthermore, none of the experimental groups showed a significant difference in plasma insulin or insulin resistance (measured using the homeostatic model assessment).
Effects of the Experimental Diets on Plasma Lipid Profiles in High-Fat Diet-Fed Mice
Data are presented as the mean ± SEM (n = 10).
Values with different letters within the same column are significantly different at P < .05 by the Duncan multiple range test.
C, cholesterol; HDL, high-density lipoprotein; LDL, low-density lipoprotein.
Effects of the Experimental Diets on the Atherogenic Index and Cardiac Risk Factor
Data are presented as the mean ± SEM (n = 10).
Values with different letters within the same column are significantly different at P < .05 by the Duncan multiple range test.
Atherogenic index = (total cholesterol-HDL cholesterol)/HDL cholesterol.
Cardiac risk factor = total cholesterol/HDL cholesterol.
Effects of the Experimental Diets on Fasting Glucose, Insulin, and Homeostatic Model Assessment Of Insulin Resistance in High-Fat Diet-Fed Mice
Data are mean ± SEM, n = 10.
Values with different letters within the same column are significantly different at P < .05 by the Duncan multiple range test.
Chia, HFD with chia seeds; HOMA-IR, homeostatic model assessment of insulin resistance; NS, not significantly different among groups; PMO, HFD with L. plantarum PMO 08; PMOChia, HFD with L. plantarum PMO 08+chia seeds; SEM, standard error of the mean.
Liver triglycerides and total cholesterol
Liver triglycerides and cholesterol contents in the experimental groups are shown in Table 7. Compared to the HFD group, liver triglyceride levels in the PMOChia and Chia groups were lower (66% and 55%, respectively). However, no significant differences were found among the different experimental diets. Hepatic total cholesterol was significantly lower in the Chia group than in the HFD group (P < .05).
Effects of the Experimental Diets on Hepatic Lipid Profiles in High-Fat Diet-Fed Mice
Data are presented as the mean ± SEM (n = 7).
Values with different letters within the same column are significantly different at P < .05 by the Duncan multiple range test.
Hepatic and epididymal mRNA levels
Figure 1 shows the expression levels of the genes involved in hepatic lipid metabolism. The level of sterol regulatory element binding protein 2 (SREBP2) mRNA was significantly lower in the PMO group than in the HFD group by 61.5% (P < .05). The PMO group showed 31% and 54% lower levels of 3-hydroxy-3-methyl-glutaryl-CoA reductase (HMGCR) and LDL receptor (LDLR) mRNA than the HFD group. Finally, CYP7A1 (cholesterol 7α-hydroxylase) mRNA levels were, respectively, higher (by 204%) in the PMOChia group compared to the HFD group. Table 8 shows the expression of monocyte chemoattractant protein 1 (MCP1), tumor necrosis factor alpha (TNF-α), and interleukin 1-beta (IL-1β) in epididymal adipose tissue. MCP1 expression in epididymal adipose tissue was significantly higher in the HFD group compared to the ND group, and the Chia group showed the lowest levels of MCP1 expression (P < .05).
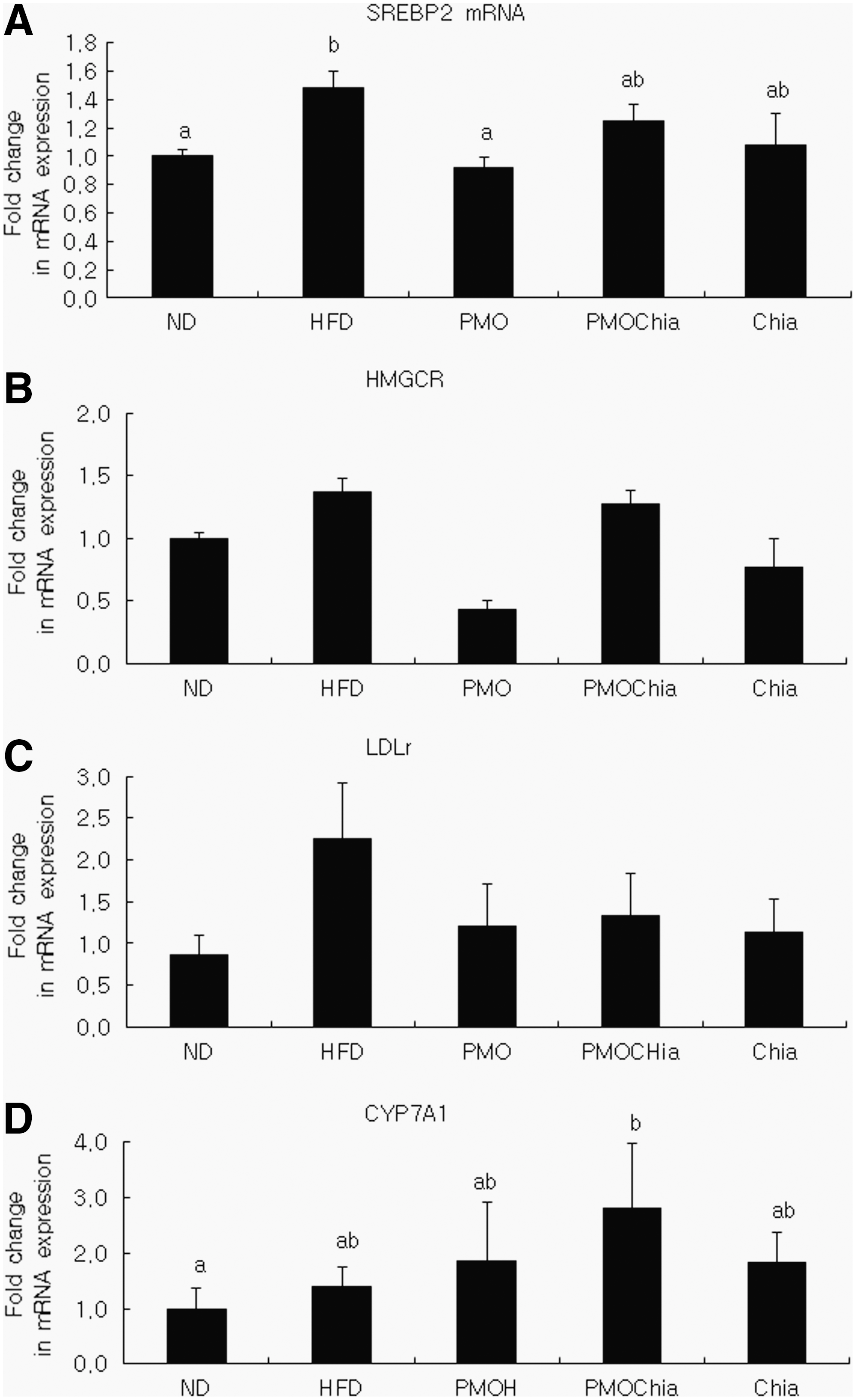
The hepatic mRNA expression levels of SREBP2
Effects of the Experimental Diets on mRNA Expression in High-Fat Diet-Fed Mice
Data are mean ± SEM (n = 6).
Values with different letters within the same column are significantly different at P < .05 by the Duncan multiple range test.
IL, interleukin; MCP, monocyte chemoattractant protein; TNF, tumor necrosis factor.
Tight junction protein mRNA levels in colonic tissue
Table 9 shows the mRNA expression of tight junction protein occludin. An HFD reduced the expression of the tight junction protein occludin by 55% (P < .05), but the PMOChia group showed a significant tendency to resist this reduction, with a 200.5% higher expression than the HFD group.
Effects of the Experimental Diets on Occludin mRNA Expression in High-Fat Diet-Fed Mice
Data are mean ± SEM (n = 6).
Values with different letters within the same column are significantly different at P < .05 by the Duncan multiple range test.
Mucosal L. plantarum DNA levels using real-time-PCR
The expression level of L. plantarum in colonic tissue is shown in Figure 2. The lactic acid concentration in colonic tissues was expressed as the ratio of L. plantarum to total bacteria (eubacteria) normalized to that of the normal dietary group, which was set to 1 (ND = 1). As shown in Figure 2, the proportion of L. plantarum was distinctly reduced in the HFD group by 0.24-fold compared to the ND group. When compared with the HFD group, the PMO and PMOChia groups showed a higher proportion of L. plantarum (208% and 448%, respectively). However, there was no significant difference in the L. plantarum expression levels among all experimental groups.

The relative amount of colonic L. plantarum in mice fed the experimental diets for 16 weeks. Bacterial DNA was amplified by qPCR using 16S rRNA of L. plantarum and eubacteria (all bacteria; housekeeping gene). Results are presented as relative expression to total bacteria (eubacteria), and the relative expression to eubacteria in the ND was set to 1. All values are mean ± SEM. Chia, high-fat diet with 4% chia seed powder; HFD, high-fat diet (45% kcal fat); PMO, high-fat diet with L. plantarum PMO 08 (1.0 × 109 CFU/mice); PMOChia, HFD with L. plantarum PMO 08 + 4% chia seed powder; qPCR, quantitative polymerase chain reaction.
Discussion
Recently, many studies have suggested that the gut microbiota play an important role in metabolic syndrome. 1,2 Thus, therapeutic probiotics that can influence gut microbiota may also modulate some risk factors that lead to metabolic syndrome. However, because the effects of probiotics are strain dependent, some probiotics have been found to modulate obesity levels in mice on HFDs, while other probiotic strains had little effect or promoted weight gain. 11 –15,35,36 The aim of this study was to evaluate whether diets that include L. plantarum PMO 08 alone and L. plantarum PMO 08 combined with chia seeds exerted beneficial metabolic effects in obese mice on HFD. The mice in the HFD control group exhibited higher total cholesterol concentrations, body weight gain, and fasting blood glucose, indicating that they were at a high risk of developing metabolic syndrome. In contrast, the PMO group showed significantly lower serum total cholesterol and LDL cholesterol concentrations compared to the HFD group. In addition, numbers of both the AI (an indicator of the severity of atherosclerosis), 37 and the ratio of LDL cholesterol to high-density lipoprotein cholesterol (a valuable tool for evaluating cardiovascular disease risk), 38 were lower in the PMO group. These all suggest that L. plantarum PMO 08 exerts a hypolipidemic effect and could be beneficial for preventing metabolic syndrome. It is reported that some probiotics have cholesterol-lowering effect, 39 but the mechanism remains uncertain. To elucidate the mechanism of how PMO 08 ameliorates dyslipidemia, we screened the altered expression of genes relevant for lipid metabolism and biosynthesis. Cholesterol homeostasis is mainly regulated by endogenous synthesis, intestinal absorption, and hepatic conversion and excretion. 40 SREBP2 is a transcription factor of target mRNAs for cholesterol-related genes, 41 and one of its downstream targets is HMGCR, which mediates the rate-limiting step of cholesterol synthesis and is the inhibition target of anticholesterol drugs. In previous studies, hepatic SREBP2 mRNA levels were decreased in an HFD group consuming probiotics compared to the control group that did not consume probiotics. This mechanism was also likely responsible for the lower total cholesterol and LDL cholesterol concentrations seen in that group. 40 This is consistent with our results for L. plantarum PMO 08, which significantly lowered serum total cholesterol and LDL cholesterol and downregulated hepatic SREBP2 mRNA expression. Some reports have suggested that probiotics could regulate cholesterol synthesis in the liver by inhibiting HMGCR, 42,43 but, although L. plantarum PMO 08 consumption tended to reduce HMGCR levels, this trend was not significant in this study. Hence, the mechanisms underlying this finding need further elucidation. Probiotic effects on cholesterol levels are primarily due to impacts on bile metabolism, through which they lower gastrointestinal cholesterol absorption and increase cholesterol excretion. 44 Cholesterol can be converted into bile salts by CYP7A1, the initial and rate-limiting enzyme for bile acid synthesis. In previous studies, L. plantarum KCTC3928 significantly lowered plasma LDL cholesterol and accelerated fecal bile acid excretion through upregulation of CYP7A1. 45 Lower levels of bile acids in hepatocytes liberate CYP7A1 from feedback inhibition and increase the removal of both hepatic cholesterol and plasma cholesterol. Our study showed that the significant increase in CYP7A1 mRNA expression triggered by L. plantarum PMO 08 combined with chia seeds could contribute to these changes in plasma and hepatic cholesterol levels. However, L. plantarum PMO 08 showed a tendency to upregulate CYP7A1 mRNA expression, which is thought to be associated with a significant reduction of plasma and hepatic cholesterol levels.
Recent investigations indicate that HFDs affect the composition of gut microbiota, which, in turn, influence gut integrity and cause a state of chronic low-grade inflammation. 46 Some gut microbiota release endotoxins, which can combine in the gut with chylomicrons or can leak through the gut wall. 47 Evidence strongly suggests that chronic low-grade inflammation is a significant factor underlying chronic disorders such as metabolic syndrome. HFDs decrease the number of tight junction protein in the gut, causing greater gut permeability, and increase leakage of lipopolysaccharide into the blood circulation. 48 On the contrary, probiotic bacteria appear to ameliorate this effect and protect against systemic inflammation and metabolic syndrome. 48 To evaluate the disruption of gut barrier integrity, the expression of mRNA coding for the tight junction protein occludin was measured by reverse transcription-PCR. Among the experimental groups, the PMOChia group showed the highest level of occludin mRNA expression, together with the largest proportion of L. plantarum in the colonic mucosa. Conversely, the lowest level of occludin mRNA expression and the smallest proportion of colonic mucosal L. plantarum were observed in the HFD group, indicating a positive correlation between intestinal L. plantarum and gut barrier function. Cani et al. demonstrated that obese mice on HFD had significantly lower numbers of bifidobacteria and higher endotoxin levels compared to mice fed an ND. 5 Subsequent administration of oligofructose as a prebiotic restored quantities of bifidobacteria, reduced endotoxin levels, and lowered levels of proinflammatory cytokines. 5 Similarly, it can be suggested that dietary fiber contained in chia seeds was metabolized by L. plantarum to inhibit the degradation of tight junction protein occludin and increase the relative abundance of lactobacilli. However, this effect has not been confirmed and the research gap is yet to be filled. Unfortunately, in our study, no significant changes in the expressions of HMGCR, LDLR, and CYP7A1 were observed. One possible reason for this is the low quantity of administered probiotics relative to the total gut microbiome, which may have been insufficient to shift the major sets of bacteria that play a role in colonic health. Furthermore, other studies have shown that ingestion of probiotics for 22 weeks improved inflammation and lipid metabolism by promoting SREBP2 mRNA expression, but this effect was not observed in an 11-week intake period. 6 Such observations suggest that sufficient time and dosage are needed to alter the intestinal microbial population to alleviate metabolic disease. It was reported that different kinds of probiotic mixture were more effective than each of their constituent single strains due to synergistic effects on host health. 49 Therefore, it can be inferred that the complementary effects of a combination of probiotic strains on gut microbiota and host health could alleviate different symptoms of metabolic syndrome, thereby generating optimal synergistic efficacy. In this study, the host inflammation, lipid metabolism in the liver and plasma, and gut barrier function were affected differently by the probiotics and chia seeds. As a result, we hypothesize that synbiotics consisting of L. plantarum PMO 08 and chia seeds would be more effective than probiotics with only L. plantarum PMO 08. In the future, it would be intriguing to compare the effects of combinations of synbiotics on metabolic syndrome to determine the optimum efficacy based on their complementary impacts on gut microbiota, as well as to identify the optimal dosage.
In conclusion, this study demonstrated that supplementation with L. plantarum PMO 08 could ameliorate metabolic syndrome by attenuating the plasma total cholesterol and LDL cholesterol concentrations in obese mice fed on HFD. These changes were partly associated with the downregulation of mRNA coding for SREBP2, a transcription factor involved in the regulation of cholesterol metabolism. Furthermore, this experiment demonstrated for the first time that chia seeds acted as a potent source of prebiotics, promoting the growth of healthy and anti-inflammatory intestinal microbiota and fortifying the gut barrier function. Our results suggest that synbiotics may be more effective than probiotics alone in terms of adhesion of intestinal bacteria and gut barrier function. In mice at least, these findings suggest that L. plantarum PMO 08 is a potent antiatherosclerotic agent, that chia seed supplementation might modulate gut microbiota, and that L. plantarum PMO 08 combined with chia seeds may offer a natural alternative to combat metabolic syndrome. However, more in-depth studies proving the cholesterol-lowering effect of L. plantarum PMO 08 and its synergistic effect with chia seeds are required for such a product to be launched in the functional food market.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by Pulmuone Co., Ltd.