
Abstract
We have previously demonstrated the hepatoprotective effect of Schisandra chinensis polysaccharides (SCP) against the liver injury induced by alcohol, high-fat diet, and carbon tetrachloride in mice. In this study, we investigated the effect of SCP against the immunological liver injury induced by concanavalin A (Con A) in mice. The results showed that SCP could significantly reduce the level of alanine aminotransferase (ALT), aspartate aminotransferase (AST), tumor necrosis factor-α (TNF-α), interferon-γ (IFN-γ), interleukin-1β (IL-1β), and interleukin-6 (IL-6) in the serum of mice with immunological liver injury. SCP could significantly decrease the content of malondialdehyde (MDA) and nitric oxide (NO) and increase the activity of superoxide dismutase (SOD) and glutathione (GSH) in the liver tissue. SCP could significantly increase the number of CD4+ and decrease the number of CD8+ in the peripheral blood, and elevate the ratio of CD4+/CD8+. SCP could significantly downregulate the expression of Kelch-like ECH-associated protein 1 (Keap1) and upregulate the expression of nuclear factor-erythroid 2-related factor2 (Nrf2) and downstream gene heme oxygenase-1 (HO-1), and downregulate the expression of toll-like receptor 4 (TLR4), myeloid differentiation primary response gene 88 (MyD88), and nuclear factor-kappa B (NF-κB) proteins. This study indicates that SCP can reduce the release of a large number of inflammatory factors to inhibit the oxidative stress in mice with the immunological liver injury induced by Con A, and its mechanism is closely related to the regulation of Nrf2/antioxidant response element and TLR4/NF-κB signaling pathways.
Introduction
Liver disease is one of the common diseases threatening human health and immunological liver injury is one of the important factors leading to acute liver failure, chronic hepatitis, and liver fibrosis. 1,2 Therefore, it is important to explore effective drugs to prevent and treat immunological liver injury. As a plant agglutinin, concanavalin A (Con A), with a mitosis-promoting activity, can activate T lymphocytes; the damage induced by it was liver specific and dose dependent. 3 The mouse liver injury model induced by Con A has been developed in recent years. Its establishment and application provide a very convenient and ideal experimental animal model for the further study of the pathogenesis of liver injury and the screening of therapeutic drugs. 4
Schisandra chinensis is the dried ripe fruit of S. chinensis Baill of Magnoliaceae, which is called “the king of protecting liver and lowering enzymes” as a traditional medicine for protecting liver in China. Our previous study showed that S. chinensis polysaccharides (SCP) has a good protective effect against the liver injury induced by alcohol, high-fat diet, and carbon tetrachloride in mice. 5 –7 However, the role of SCP on immunological liver injury has not been further studied. Therefore, this study investigated the protective effect of SCP against immunological liver injury and its mechanism.
Materials and Methods
Experimental animals, materials and reagents
Male ICR mice (6-week old) were provided by Changchun Yisi Experimental Animal Technology Co., Ltd. (Changchun, Jilin, China). The mice were kept in a sterile laboratory room, with free access to food and water, at 20–24°C, in humidity that was 45–55, and 12-h of light/12-h dark cycle. All animal experimental procedures were approved by Council of Animal Care and Use of Beihua University (no. 20180402).
Con A (type IV) for injection was obtained from Sigma (Louis, MO, USA); kits for the detection of alanine aminotransferase (ALT), aspartate aminotransferase (AST), superoxide dismutase (SOD), malondialdehyde (MDA), nitric oxide (NO), and glutathione (GSH) were obtained from Nanjing Jiancheng Bioengineering Institute (Nangjing, Jiangsu, China); ELISA kits for the detection of interleukin-1β (IL-1β), interleukin (IL)-6, tumor necrosis factor-α (TNF-α), and interferon-γ (IFN-γ) were obtained from Shanghai Lengton Bioscience Co, Ltd. (Shanghai, China); polyclonal antibodies nuclear factor-erythroid 2-related factor2 (Nrf2), heme oxygenase-1 (HO-1), Kelch-like ECH-associated protein 1 (Keap1), toll-like receptor 4 (TLR4), myeloid differentiation primary response gene 88 (MyD88), nuclear factor-kappa B (NF-κB), and β-actin were obtained from Abcam (San Francisco, USA); and electrogenerated chemiluminescence (ECL) color liquid was obtained from Biyuntian Biological Products Co, Ltd (Beijing, China).
Epics-XL Flow Cytometer (Beckman Coulter, USA); Infite M200 Microplate reader (TECAN, Switzerland); Western blot electrophoresis and electroporator (Bio-Rad, USA); ChampChemi Professional+ automatic multicolor fluorescence and chemiluminescence gel imaging system (Beijing Saizhi Technology Co, Ltd., China).
Preparation and analysis of SCP
The dried ripe fruit of S. chinensis was obtained from Ji'an City Rare Seedling Breeding Base in Jilin Province, China. The preparation method of SCP refers to our previous research (water extraction and alcohol precipitation method).
8
Total carbohydrate content of SCP was determined by phenol-sulfuric acid method with glucose as standard.
9
Using
The extract of 129.5 g SCP was obtained from 1.5 kg S. chinensis, with a yield of 8.67%. The chemical properties of SCP are listed in Table 1. The samples are kept in Pharmacy Laboratory, College of Pharmacy, Beihua University (sample no.: 20171013-1).
Chemical Properties of Schisandra chinensis Polysaccharides
Ara, arabinose; Car, carbohydrate; Gal, galactose; GalA, galacturonic acid; Glc, glucose; GlcA, glucuronic acid; Man, mannose; Pro, protein; Rha, rhamnose; UroA, uronic acid.
Animal grouping and administration
Mice were randomly divided into five groups, control (CON), model (MOD), low-dose SCP (SCP-L), medium-dose SCP (SCP-M), and high-dose SCP (SCP-H), with 15 mice in each group. Mice in the SCP-L, SCP-M, and SCP-H groups were intragastrically given the corresponding doses of SCP aqueous solution (0.2, 0.4, and 0.8 mg/10 g), and those in CON and MOD groups were intragastrically given the same volume of distilled water, successively for 21 days. On the 20th day after the administration, mice in the MOD, SCP-L, SCP-M, and SCP-H groups were intravenously injected with Con A (40 mg/kg) and mice in CON group were intravenously injected with a same volume of normal saline. Eight hours after the injection of Con A, the mice were anesthetized with ether and the blood samples were collected by removing the eyeballs of mice, and then the mice were killed by inhalation of CO2; the serum, liver, spleen, and thymus were isolated and stored at −80°C.
Determination of organ indexes and histomorphology observation
The liver, spleen, and thymus were weighed after the excess tissue and fascia on them were removed. The liver, spleen, and thymus indexes were calculated according to the following equation:
The liver, spleen, and thymus were fixed in 10% formalin solution at room temperature for 24 h, and then routinely sectioned (5 μm). The slices were embedded in paraffin and stained with hematoxylin-eosin staining for 10 min at room temperature. Pathological changes were observed under an optical microscope (magnification, 100 ×).
Determination of serum ALT, AST, TNF-α, IFN-γ, IL-1β, and IL-6
ALT and AST contents in the serum were detected by following the instructions of the kits, and TNF-α, IFN-γ, IL-1β, and IL-6 in the serum were detected by ELISA method.
Determination of MDA, SOD, GSH, and NO
Around 0.2 g of the left liver lobe was taken, and 10% homogenate of the liver was prepared. The levels of MDA, NO, SOD, and GSH were determined according to the instructions of the kits.
Determination of CD4+ and CD8+ percentages and CD4+/CD8+ ratios
The blood was placed in test tubes treated with heparin for anticoagulation, and fully vibrated and evenly mixed. CD3+ was labeled with fluorescein isothiocyanate (FITC), CD4+ was labeled with PE-CY5, and CD8+ was labeled with phycoerythrin (PE), and the positive expression rates were detected by flow cytometry.
Determination of Nrf2, HO-1, Keap1, TLR4, MyD88, and NF-κB protein expressions
Nrf2, HO-1, Keap1, TLR4, MyD88, and NF-κB protein expressions in the liver tissue of mice were detected by Western blot. Fifty milligrams of the liver tissue of mice was cut into pieces, and then placed in radio immunoprecipitation assay (RIPA) lysis buffer containing protease inhibitor on ice for 1 h for the lysis of cells, and the lysate was centrifuged at 12,000 g for 5 min to obtain the supernatant. The protein content in the supernatant was determined by butyleyanoacrylate (BCA) method. The proteins were separated on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred onto a polyvinylidene difluoride membrane. After the blocking with 5% skim milk for 1.5 h, the first antibodies were added onto the membrane, which was incubated at 4°C overnight, and the membrane was washed with Tris-HCl buffer saline Tween (TBST) three times, 10 min each time, and then the second antibodies were added onto the membrane at room temperature, which was incubated for 1 h, and after the membrane was washed with TBST three times, 10 min each time, ECL developing liquid was added onto it. The images were taken using chemiluminescent imaging system (FluorChem, CA, USA), and β-actin was used as the internal reference to compare the expression of proteins among the different groups. The gray values of bands in each group were determined by ImageJ (image processing and analysis in Java), and the ratio of the gray value to the gray value of β-actin in each group was taken as the relative expression of proteins.
Statistical analysis
All values are expressed as mean ± standard deviation. The number of samples in each group was expressed as “n”. SPSS software (Windows version 19.0) was used for the statistical analysis. One-way analysis of variance was used for the comparison between groups. P < .05 was considered significant in statistics.
Results
Effects of SCP on organ indexes
Compared with those in the control group, the liver, spleen, and thymus indexes increased significantly in the model group (P < .05). Compared with those in the model group, the liver, spleen, and thymus indexes decreased significantly in SCP-treated groups (P < .05, P < .01). The results are shown in Figure 1.
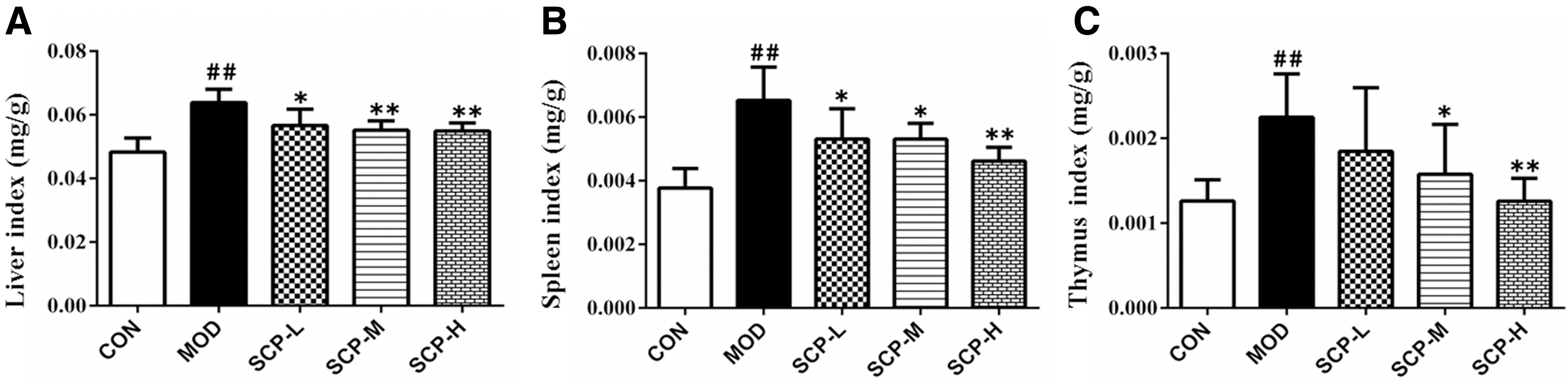
Effects of SCP on organ indexes.
Effects of SCP on the histomorphology
As shown in Figure 2, the intralobular beam in the thymus could be seen, the size and shape of the thymic cortex were normal, the medulla inside each cortex of thymus was visible, and the medullary structure was clear in the control group. Compared with those in the control group, lymphocytes in the thymic cortex were decreased, the boundaries between the medulla and cortex were unclear, and the volumes of medulla decreased in the model group. Compared with those in the model group, the medullary and cortical structures were clear, the boundaries between the medulla and cortex were distinct, and the number of lymphocytes was normal in SCP-H and SCP-M groups, while the boundaries between the medulla and cortex were not clear in SCP-L group, but slightly clearer than those in the model group.
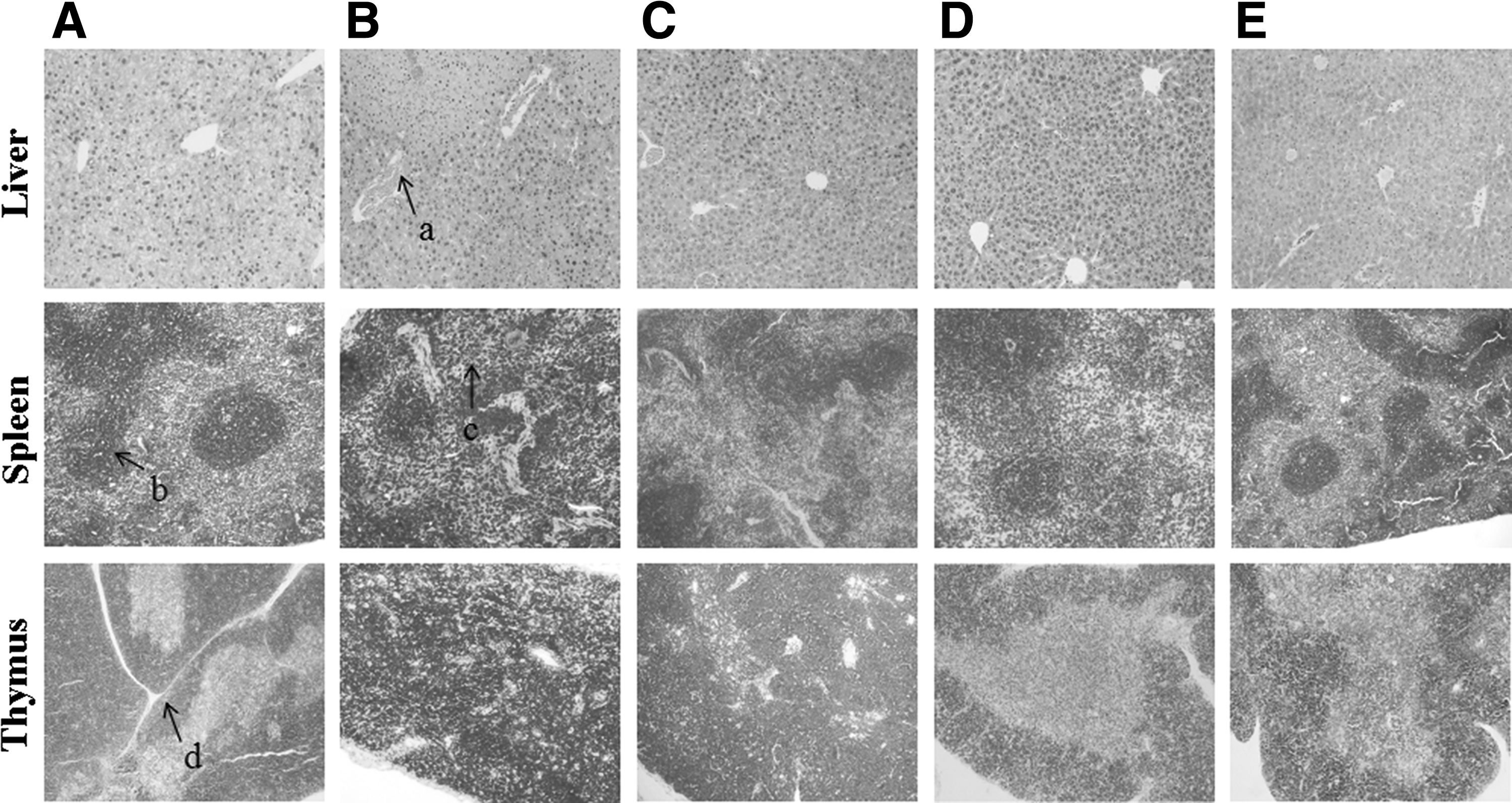
Effects of SCP on histomorphological changes in the liver, spleen, and thymus of mice.
The histomorphological observation on the spleen showed that the number and shape of splenic corpuscles were normal and the boundary between red and white pulp was clear in the control group. Compared with that in the control group, the number and volume of splenic corpuscles decreased significantly in the model group, the number of splenic corpuscles in the membranous area decreased slightly in the SCP-L group, the number of splenic corpuscles in SCP-H and SCP-M groups was similar to that in the control group, and the boundary between white and red pulp was clear in SCP-H and SCP-M groups.
The histomorphological observation on the liver showed that the hepatic lobules were intact, the hepatic cells were arranged regularly, and the central hepatic vein and the hepatic hilar region were normal in the control group; the edema and local necrosis of hepatocytes, and the dilatation and hyperemia of central vein could be seen, which were mainly located around the central vein in the model group, indicating the liver tissue injury was serious; the structure of hepatic lobules was almost normal and the hepatic sinusoidal congestion was alleviated obviously in each SCP-treated group.
Effects of SCP on the number and proportion of CD4+ and CD8+
As shown in Table 2 and Figure 3, compared with that in the control group, the number of CD4+ was significantly lower (P < .05), the number of CD8+ was significantly higher (P < .05), and the ratio of CD4+/CD8+ was significantly decreased in the model group (P < .05); compared with that in the model group, the number of CD4+ and the ratio of CD4+/CD8+ were increased, and the number of CD8+ was decreased in each SCP-treated group (P < .05).
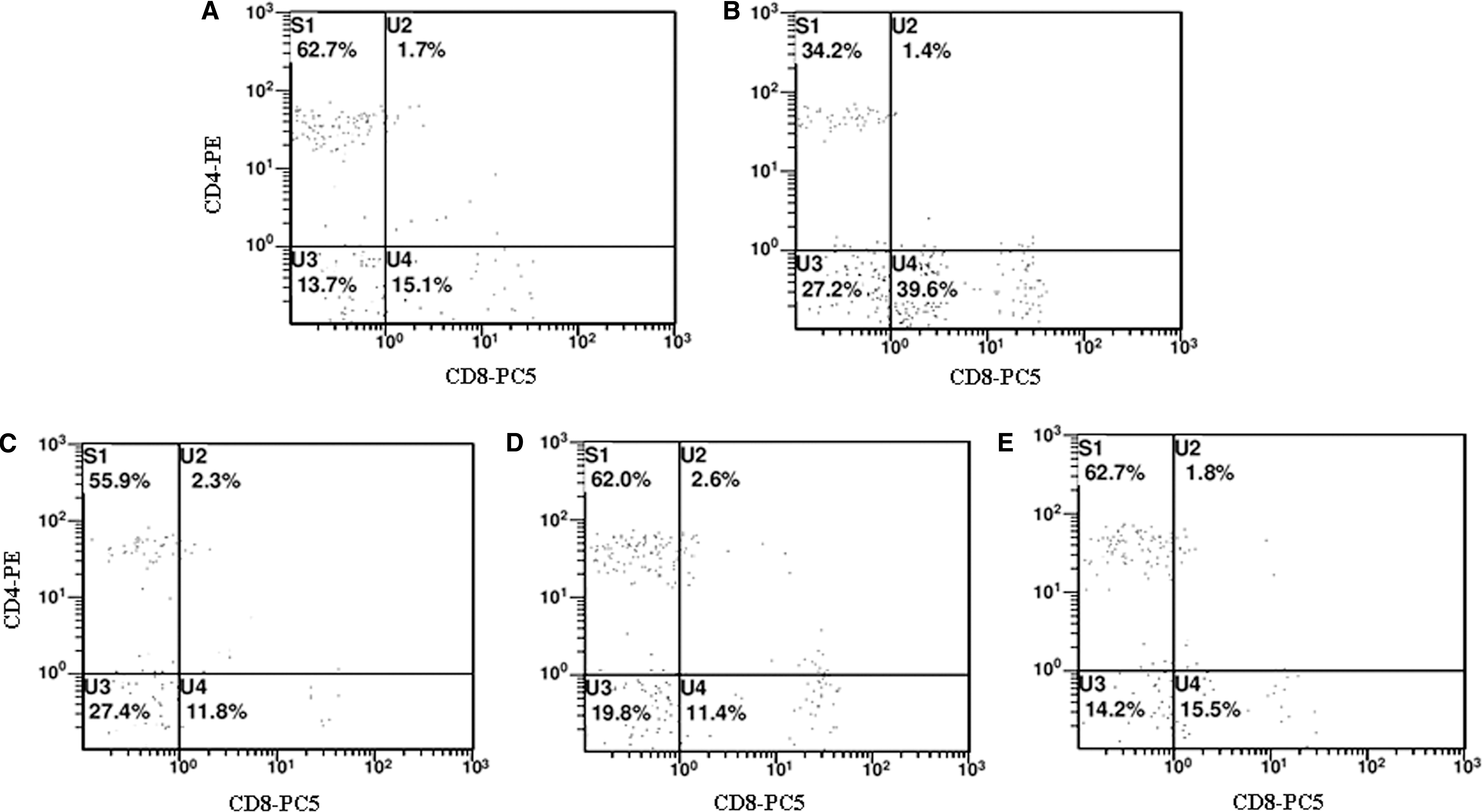
Effects of SCP on the number and proportion of CD4+ and CD8+ T lymphocytes in the peripheral blood.
Effects of Schisandra chinensis Polysaccharides on the Number and Proportion of CD4+ and CD8+ T Lymphocytes in the Peripheral Blood
Values are expressed as the mean ± standard deviation (n = 15).
P < .05 and ## P < .01 versus the control group.
P < .05 and ** P < .01 versus the model group.
CON, control group; MOD, model group; SCP-H, S. chinensis polysaccharide high-dose group; SCP-L, S. chinensis polysaccharide low-dose group; SCP-M, S. chinensis polysaccharide medium-dose group.
Effects of SCP on the serum AST and ALT levels
As shown in Figure 4, compared with those in the control group, ALT and AST levels in the serum of mice in the model group were significantly higher (P < .01); compared with those in the model group, ALT and AST levels in the serum of mice in SCP-treated groups were significantly lower (P < .05, P < .01).
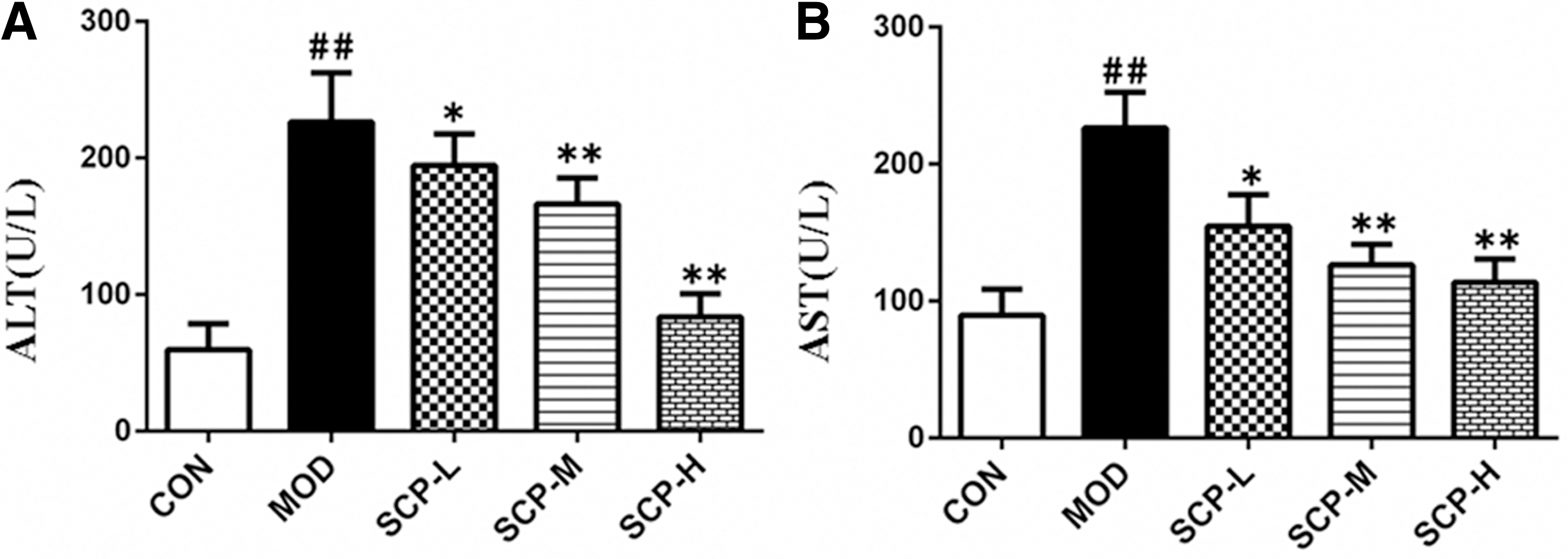
Effects of SCP on serum AST and ALT levels.
Effects of SCP on TNF-α, IFN-γ, IL-1β, and IL-6 levels
As shown in Figure 5, compared with those in the control group, TNF-α, IFN-γ, IL-1β, and IL-6 contents of mice in the model group increased significantly (P < .05, P < .01); compared with those in the model group, TNF-α, IFN-γ, IL-1β, and IL-6 contents of mice in SCP-treated groups decreased significantly (P < .05).
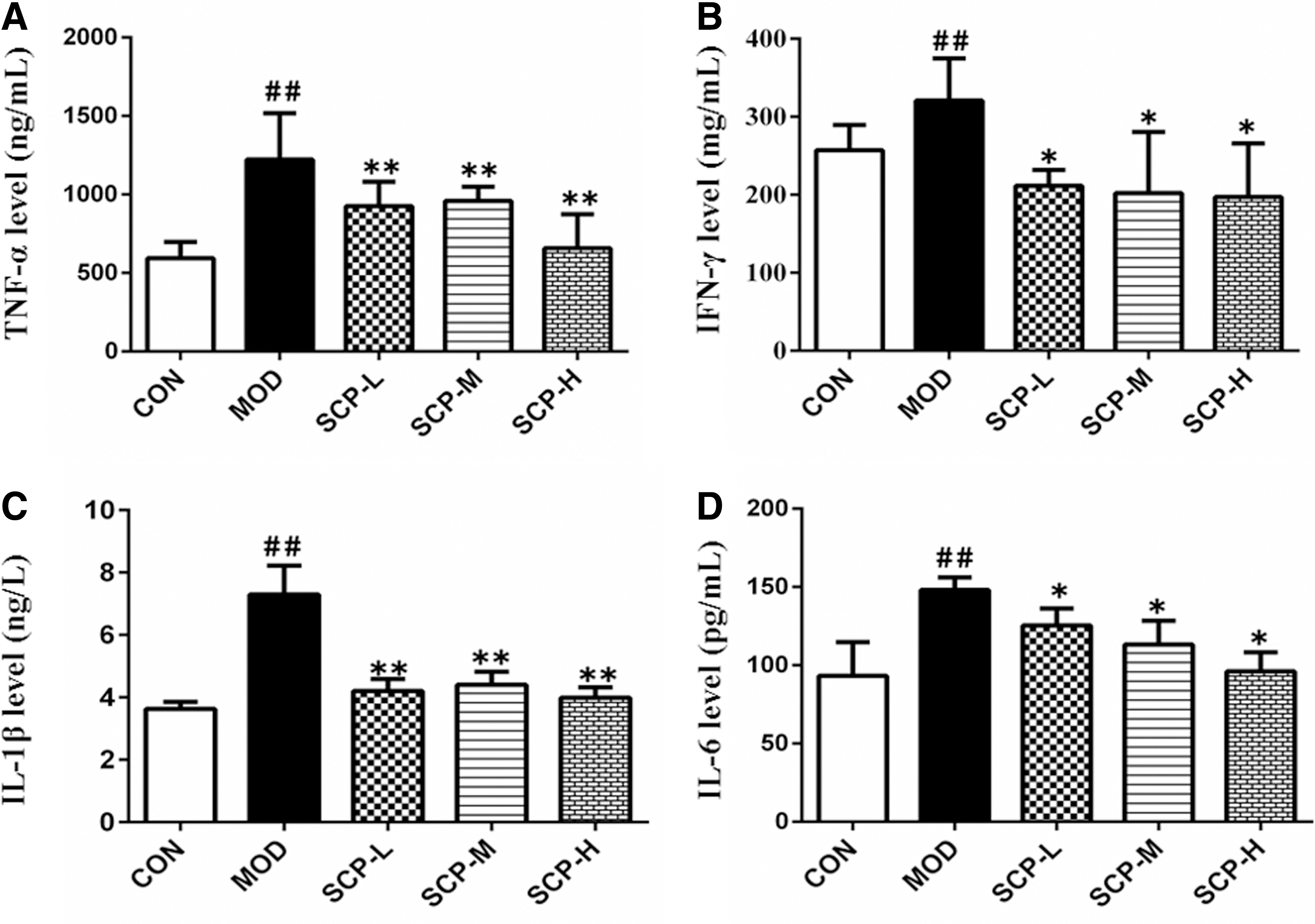
Effects of SCP on TNF-α, IFN-γ, IL-1β, and IL-6 contents in the serum.
Effects of SCP on MDA and NO levels and SOD and GSH activities
As shown in Figure 6, compared with those in the control group, the MDA and NO levels increased significantly (P < .01), and the SOD and GSH activities decreased significantly (P < .01) in the liver tissue of mice in the model group; compared with those in the model group, the MDA and NO levels decreased significantly (P < .05, P < .01), and the SOD and GSH activities increased significantly (P < .05, P < .01) in the liver tissue of mice in SCP-treated groups.
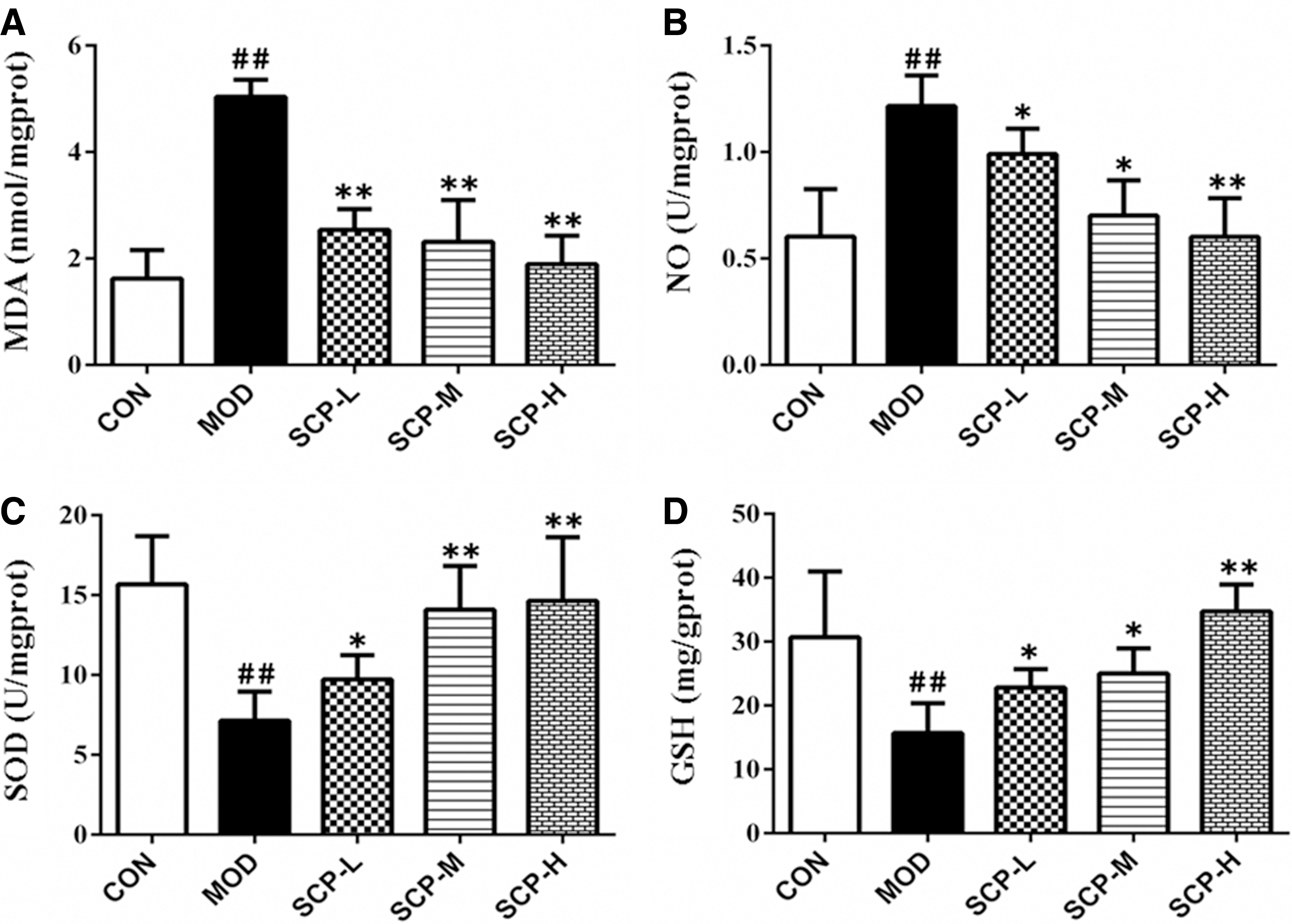
Effects of SCP on MDA and NO levels and SOD and GSH activities in the liver tissue.
Effects of SCP on the expression of Nrf2, HO-1, and Keap1 proteins
As shown in Figure 7, compared with that in the control group, the expression of Keap1 protein was significantly elevated (P < .01), and the expression of Nrf2 and HO-1 proteins was significantly reduced (P < .01) in the model group; compared with that in the model group, the expression of Keap1 protein was significantly reduced (P < .05, P < .01), and the expression of Nrf2 and HO-1 proteins was significantly elevated (P < .01) in SCP-treated groups.
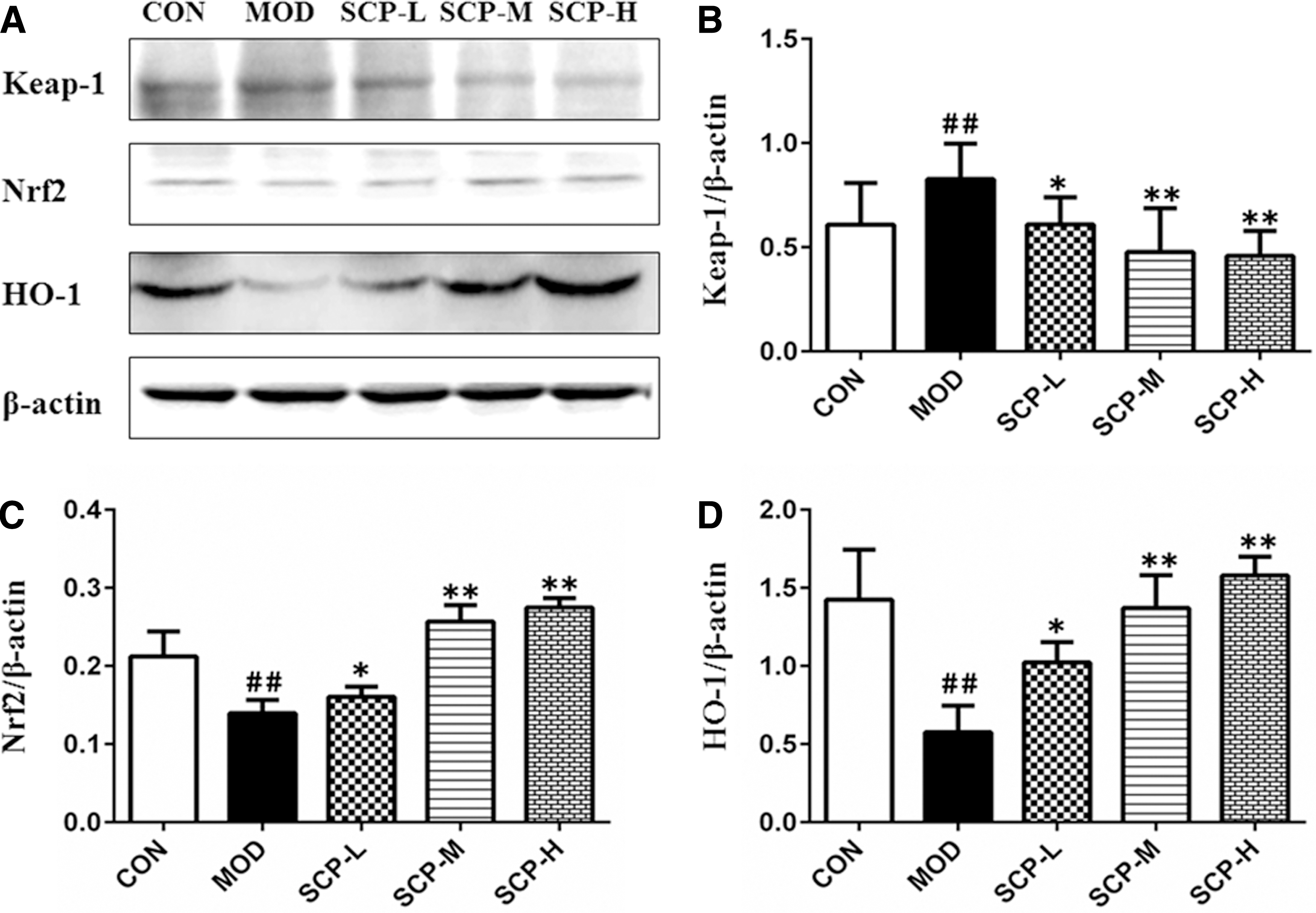
Effects of SCP on the expression of Keap1, Nrf2, and HO-1 proteins in the liver tissue.
Effects of SCP on the expression of TLR4, MyD88, and NF-κB proteins
As shown in Figure 8, the expression of TLR4, MyD88, and NF-κB in the model group was significantly higher than that in the control group (P < .01), and the expression of TLR4, MyD88, and NF-κB in SCP-treated groups was significantly lower than that in the model group (P < .05, P < .01).
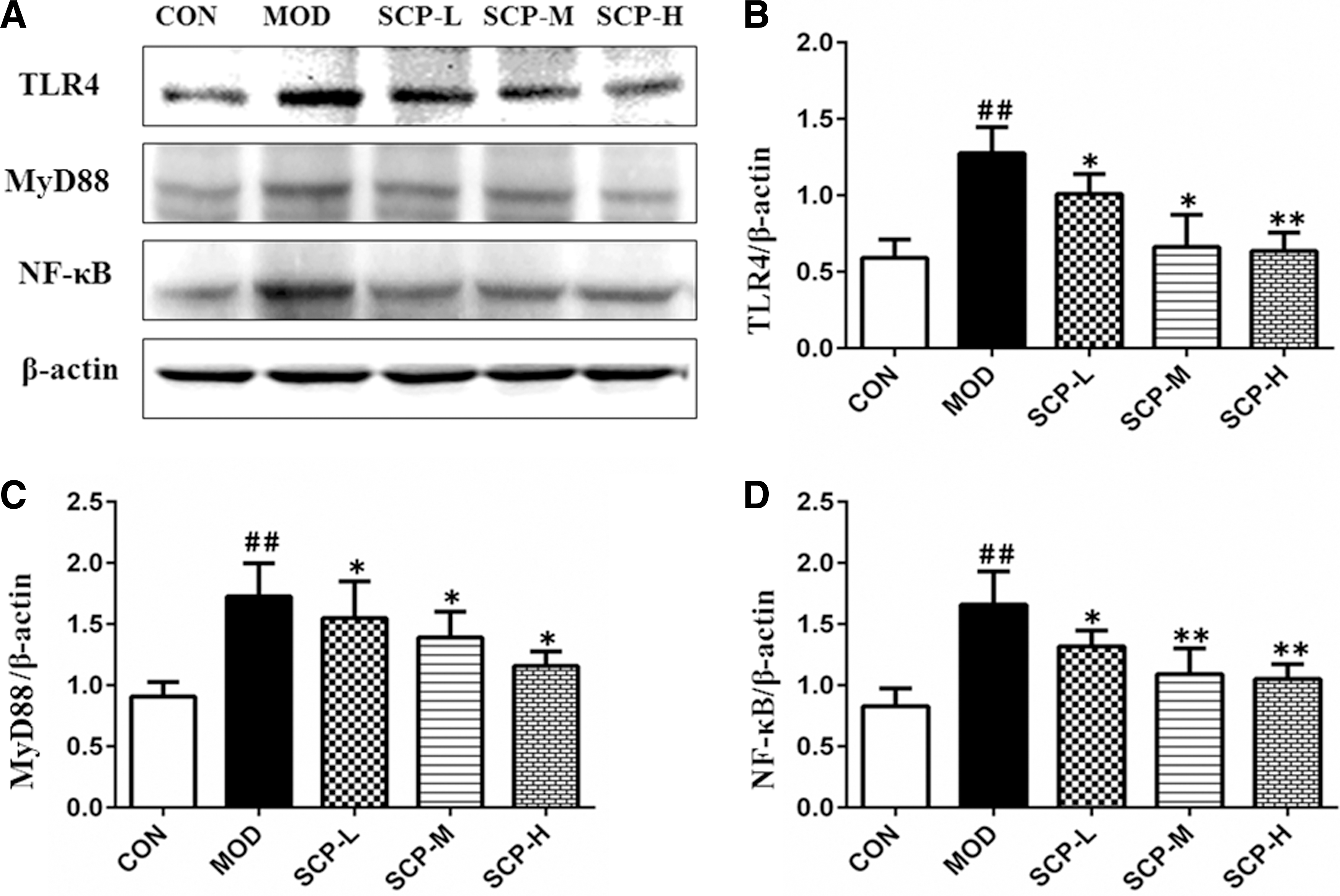
Effects of SCP on the expression of TLR4, MyD88, and NF-κB proteins in the liver tissue.
Discussion
Con A is a widely used mitogen that can activate T lymphocytes, and the immunological liver injury model induced by Con A in mice in this study is considered to be an ideal one used for the study on pathological mechanisms of human viral hepatitis and autoimmune liver diseases. 13 After the induction of Con A, a large number of immune cells accumulate in the liver and a large number of free radicals are produced, which can attack the unsaturated fatty acids in the membrane phospholipid to damage the liver cell membrane and mitochondrial membrane, leading to the increase of membrane permeability to increase the serum ALT and AST levels, and a large area degeneration of liver tissue cells with the severe infiltration, necrosis, and apoptosis of inflammatory cell. 14,15
Immune organ index is an initial index of immune function, and liver index can reflect the degree of liver injury and inflammation. After the induction of Con A, the proliferation of excessive lymphocytes can be stimulated to lead to a massive production of proinflammatory factors such as TNF-α, IL-1β, and IL-6, which may activate the TLR4/NF-κB signaling pathway, and then further promote the release of downstream inflammatory factor TNF-α. A large number of activated immune cells cause an excessive immune response and the enlargement of spleen and thymus, and subsequently, a large number of activated immune cells accumulate in the liver, causing the enlargement and injury of liver. 16,17
T cells are a large cell subset with the most important functions in the body's immune system. The abnormal number and function of the different lymphocyte subsets may lead to immune disorders and a series of pathological changes, and CD4+ T lymphocytes are mainly composed of Th, which can promote the proliferation and differentiation of B cells, T cells, and other immune cells, and coordinate the interaction among immune cells. 18 Con A mainly induces the immune response mediated by CD4+ T lymphocytes (Th cells) and the CD8+ T lymphocytes were mainly composed of cytotoxic lymphocytes; the increase or decrease in the proportion of lymphocyte subsets reflects the corresponding changes of the body's immune functions, and changes in the ratio of CD4+/CD8+ indicate the out-of-balance of the body's immune state. 19 The results showed that SCP could increase the level of CD4+ and decrease the level of CD8+, and increase the ratio of CD4+/CD8+, suggesting that SCP can effectively protect the hepatocytes and alleviate the liver injury.
Excessive TNF-α in vivo is an important factor to induce the damage of liver cells. TNF-α, a highly active inflammatory factor secreted by activated Kuppfer cells in the liver, with a cytotoxic function, can activate T and B lymphocytes and enhance the activity of natural killer cell and lymphocytes, thus causing a liver cell immunological injury, and also induce the secretion of IL-1, IL-6, and other cytokines to aggravate the inflammatory response of the liver and mediate the damage of liver cells. 20 –22 IFN-γ secreted by cytotoxic T cells can enhance the function of natural killer (NK) cells and macrophages to kill and engulf the target cells, and a large number of activated T lymphocytes and IFN-γ accumulated in the liver destroy the vascular endothelial cells, resulting in liver injury. 23
As the most important transcription factor of inflammatory response, NF-κB can bind to the fixed nucleotide sequence in the promoter region of many cytokines and inflammatory mediators to activate the transcription, playing an important role in the immune response and inflammatory response in the body, and can be activated by inflammatory cytokines and then induce the increase in the expression of inflammatory cytokines. 24 TLRs are immune recognition receptors and the TLR4 on the surface of liver sinusoidal endothelial cells is one of the most widely studied member in TLR family. After activation, TLR4 mediates the signal transduction by recruiting MyD88, and then activates nuclear factor NF-κB to initiate the TLR4/NF-κB signaling pathway to promote the release of downstream inflammatory factors TNF-α, IL-1, IL-2, IL-6, and IL-8, which triggers the inflammatory cascade, regulates immune cells and inflammatory cells, and participates in the proliferation, differentiation, and apoptosis of immune cells, thereby exacerbating the liver injury. 25,26 TLR4/NF-κB signaling pathway, an important link in the mechanism of immune response, is involved in the formation of immunological liver injury. It was found in this study that SCP could downregulate the overexpression of TLR4, MyD88, and NF-κB proteins by regulating the TLR4-MyD88-NF-κB signaling pathway, and then inhibit the expression of IL-6, IL-1β, and TNF-α and IFN-γ, playing an anti-inflammatory role to reduce the liver injury.
Immunological liver injury is accompanied by a severe oxidative stress, in which NO is a typical free radical in vivo and acts as a neurotransmitter to mediate the immune response, and when liver injury occurs, the synthesis of NO in Kuppfer cells in the liver increases and the liver is in the state of oxidative stress. 27 SOD, one of the important antioxidant enzymes in the body, can scavenge the excessive free radicals in the body, thus alleviating the liver injury, GSH-Px can better scavenge low molecular free radicals, hydrogen peroxide and lipid peroxides, and both have significant antioxidation. 28 MDA, an end product of lipid peroxidation, can seriously destroy the structure of cell membrane to cause the swelling and necrosis of cells, so the level of MDA can reflect the degree of lipid peroxidation and cell injury. 29
Nrf2, one of the main regulatory factor of antioxidant stress, interacts with antioxidant response element (ARE) to regulate the expression of downstream antioxidant enzymes and enhance the ability of cells to scavenge reactive oxygen free radicals, thus alleviating the injury of cells, tissues, and organs induced by oxidative stress. 30 –32 Keap1 is a multiregion repressor protein of Kelch family in the cytoplasm and inhibits the expression of Nrf2, and Nrf2 is an important transcription factor related to the self-protection of cells; when they are stimulated by oxidative stress, Keap1 and Nrf2 are uncoupled, the Nrf2 dislocates into the nucleus to bind to Maf protein to form heterogeneous dimer, and then the dimer binds to ARE to activate the expression of downstream target genes such as HO-1, thus playing a role of antioxidant damage. 33 –35 Our results showed that SCP could decrease the expression of Keap1 and increase the expression of Nrf2 and its downstream gene HO-1 by activating the Nrf2 signaling pathway to increase the activity of GSH and SOD and decrease the content of MDA and NO, inhibiting the occurrence of oxidative stress to alleviate the liver injury.
To sum up, SCP can reduce the content of serum transaminases in mice with immunological liver injury induced by Con A, improve the pathological injury of life, such as with inflammation, hemorrhage, and necrosis, and regulate the Nrf2/ARE and TLR4/NF-κB signaling pathways, while at the same time alleviate the oxidative stress and inflammatory reaction (Fig. 9), thus playing a protective effect against immunological liver injury.

Potential mechanism of hepatoprotection effect of SCP on the immunological liver injury induced by concanavalin A.
Footnotes
Acknowledgments
This work was supported by the Natural Science Foundation of Jilin Province (no. 20170309006YY; 20170307016YY) and Science and Technology Project of Jilin Provincial Education Department (no. JJKH20180373KJ).
Author Disclosure Statement
No competing financial interests exist.