
Abstract
Obesity is caused by an energy imbalance between food intake and energy expenditure, and has detrimental effects on human health. Platycodon (Platycodon grandiflorum) widely grows in Korea, Japan, and China. It has long been used for food and as a medicinal product. However, the mechanism of the improvement of obesity by platycodon was still not clear. Therefore, we investigated the detailed mechanisms of the antiobesity activity of platycodon extracts. Twenty mice (C57BL/6J) were placed into five groups. The test group received 1 g/kg platycodon extracts. The positive control group received 10 mg/kg orlistat, while the negative control and normal control groups received phosphate-buffered saline. The extracts were given orally daily for 8 weeks. The in vivo treatment of platycodon extracts reduced body weight gain by 7.5%, improved plasma lipid profiles. In the groups given platycodon extracts, leptin was significantly decreased whereas adiponectin was increased. Furthermore, platycodon extracts downregulated lipogenic gene (e.g., lipoprotein lipase, acetyl-CoA carboxylase, and fatty acid synthase) expression and increased lipolysis genes (e.g., carnitine palmitoyltransferase 1α, hormone-sensitive lipase, and uncoupling proteins 2) in liver and white adipose tissue. In addition, platycodon extracts inhibited the expression of key adipogenic transcriptional factors. In conclusion, we have demonstrated that platycodon extracts ameliorate high-fat diet-induced obesity and its related metabolic disease by regulating multiple pathways. Dietary supplementation of platycodon extracts as a functional food and medicinal ingredients may be suitable for prevention and treatment of obesity.
Introduction
Recently, in modern society, obesity is rapidly increasing in the population due to westernization of dietary life, changes in living environment, and excessive nutrition due to increased processed food and decreased physical activity. 1 Obesity is caused by an energy imbalance between calorie intake and energy excess, in which the remaining energy is stored as body fat, which is caused by the accumulation of intracellular triglycerides (TGs) in adipocytes by adipogenesis process. 2 The adipogenesis process requires the activation of transcription factors such as peroxisome proliferation-activated receptor gamma (PPARγ), CCAAT/enhancer-binding protein (C/EBP) and sterol regulatory element-binding proteins (SREBP). 3 –5 Furthermore, obesity is known to be a cause of various diseases such as diabetes, hyperlipidemia, cardiovascular disease, and arteriosclerosis, and it has become a serious disease worldwide. 6 Strategies for the treatment of obesity include the regulation of fat and protein metabolism or blocking the storage of fat and protein, decreased food intake, increased fat decomposition, increased thermogenesis, and inhibition of nutrient uptake. 7 –11 To treat obesity, obesity drugs developed so far include orlistat, mazindol, and phentermine to suppress fat absorption and suppress appetite. However, these drugs have adverse effects such as fat and gastrointestinal diseases, mouth dryness, and sleeping disorders. Therefore, more effective and safe antiobesity substances are required from natural materials. 12 –14
Platycodon (Platycodon grandiflorum) widely grows in Korea, Japan, and China. It is an alkaline food which is abundant in minerals, including K, Mg, Ca, Na, Zn, and Fe, and fiber. 15 It has long been used for food and a medicinal product. Platycodin A, C, D, and polygalacin D, which are triterpenoid saponins, are known to be the main pharmacological components of the platycodon and contain sterol components, including stigmasterol. 16 These saponins have been reported to have various effects such as antiobesity, improvements in blood glucose and cholesterol metabolism, anticancer, anti-inflammation, and alleviation of atopic dermatitis. 17 –20 However, the mechanism involved in the improvement of obesity by platycodon are still not clear. Thus, the aim of this study was to investigate the effect of platycodon on lipogenesis and lipolysis in high-fat diet (HFD)-induced obese mice.
Materials and Methods
Sample preparation
The whole platycodon plants were washed and then were extracted using distilled water at 120°C for 4 h, followed by filtering of the extract. The extracts were concentrated using a rotatory evaporator and then finally freeze-dried to obtain extract powder.
Animals and treatments
All animal experiment procedures were in accordance with the Animal Experiments Guidelines of the National Institute of Agricultural Sciences and were approved by the Institutional Animal Care and Use Committees (IACUC) of the National Institute of Agricultural Sciences (NAS201701). The mice were maintained at a temperature of 23 ± 3°C and a humidity of 40–60% in a light-controlled (12 h light/dark cycles) cages.
Male C57BL/6J mice (8 weeks old) were obtained from the Central Laboratory. Animal, Inc. (Seoul, Korea). The mice were kept for 1 week to acclimatize, and then divided into four groups (five mice per group) as follows. (1) Normal group: mice fed a normal chow diet, (2) HFD group: mice fed HFD, (3) Orlistat group: mice fed HFD plus orlistat (10 mg/kg, positive control), and (4) Platycodon group: mice fed HFD plus platycodon extract (1 g/kg) treatment. The doses of platycodon extract, 1 g/kg per oral (p.o.), were selected based on conducting pilot experiments as follows (dose–effective relationship). We performed the pilot study at 0.25, 0.5, and 1 g/kg concentrations in mice. As a result, there was a tendency to improve mental stress at 0.25 and 0.5 g/kg but not significant, and it was significant at 1 g/kg. Therefore, we used the 1 g/kg concentration in this study (data not shown). The treatment of platycodon extract or orlistat was administered once a day for 8 weeks via oral gavage (p.o). Body weight and food intake were measured weekly.
Plasma and tissue collection
After 8 weeks, all mice were fasted overnight and then sacrificed. Blood was collected from the abdominal aorta with heparinized syringes. To obtain the plasma, the blood sample was centrifuged at 3000 g for 15 min. The liver and white adipose tissues (WATs) were collected and stored at −80°C.
Biochemical analysis
TG, total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C), leptin, and adiponectin in plasma were measured with commercial kits (Abcam, Cambridge, MA, USA) and atherogenic index (AI) and cardiac risk factor (CRF) were calculated as per the following equations: AI = [(TC) – (HDL-C)]/(HDL-C), CRF = (TC)/(HDL-C).
Real-time reverse transcription polymerase chain reaction analysis
Total RNA was isolated from the tissue samples using the RNeasy Mini Plus Kit (Qiagen, Valencia, CA, USA). Complementary DNA was synthesized from isolated RNA (500 ng; Promega, Madison, WI, USA) according to the instructions of the manufacturer. Quantitative polymerase chain reaction (PCR) was carried out using 2X SYBR Green mix (Qiagen). All results were normalized to glyceraldehyde 3-phosphate dehydrogenase expression.
Western blots analysis
Liver and WATs were lysed in lysis buffer containing 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 0.5% NP-40, 0.5% triton X-100, 0.1% Na-deoxycholate, 1 mM EDTA, protease inhibitor, and phosphatease inhibitor. After centrifuging the lysates (12,000 g, 10 min, 4°C), the supernatants were collected. Protein concentration was measured using a Bicinchoninic Acid (BCA) Assay Kit (GenDEPOT). The protein (10 μg) was separated by 4–20% sodium dodecyl sulfate-polyacrylamide gel electrophoresis, transferred to a polyvinylidene fluoride membranes (Bio-Rad, Hercules, CA, USA), and incubated with the appropriate antibody. The resulting bands were detected using enhanced chemiluminescence (ECL) reagents (Pierce, Rockford, IL, USA), the immune-signals were captured using the ChemiDoc image detector (Bio-Rad).
Statistical analysis
Statistical analysis was carried out by Student's t-test with SPSS statistical software (version 12.0). P value <.05 was considered statistically significant. Data are represented as means ± standard error of the mean from three independent experiments except where indicated.
Results
Platycodon suppresses weight gain in HFD-fed mice
As shown in Figure 1, the initial body weights were similar in all groups. At the end of 8 weeks, the body weight gain in Orlistat and Platycodon group was reduced by 25.56% and 7.50%, respectively. There were no significant differences in the food efficiency ratio of Orlistat and Platycodon groups with any alteration in food intake compared to the HFD group.

Effect of platycodon extract on body weight
Platycodon prevents change in lipid profiles
As shown in Table 1, the plasma levels of TG, TC, HDL-C, LDL-C, AI, and CRF in HFD group significantly increased compared with the normal group. Platycodon treatment significantly prevented the increases in TG and TC as well as the LDL-C levels. In addition, the HDL-C levels were significantly increased.
Effect of Platycodon Extracts on Lipid Profiles in Mice Fed a High-Fat Diet
AI = (TC – HDL-C)/HDL-C.
CRF = TC/HDL-C.
P < 0.05 versus HFD group.
AI, atherogenic index; CRF, cardiac risk factor; HDL-C, high-density lipoprotein cholesterol; HFD, high-fat diet; LDL-C, low-density lipoprotein cholesterol; TC, total cholesterol; TG, triglyceride.
Platycodon regulates the leptin and adiponectin content in plasma
The plasma leptin concentration was significantly higher in HFD group (62.09 ng/mL) and in the normal group (3.80 ng/mL). Both Orlistat and Platycodon groups lowered the leptin concentration by 50.04 and 51.30 ng/mL, respectively (Fig. 2A), whereas the plasma adiponectin concentration of Platycodon group was significantly increased compared to the HFD group (Fig. 2B).
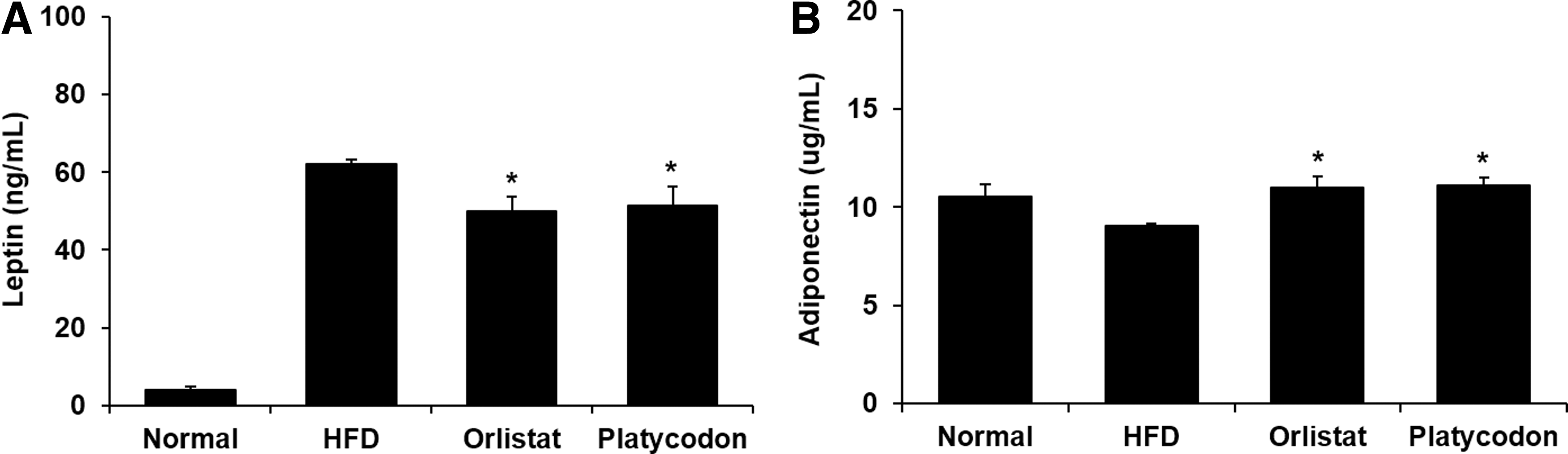
Effect of platycodon extracts on production of leptin
Platycodon suppresses the lipogenesis-related genes and protein expression in liver
To confirm that platycodon regulates lipogenesis in liver, we measured the lipogenesis-related genes and protein expression by real-time reverse transcription PCR and western blot analysis. HFD group showed significantly increased messenger RNA (mRNA) expression of fatty acid synthase (FAS), acetyl-CoA carboxylase (ACC), and lipoprotein lipase (LPL) (Fig. 3A), while Platycodon group markedly reduced the mRNA expression of these genes. Besides, the protein expression of FAS, ACC, and LPL in liver of HFD group was elevated compared with those of normal mice. However, the protein expression was significantly decreased in Platycodon group compared to the HFD group (Fig. 3C). These results suggest that platycodon extract may inhibit lipid accumulation by downregulating lipogenic-specific factors.
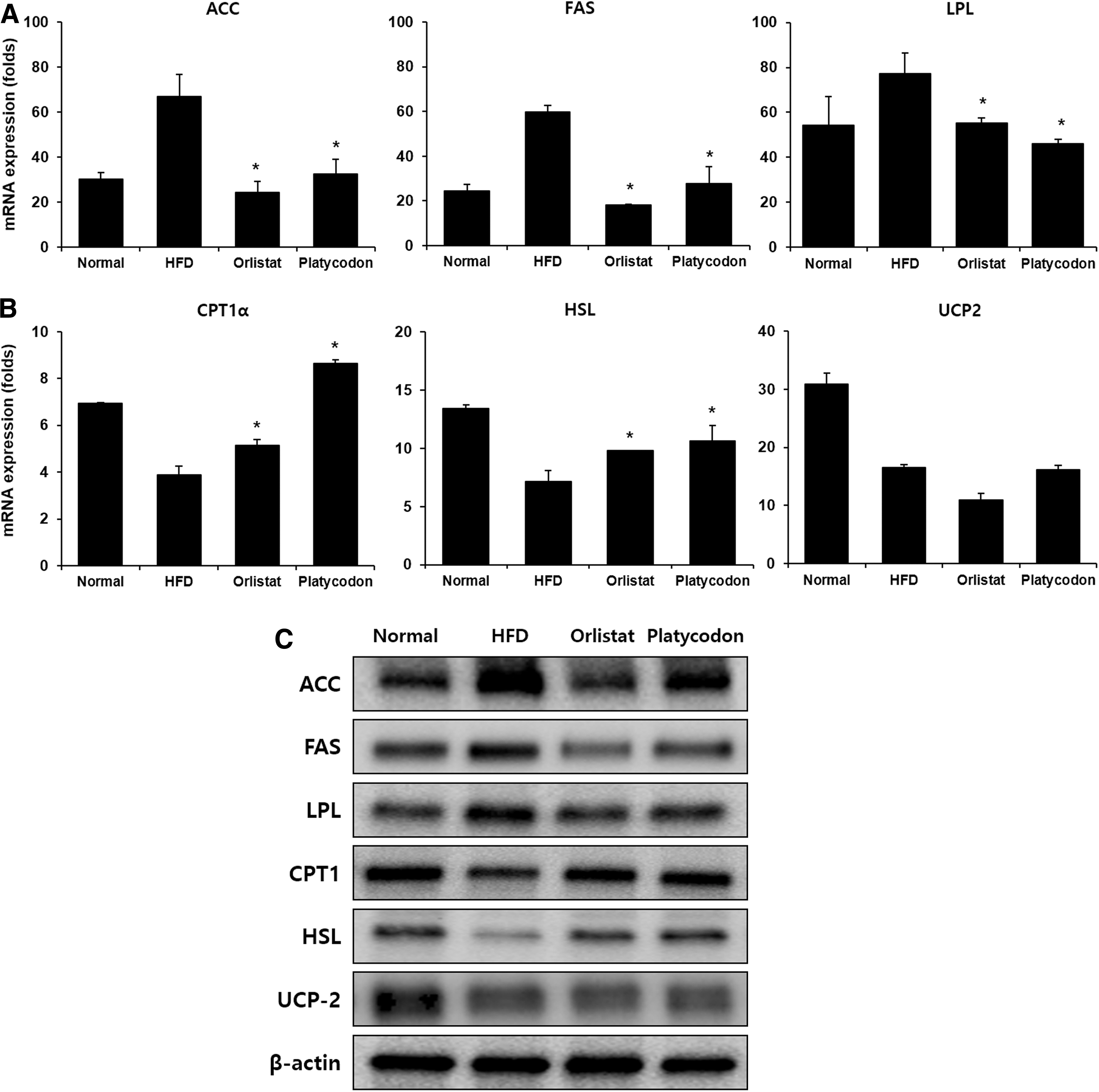
Effect of platycodon extracts on lipogenesis and fatty acid oxidation-related genes in the liver of mice fed HFD.
Platycodon upregulates the lipolysis-related genes and protein expression in liver
To confirm the antiobesity effect of platycodon extract due to the lipolysis in liver, the mRNA and protein expression of carnitine palmitoyltransferase 1α (CPT1α), hormone-sensitive lipase (HSL), and uncoupling proteins 2 (UCP2) in liver were measured. As shown in Figure 3B and C, platycodon treatment significantly increased CPT1α and HSL expression compared to HFD group, while platycodon had no effect on UCP2 expression in liver. This suggests that platycodon only enhanced fatty acid oxidation in liver but no effect on thermogenesis.
Platycodon regulates transcription factors of adipogenesis in liver
It is known that adipocytes secrete adipogenic-specific proteins such as PPARγ, SREBP-1c, and C/EBPα during differentiation for fat accumulation. 3 –5 Hence, we investigated the adipogenic-specific proteins levels in liver treated with platycodon. As shown in Figure 4, platycodon treatment significantly decreased the levels of PPARγ and C/EBPα in liver. These results suggest that the weight loss of adipose tissue in the platycodon treatment group could be attributed lipids not being used for hypertrophy of adipocytes due to the reduction of adipocyte differentiation.
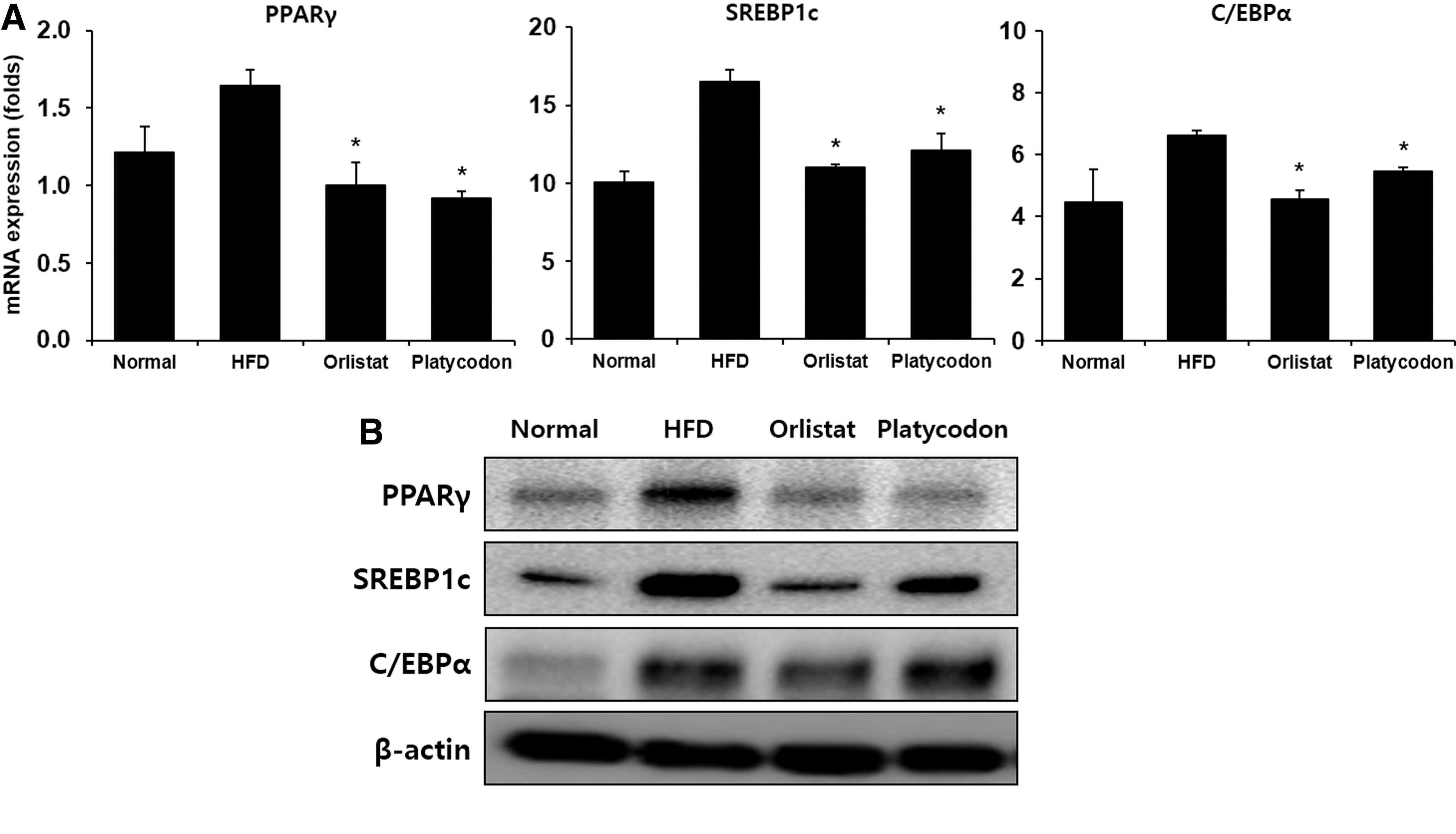
Effect of platycodon extracts on expression of adipogenic transcription factors in mice fed HFD.
Platycodon regulates lipogenesis-, lipolysis-, and adipogenesis-related gene expression in WAT
We investigated the effects of platycodon extracts on the expression of lipogenesis-, lipolysis-, and adipogenesis-related genes by the mRNA expression levels in the WAT. As shown in Supplementary Figure S1, in the HFD group, mRNA expression of ACC, FAS, and LPL was increased, however, in the Platycodon group, these genes were significantly decreased, whereas in the case of lipolysis-related genes, the mRNA expression of CPT1α, HSL, and PPAR-α in the Platycodon group was increased compared to the HFD group. The adipogenesis-related gene expression showed whether platycodon extracts affect adipose tissues of HFD group. PPARγ, C/EBPα, and SREBP-1c were markedly downregulated in the Platycodon group compared with HFD group.
Discussion
In this study, we investigated the antiobesity of platycodon extract in HFD-induced obese C57BL/6 mice. To confirm the hypothesized mechanisms, experiments evaluated the biochemical parameters associated with obesity.
Blood TG and HDL-C are negatively correlated. In the case of obesity, high TG and low HDL-C levels are typically shown in the blood. To improve the obesity, many people want to regulate the TG and HDL-C. 21 In our study, HFD increased the elevation of plasma TG levels. After platycodon treatment, the mice had lower TG levels in plasma which were similar to those in the normal group. These results are consistent with a previous study 22 that HFD-fed mice showed increase in TC and LDL-C and reduction in the HDL-C levels. Platycodon treatment also lowered the levels of TC and LDL-C in plasma and increased HDL-C. These results suggest that platycodon extract may inhibit obesity by lowering TG and increasing HDL-C thereby improving the lipid profile in blood.
The adipokines such as leptin and adiponectin secreted by adipose tissue regulate lipid metabolism. Leptin regulates appetite and energy metabolism by suppressing appetite and providing a feeling of satiety. Leptin is also known to increase in obesity. 23,24 Serum leptin concentration is an indicator of body fat mass. 25 On the contrary, adiponectin is known to be a hormone that decreases its expression level in obese people, reduces appetite, reduces inflammation, and promotes insulin sensitivity. 26 In addition, the higher the visceral fat, the lower the expression level of adiponectin, which is reported to increase the risk of obesity, cardiovascular, and metabolic disease. 27 In this study, we found that leptin levels were increased in the HFD group, and with platycodon intake it was significantly decreased. On the other hand, concentrations of adiponectin showed a tendency to increase in the Platycodon group. This suggests that intake of platycodon is effective in improving the changes of dyslipidemia and adipokine caused by high fat intake.
During lipogenesis, lipogenesis-related genes such as ACC, FAS, and LPL expression is increased and SREBP-1c induces the synthesis of fatty acids by controlling the expression of several genes that regulate fatty acid synthesis. 28 Expression of ACC, FAS, and LPL was increased in the liver of the HFD group, but decreased in the Platycodon group. This indicates that the TG reduction observed in this study is highly related to the expression of lipogenic genes. Also, we found that the Platycodon groups had significantly higher levels of lipolysis and fatty acid oxidation-related genes (CPT1α and HSL) in their liver tissue. This result suggests that platycodon can activate lipolysis and inhibiting adipogenesis pathways.
PPARγ, C/EBPa, and SREBP-1c are transcriptional factors that play a major role in the adipocyte's differentiation. PPARγ is expressed at an early stage during the adipocyte differentiation process, which promotes lipogenic gene expression. PPARγ is regulated by SREBP-1c, and SREBP-1c induces fatty acid synthesis such as ACC and FAS. 29,30 Recent studies have shown that the expression of SREBP-1c is reduced in leptin-treated mice, and that the reduced expression of SREBP-1c may have an effect on the inhibition of fatty acid synthesis. 31 This suggests a link between leptin and genes involved in lipogenesis, in this study, the decrease in leptin concentration by platycodon regulates the expression of SREBP-1c and PPARγ, resulting in the inhibition of FAS expression by participating in fatty acid synthesis. In this regard, it is considered that platycodon inhibits the formation and accumulation of fat in the body by inhibiting the proliferation and differentiation function of adipocytes by regulating the adipogenic gene.
Triterpenoid saponins such as platycodin A, C, and D and polygalacin D are the main pharmacological components that were reported to have antiobesity effects. Among various components, in particular, many studies of antiobesity effect on platycodin D have been reported. Platycodin D inhibits adipogenesis in 3T3-L1 cell 32 and ameliorates HFD-induced obesity in mice. 33 Recently, platycodin D has been reported to improve the obesity regulating adipogenic markers by AMP-activated protein kinase. 34 These are consistent with our results, we also confirmed that platycodon extracts regulate the expression adipogenic factors (Fig. 4). Zhao et al., 35 reported that platycodin shows antiobesity effects by inhibiting pancreatic lipase. Specific binding to lipase would promote the excretion of lipid with the stools. The fecal outputs in platycodin-treated group were increased by 2.1–3.2-fold in a dose-dependent manner, confirming that platycodin makes fecal TG, to be excreted without being digested. In our result, we also confirmed that fecal TG increased with platycodon extract treatment (Supplementary Fig. S2). To understand the TG metabolism in platycodon treatment, we performed oral lipid tolerance tests. Our results showed that the Platycodon group showed significantly reduced circulating TG after oral administration of lipid emulsion compared to HFD group (Supplementary Fig. S3). These results supported that platycodon extracts helps facilitate TG metabolism and increases TG fecal excretion, which contribute to in the antiobesity efficacy.
In conclusion, the present study revealed that platycodon extract was effective in preventing the increase in body weight gain, plasma TG, TC, LDL-C, and leptin levels and promotes the HDL-C and adiponectin levels. Moreover, platycodon extracts are associated with regulating of lipogenic-, lipolysis-, and adipogenic factors. These results are likely to be caused by complex mixture of bioactive compounds (triterpenoid saponins and sterol components, and so on). In the future, it will be necessary to study the identification of physiologically active substances contained in platycodon. The results of this study indicate that platycodon is a natural product that has the potential to be developed as a functional food and medicinal ingredients for improvement and treatment of obesity and other metabolic diseases in human.
Footnotes
Acknowledgment
This study was carried out with the support of the Research Program for Agricultural Science and Technology Development (Project No. PJ01169501, National Institute of Agricultural Sciences, Rural Development Administration, Republic of Korea).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.