
Abstract
Naringin and its aglycone, naringenin, occur naturally in our regular diet and traditional Chinese medicines. This study aimed to detect an effective therapeutic approach for cough variant asthma (CVA) through evaluating the relaxant effect of these two bioactive herbal monomers as antitussive and antiasthmatic on rat tracheal smooth muscle. The relaxant effect was determined by measuring muscular tension with a mechanical recording system in rat tracheal rings. Cytosolic Ca2+ concentration was measured using a confocal imaging system in primary cultured tracheal smooth muscle cells. In rat tracheal rings, addition of both naringin and naringenin could concentration dependently relax carbachol (CCh)-evoked tonic contraction. This epithelium-independent relaxation could be suppressed by BaCl2, tetraethylammonium, and iberiotoxin (IbTX), but not by glibenclamide. After stimulating primary cultured tracheal smooth muscle cells by CCh or high KCl, the intracellular Ca2+ increase could be inhibited by both naringin and naringenin, respectively. This reaction was also suppressed by IbTX. These results demonstrate that both naringin and naringenin can relax tracheal smooth muscle through opening big conductance Ca2+-activated K+ channel, which mediates plasma membrane hyperpolarization and reduces Ca2+ influx. Our data indicate a potentially effective therapeutic approach of naringin and naringenin for CVA.
Introduction
Cough variant asthma (CVA) is considered to be a clinical variant form of asthma with an intensely irritating dry cough as the predominant presenting symptom. 1 –3 Although having no sign of wheezing, shortness of breath, or other symptoms compared with asthma patients, CVA patients exhibit airway hyperresponsiveness and show excellent response to bronchodilators, which relax airway smooth muscle. 4 –8 CVA is associated with sputum eosinophilia and eosinophilic airway inflammation. 4 The mechanisms underlying cough and bronchoconstriction of CVA have been suggested as differences in location or degree of activation of eosinophils and allergic cytokines. 3 CVA is treated in a similar manner as classic asthma. 2 The patient is generally given an asthma inhaler with albuterol, ipratropium, and/or inhaled corticosteroids, 4 which inhibit bronchoconstriction by preventing bronchial smooth muscle contraction, bronchial wall inflammation, and increased mucosal secretion.
Slender spindle-shaped smooth muscle cells distribute in the respiratory tract. Airway diameter and airflow resistance are closely related to the activity of smooth muscle cells. Smooth muscle cells can maintain the stability of respiratory tract volume via resisting the applied load, controlling the size of dead space to assist the lung contraction, exhaling gas and foreign body, adjusting the diameter of airway, and adapt to cough. 9 Intracellular Ca2+ concentration ([Ca2+]i) of smooth muscle cells can affect contraction by regulating the interaction of myosin and actin. A high concentration of Ca2+ activates myosin light chain kinase in contractile smooth muscle cells. The increase of Ca2+ in cytosol mainly depends on the release of stored Ca2+ from the sarcoplasmic reticulum and the transportation of extracellular Ca2+ through ion channels. 10
Bronchodilators targeting airway smooth muscle act as essentially conventional treatments for CVA. Interest in discovering complementarily effective bronchodilators as potential small molecular targets has risen in recent years. Natural products have been reported to be an effective alternative treatment strategy for CVA. Therefore, much attention is focused on the study of natural products.
Naringin (Fig. 1A), a kind of dihydroflavone, widely exists in many Citrus fruits, such as orange (Citrus sinensis), grapefruit (Citrus paradisi), and tangerine (Citrus reticulata). Naringenin (Fig. 1B), the aglycone of naringin, can also be found but to a lesser extent in the regular diet. Previous pharmacokinetic studies of naringin have suggested that naringenin is the most common form in serum after oral administration of naringin because naringin can be metabolized into absorbable naringenin by intestinal flora. 11,12
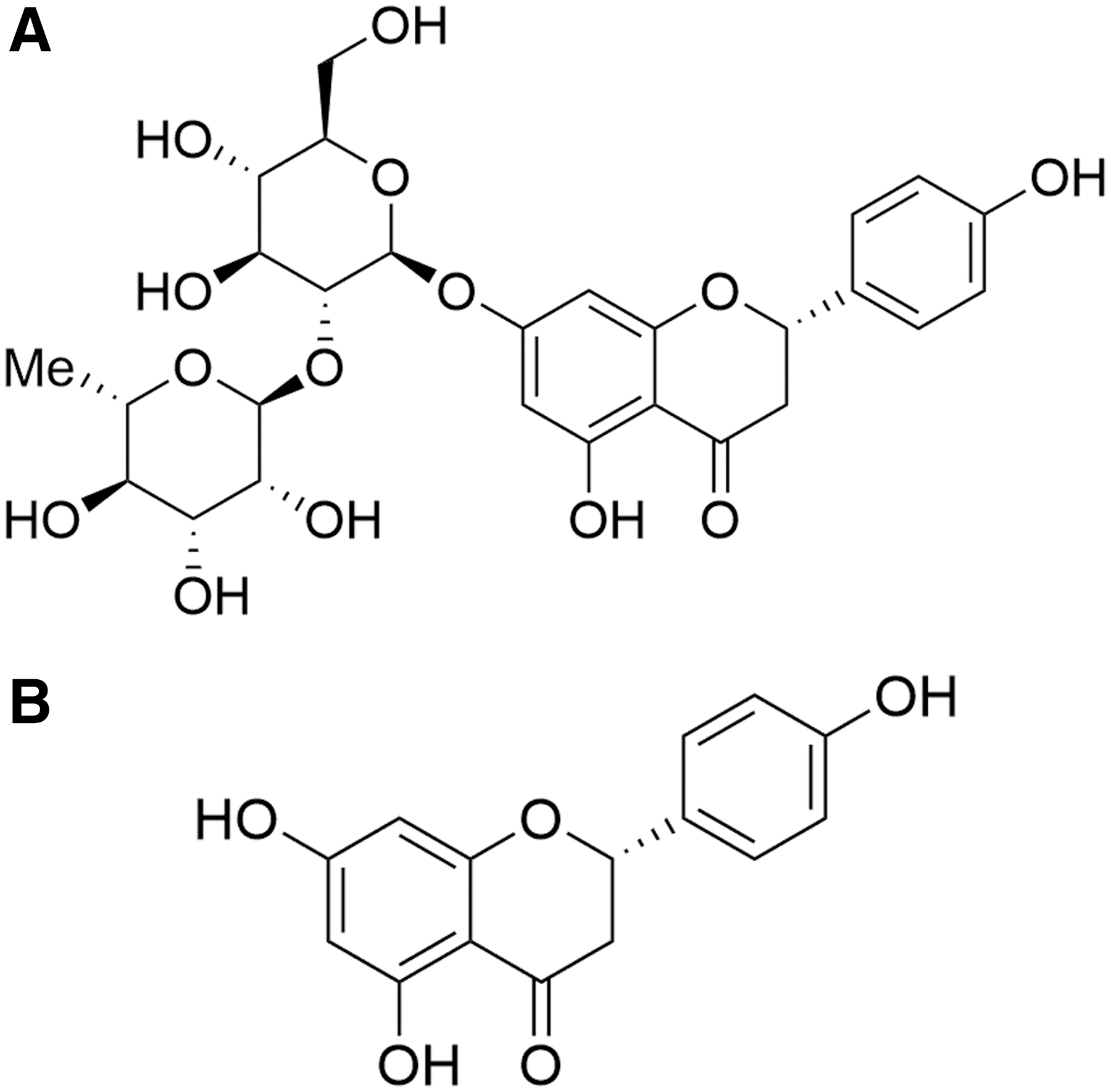
Chemical structure of naringin
Previous studies have indicated that naringin and its major metabolite naringenin exhibit antioxidative, anti-inflammatory, antitussive, antiasthmatic, and expectorant effects both in vitro and in vivo. 13 –22 Among them, it is worth noting that naringin and naringenin remarkably attenuate the airway hyperresponsiveness and cough reflex sensitivity in various animal models of respiratory diseases. 16,23 This evidence suggests that naringin and naringenin may have the ability to relax the airway smooth muscle, resulting in the antitussive effect, but the underlying mechanism remains unclear. Therefore, the current study is designed to investigate the regulatory mechanisms of naringin and naringenin on relaxing airway smooth muscle and evaluate therapeutic potentials for the treatment of CVA.
Materials and Methods
Animal preparation
Sprague–Dawley rats (100–200 g) of either sex were purchased from Guangdong Medical Laboratory Animal Center (Guangzhou, China). Rats were housed under controlled conditions with a 12:12-h light–dark cycle, temperature of 21°C, and 60% humidity, with food and water ad libitum. The experiment was approved by the Animal Care and Use Committee of the School of Life Sciences (Permit No. SLS2012–003, Sun Yat-sen University, P.R. China). Adequate measures were taken to minimize the pain of the animals.
Chemicals
Naringin, prepared by our laboratory, was extracted from Citrus grandis “Tomentosa” by water, and deposited in ethanol, with a concentrated filtrate obtained after a 1:10 recrystallization, purity >98.3%, finally determined by peak area normalization. 24 Naringenin, CCh (carbachol), GLI (glibenclamide), TEA (tetraethylammonium), IbTX (iberiotoxin), and collagenase I were purchased from Sigma-Aldrich (St. Louis, MO, USA). All of the above reagents were dissolved in DMSO (dimethyl sulfoxide). The final concentration of DMSO was 0.1%, which had no significant effects on contractile tension and intracellular Ca2+ content. Fluo-3 AM was purchased from Molecular Probes. NaCl, KCl, KH2PO4, NaHCO3, MgSO4, CaCl2, BaCl2, and glucose were from Guangzhou Chemical Pharmaceutical Factory (Guangzhou, China). The purity of all chemical reagents was at least analytical grade.
Contractile tension measurements
Rats were sacrificed by CO2 asphyxiation, and tracheal tissues were dissected out and put into Krebs-Henseleit Solution (K-H solution, containing 117 mM NaCl, 4.7 mM KCl, 1.2 mM KH2PO4, 25 mM NaHCO3, 1.2 mM MgSO4, 2.56 mM CaCl2, and 11.1 mM glucose, finally adjusted to pH 7.4), ventilated in 95% O2 and 5% CO2 at 37°C. To avoid tissue damage, the hard tearing was displaced by blunt separation. The trachea was cut into 2–3 mm tracheal rings under dissecting microscopy after removal of blood vessels and connective tissues. The epithelium was removed by gentle mechanical friction to minimize damage to the smooth muscle.
After dissection, the portions of trachea were mounted with thin wire threaded onto a hook, attached to a tension transducer and placed in a thermostatic perfusion system containing 10 mL K-H solution of ventilated in 95% O2 and 5% CO2 at 37°C. Ten millinewton preload tension was applied before each experiment and equilibrated for 30 min. After the application of 200 nM CCh caused a tonic contractile response for 10–15 min, naringin or naringenin was added. Different channel blockers were used to investigate the involvement of ion channels in the relaxant action of naringin and naringenin. Tracheal ring tension was recorded by a mechanical recording system (BL-420E+; Chengdu Taimeng Technology, Chengdu, China).
Cell culture
Rat tracheal smooth muscle cells were cultured as described previously, with some modifications. 25 In brief, acutely dissociated rat tracheas were enzymatically digested for 50 min at 37°C by 0.1% collagenase I after mincing for 30 min. Isolated cells were harvested and cultured in Dulbecco's modified Eagle's medium/F12 (DMEM/F12; Gibco, USA) with 10% fetal bovine serum (HyClone, Australia) and 100 U/mL penicillin–streptomycin solution (Gibco), in a humidified 5% CO2 37°C incubator.
Immunofluorescence
Primary cultured rat tracheal smooth muscle cells grown on coverslips were incubated with DMEM/F12 for 3 days. After being fixed in 4% paraformaldehyde for 15 min at room temperature, cells were permeated with 0.3% Triton and 5% bovine serum albumin mixed solution for 1 h. The cells were incubated with anti-α-smooth muscle actin (α-SMA) antibody (BOSTER, Wuhan, China) or keratin (27/28) antibody (Santa Cruz Biotechnology, Dallas, USA) for 2 h at room temperature. After washing with phosphate-buffered saline (HyClone, China) for three times, cells were, respectively, incubated with an anti-mouse IgG Cy3-conjugated antibody (120087; Jackson ImmunoResearch, USA) to mark α-SMA or an anti-goat IgG FITC-conjugated antibody (E031230-01; EARTHOX, USA) to mark keratin. Nuclei were stained with DAPI (Cell Signaling Technology, USA). Fluorescence was observed using a fluorescence microscope (Olympus IX83).
Measurement of [Ca2+]i
[Ca2+]i was measured by fluorescence intensity assay. After culturing the rat tracheal smooth muscle cells on coverslips for 3 days, cells were washed by normal physiological saline solution (contained 140 mM NaCl, 5 mM KCl, 1 mM MgCl2, 2.5 mM CaCl2, 10 mM HEPES, and 10 mM glucose, finally adjusted to pH 7.4) for three times and incubated with 5 μM fluo-3 AM for 60 min at room temperature. Fluorescence signal was recorded by the laser scanning confocal imaging system (TCS-SP5; Leica Microsystems, Germany). Fluo-3 AM was excited at 488 nm and analyzed at 530 nm. After equilibrating for 3–5 min, cells were, respectively, treated with KCl and CCh to increase [Ca2+]i. Subsequently, naringin or naringenin was added with the presence or absence of relevant inhibitors. The change of fluorescence intensity was normalized to the initial intensity.
Data analysis and statistics
The data are represented as the mean ± standard error of the mean (n is the number of tissue preparations or experiment replications). Student's t test was used to evaluate statistical significance. A value of P < .05 was considered statistically significant.
Results
To test whether naringin and naringenin have the ability to regulate airway tone, acutely isolated rat airway tissue was subjected to contractile tension tests. The application of 200 nM CCh caused a tonic contractile response. After 10–15 min, tension remained stable. Naringin (1 mM) or naringenin (1 mM) was added and caused a significant relaxation response (Fig. 2A, B). The relaxation was concentration-dependent (Fig. 2C), with an EC50 of ∼0.71 mM (naringin) and 0.89 mM (naringenin).

Relaxant effect of naringin and naringenin on rat tracheal rings. Addition of naringin (1 mM)
The epithelium-derived relaxing factors released by airway epithelial cells can relax the underlying smooth muscle. 26 To determine whether the epithelium is implicated in naringin- and naringenin-induced relaxation, the epithelium was mechanically removed by gently rubbing the lumen. The results showed that both naringin and naringenin exhibited a similar relaxant effect on epithelium-intact and epithelium-denuded rat tracheal rings (Fig. 3), suggesting that the naringin- and naringenin-induced relaxations are epithelium-independent.
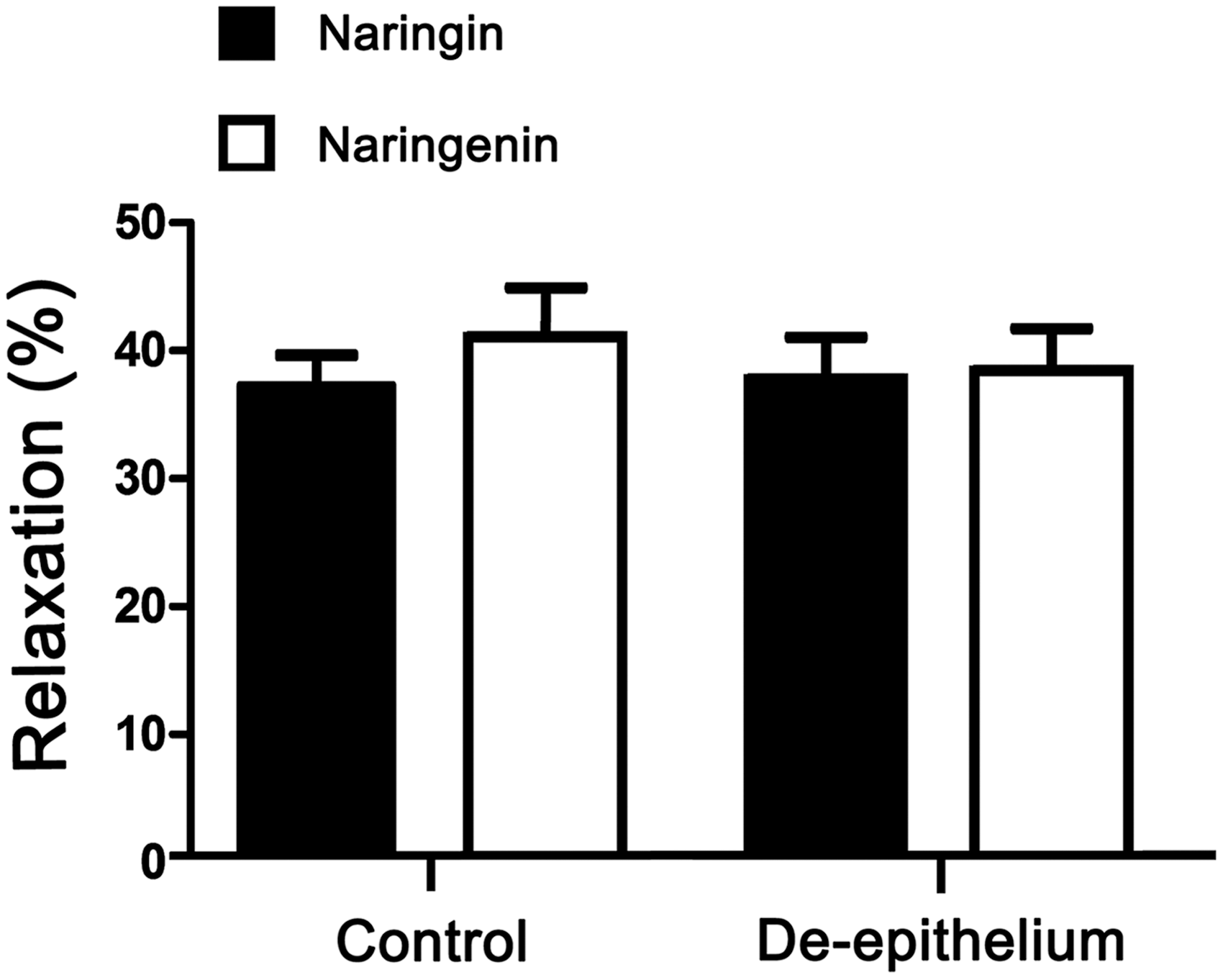
Effect of airway epithelium on naringin- and naringenin-evoked relaxation response in rat tracheal rings. There was no significant difference. Data are presented as the mean ± SEM (n = 5).
To investigate the involvement of ion channels in the relaxation effect of naringin and naringenin, different K+ channel blockers (Fig. 4) were used. Pretreatment of non-specific K+ channel blocker BaCl2 (500 μM) almost abolished the relaxant effect of naringin and naringenin. While adenosine triphosphate (ATP)-sensitive K+ channel blocker GLI (10 μM) failed to attenuate the naringin- and naringenin-induced relaxation, voltage-gated and Ca2+-activated K+ channel blocker TEA (10 μM) and big conductance Ca2+-activated K+ channel (BKCa)-specific blocker IbTX (100 nM) strongly inhibited the response. Since the activation of BKCa usually results in an influx of K+, the above results hinted that the epithelium-independent relaxation of smooth muscle induced by naringin and naringenin could be attributed to the BKCa-dependent membrane hyperpolarization.

Effect of different K+ channel blockers on the naringin- and naringenin-evoked relaxation in rat tracheal rings. Pretreated tissues with BaCl2 (500 μM), TEA (10 μM), and IbTX (100 nM), naringin (1 mM) could not induce the relaxation effect, except for GLI (10 μM) pretreatment
It is well known that K+ channel-mediated smooth muscle membrane hyperpolarization can reduce [Ca2+]i and induce a relaxation response in the tracheal ring. 10,27,28 To investigate whether naringin and naringenin could reduce the [Ca2+]i, [Ca2+]i was measured by real-time laser scanning confocal imaging system in primary cultured tracheal smooth muscle cells.
First, the primary cultured cells were authenticated as smooth muscle cells using specific marker α-SMA (Fig. 5). [Ca2+]i, indicated by fluo-3 fluorescence intensity, promptly increased by the application of CCh (200 nM) and then attenuated by naringin (100 μM) and naringenin (100 μM), respectively (Fig. 6). The same patterns of [Ca2+]i changes were observed when the smooth muscle cells were depolarized by high concentrations of KCl (30 mM) (Fig. 7). Decreases of [Ca2+]i induced by naringin (100 μM) and naringenin (100 μM) were inhibited by 100 nM IbTX (Fig. 7). The above results suggested that both naringin and naringenin can attenuate the [Ca2+]i increase induced by CCh and KCl, to exercise relaxation effects via activation of BKCa.

Marking primary cultured cells with anti-α-SMA antibody

Effect of naringin and naringenin on CCh-evoked [Ca2+]i increase in primary cultured tracheal smooth muscle cells. Naringin and naringenin, respectively, reduced CCh-elicited [Ca2+]i increase on average time-course traces
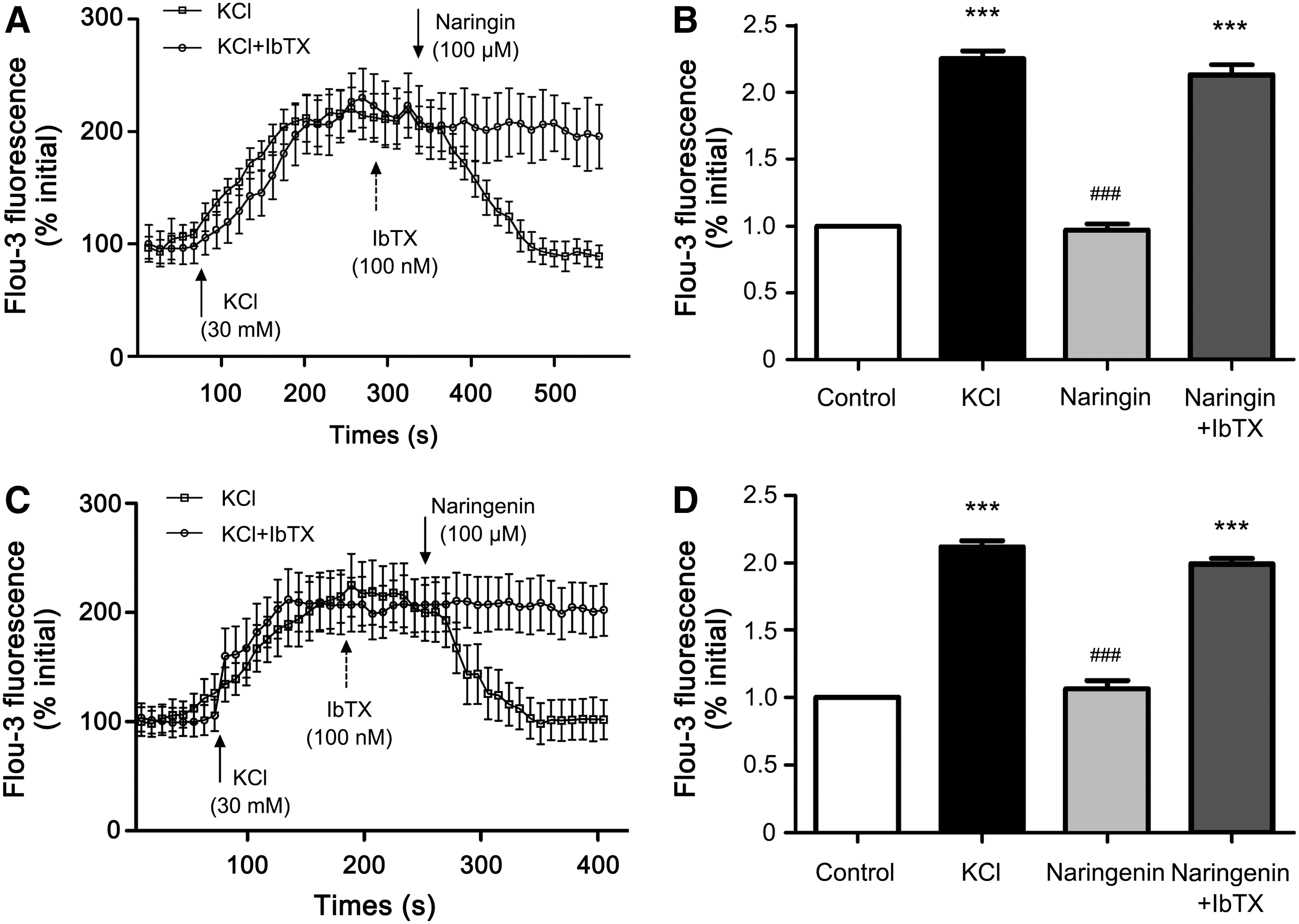
Effect of naringin and naringenin on KCl-evoked [Ca2+]i increase in primary cultured tracheal smooth muscle cells. Naringin
Discussion
CVA is a type of asthma lasting at least 6–8 weeks. The main symptom of CVA is dry, nonproductive cough. Previous studies have indicated that CVA is one of the most common causes of coughing worldwide. 29 Without adequate treatment, 30–40% CVA patients may progress to classic asthma. 30 Their response to bronchodilators indicates that the cough of CVA may be due to bronchoconstriction. Therefore, the drug with relaxation effect on airway smooth muscle has a therapeutic effect on CVA to certain degree. 4,7
As a potential pharmaceutical target for regulating bronchial relaxation during bronchoconstriction, 31 BKCa hyperpolarizes the airway smooth muscle with large outward K+ current. 32,33 Therefore, activation of BKCa is the most common treatments for bronchoconstriction. The present study initially demonstrated that naringin and naringenin have the ability to trigger IbTX-sensitive hyperpolarization via BKCa activation in airway smooth muscle tissue and regulate [Ca2+]i in primary cultured rat tracheal smooth muscle cells, ultimately producing relaxation effect.
As plant-based small molecules, naringin and naringenin have been reported to relieve cough and treat CVA in respiratory diseases. 16,19,23 Naringin can reduce airway inflammation in an ovalbumin-induced guinea pig CVA model. 23 Therefore, naringin and naringenin can play a certain therapeutic effect on CVA diseases from the perspective of anti-inflammatory, but their regulation and mechanism of airway smooth muscle tension contraction are still unclear. This study aimed to investigate the relaxation mechanism of naringin and naringenin and discuss their potential as a bronchodilator in the treatment of CVA diseases.
Growing evidence suggests that naringin is a peripheral antitussive by inhibiting responses of rapidly adapting receptors, but without function of the sensory neuropeptides system or the modulation of ATP-sensitive K+ channels. 19 Our previous study demonstrated that naringenin was able to produce BKCa-dependent relaxant effect on rat colonic smooth muscle by hyperpolarizing the membrane and decreasing Ca2+ influx. 34 Similarly, naringenin concentration dependently induced activation of BKCa in vascular smooth muscle cells and mediated relaxation, so did naringin, but with a lower vasorelaxant effect. 35 Furthermore, naringenin has the ability to increase BKCa activity in HEK293T cells. 36 These results suggest that naringin and naringenin may regulate smooth muscle constriction by BKCa-mediated hyperpolarization in rat airway.
In the present study, we found that both naringin and naringenin could dose dependently stimulate a relaxation response in CCh-induced contraction (Fig. 2). Compared with the relaxation response elicited by naringin, naringenin-induced dose–response curve was much more significant at the large amount, indicating that naringenin generates a stronger relaxation effect than naringin. This may be attributed to naringenin's lower polarity and smaller molecular weight, which make it easier to pass through the cell membrane.
It is well known that the relaxation of smooth muscle cells is divided into epithelium-dependent and nonepithelial-dependent types. Previous studies have demonstrated that airway epithelium can release PGE2, NO, and other epithelial relaxing factors to remit the contraction of smooth muscle. 26,37 In our study, the data (Fig. 3) revealed that the relaxation effects of naringin and naringenin are largely attributed to direct stimulation on the airway smooth muscle rather than a secondary response mediated by the airway epithelium.
[Ca2+]i of smooth muscle cells can affect contraction by regulating the interaction of myosin and actin. 38 The increase of Ca2+ in cytosol mainly depends on the release of stored Ca2+ from sarcoplasmic reticulum and transport the extracellular Ca2+ through ion channels. 10 In nonepithelial-dependent relaxation, the mechanisms of removing intracellular Ca2+ mainly involve the activation of potassium channels to hyperpolarize the cell membrane, and the antagonism of calcium channels to prevent Ca2+ influx. 10,39 Relaxation mechanisms caused by activation of potassium channel include hindering the voltage-gated calcium channel opening by membrane hyperpolarization, activating the Na+-Ca2+ exchanger by Na+-K+ pump excitation, regulating and affecting the release of intracellular Ca2+. 31
K+ channels have a great influence on the electrical stabilities of smooth muscle cells. It mainly includes large-conductance K+ channel and ATP-sensitive K+ channel. The large-conductance K+ channel is activated by intracellular cAMP, cGMP, increase of Ca2+ concentration, and inhibited by IbTX. 31,40 The ATP-sensitive K+ channel is activated by decreases in intracellular ATP, and inhibited by GLI. 41,42 K+ channel blockers were used to screen K+ channels which were activated by naringin and naringenin, the observation showed that nonspecific K+ channel blocker BaCl2, voltage-gated, and Ca2+-activated K+ channel blocker TEA and BKCa-specific blocker IbTX could interdict the naringin- and naringenin-induced relaxation effect, while ATP-sensitive K+ channel blocker GLI had no effect (Fig. 4). These results demonstrated that both naringin and naringenin have the ability to induce a relaxation response by stimulating BKCa-mediated K+-induced hyperpolarization on airway smooth muscle.
Previous studies have shown that the [Ca2+]i of smooth muscle cells acts as a regulatory signal for contraction. 38,43,44 Thus, it is important to explore whether naringin and naringenin could regulate [Ca2+]i in primary cultured rat tracheal smooth muscle cells. Fluorescence intensity assay showed that the increase of [Ca2+]i caused by CCh and high concentration of KCl could be, respectively, reduced by naringin and naringenin (Figs. 6 and 7). Furthermore, the reconversions of naringin and naringenin were abrogated by IbTX, the BKCa channel inhibiter (Fig. 7). These results were in good agreement with the result of the contractile tension test. The experiment results suggest that the mechanism of naringin- and naringenin-induced relaxation shares a common regulatory pathway by activating BKCa-induced hyperpolarization and reducing [Ca2+]i.
In conclusion, this study indicated that naringin and naringenin both exert a potent relaxation effect on tension in the CCh-induced rat tracheal ring constriction model. By mediating the activation of BKCa, naringin and naringenin reduce the [Ca2+]i to induce relaxation of airway smooth muscle. The results indicate that the mechanism of naringin and naringenin provides a potential treatment strategy for CVA.
Footnotes
Acknowledgments
The work was supported by the National Major Scientific and Technical Special Project of China (No. 2015ZX09101014) and the Applied Science and Technology R&D Special Fund Project of Guangdong Province (No. 2015B020234004).
Author Disclosure Statement
No competing financial interests exist.