
Abstract
The present study investigated the immunomodulatory activity and associated mechanisms of heat-treated Lactobacillus plantarum LM1004 (HT-LM1004) in a cyclophosphamide (CTX)-induced mouse model of immunosuppression. HT-LM1004 induced phagocytic activity and nitric oxide production in RAW264.7 macrophages and stimulated the release of tumor necrosis factor (TNF)-α, interferon (IFN)-γ, interleukin (IL)-2, and IL-12p70. In mice with CTX-induced immunosuppression, oral HT-LM1004 administration restored thymus and spleen indices, including spleen weight. Consistent with the in vitro results, HT-LM1004 increased TNF-α, IFN-γ, IL-2, and IL-12p70 levels in mice after 14 days of treatment and enhanced the natural killer (NK) cell activity of splenocytes from mice with CTX-induced immunosuppression against YAC-1 lymphoma cells. The method of HT-LM1004 generation influenced this activity: L. plantarum LM1004 grown in a membrane bioreactor, which reduced the size of the cells to <1.0 μm through physical stress (micronization), promoted NK cell cytotoxicity to a greater extent than LM1004 subjected to heat treatment alone. These findings indicate that HT-LM1004 without or with micronization can reverse CTX-induced immunosuppression without adverse side effects by potentiating NK cell function.
Introduction
Dysregulation of the immune system can lead to a variety of diseases. 1,2 Immunosuppression induced by chemotherapy causes systemic toxicity in patients. 3 Cyclophosphamide (CTX) is an alkylating cytotoxic drug that is effective in the treatment of cancer and autoimmune diseases in part because of their ability to modulate immune responses. 4 However, high doses of CTX administered over a long period of time can lead to immunosuppression and intestinal problems such as viral infection and gut intestinal dysbiosis. 5,6 In immunocompromised patients, bacterial and fungal infections can be fatal if left untreated. 7 The risk of infection can be mitigated by enhancing immune responses using natural or chemical adjuvants. However, immunoregulatory agents, especially those comprising a single component, can cause side effects such as fever, headache, hypertension, and neurotoxicity. 8 Safe immune-potentiating agents that can prevent immunodeficiency in cancer patients undergoing chemotherapy have been investigated, 9 including dietary supplements and functional foods that promote health. 10
Probiotic foods have been shown to improve immune health status and gut intestinal dysbiosis in immunocompromised patients. 11 Probiotics are defined as live microorganisms that exhibit beneficial effects on the host when administered at an appropriate dose, 12 including enhancement of the immune response; improvement of intestinal health, serum hypercholesterolemia, and hyperlipidemia; and cancer prevention. 13 Lactic acid bacteria (LAB) in fermented foods are probiotics that act as immune-potentiating agents but are also normal constituents of healthy human intestinal microflora. 14 Several Lactobacillus species, including Lactobacillus rhamnosus, Lactobacillus acidophilus, Lactobacillus casei, and Lactobacillus reuteri, have been shown to exert protective effects against pathogenic infection by regulating immunity. 14 –16 Lactobacillus plantarum is found in many fermented foods as well as anaerobic plant matter, and is widely used as a starter for fermented dairy products. 17
Lactobacilli can stimulate the immune system and enhance host defense against pathogens. 18 –20 The main LAB species in Korean kimchi, a traditional Korean fermented vegetable, is L. plantarum, which is a probiotic with strain-specific immunomodulatory activity that can alleviate chronic intestinal inflammation and irritable bowel syndrome. 15,21 Lactobacilli also stimulate the expression of interleukin (IL)-12 and interferon (IFN)-γ in monocytes and induce their differentiation into type 1 helper T cell subsets. 22,23 Additionally, live L. plantarum HY7712 isolated from kimchi reversed the inhibition of natural killer (NK) and T cell activities caused by CTX and γ-irradiation in mice. 24,25
We previously isolated L. plantarum strain LM1004 (KCCM 43246) from Korean kimchi, which has been filed with LactoMason Co., Ltd. (Jinju, Korea). We generated the heat-treated (HT) or micronized/HT (MHT) forms of LM1004 (HT-LM1004 and MHT-LM1004) by incubating live cells under conditions of physical stress in a specially designed membrane bioreactor (MBR; Korea patent No. 10-1245208). MHT-LM1004 is a dead, shrunken, granular, and smaller (<0.5 to 1.0 μm in size) form of the rod-shaped live L. plantarum LM1004.
In this study, we evaluated the immune-enhancing potential of HT-LM1004 in vitro by measuring changes in nitric oxide (NO) and cytokine levels in mice splenocytes and in vivo using a mouse model of CTX-induced immunosuppression. We also compared the efficacy of MHT-LM1004 and HT-LM1004 in promoting immune cell activity using mouse splenocytes. Our results demonstrated that HT-LM1004 can be used to restore a normal immune status in immunocompromised mice.
Materials and Methods
Materials
Roswell Park Memorial Institute (RPMI) 1640 medium, fetal bovine serum (FBS), penicillin, and streptomycin were obtained from Life Technologies (Carlsbad, CA, USA). Enzyme-linked immunosorbent assay (ELISA) kits for tumor necrosis factor (TNF)-α, IFN-γ, IL-2, IL-4, IL-10, and IL-12p70 were from R&D Systems (Minneapolis, MN, USA). 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) was from USB Corp. (Cleveland, OH, USA), and the lactate dehydrogenase (LDH) release detection kit was from Roche Applied Science (Indianapolis, IN, USA). All kits were used according to the manufacturers' protocols. CTX monohydrate, lipopolysaccharide (LPS), concanavalin (Con)A, and dimethyl sulfoxide (DMSO) were from Sigma-Aldrich (St. Louis, MO, USA).
Preparation of HT-LM1004 and MHT-LM1004
LM1004 was incubated for 48 h at 37°C in de Man, Rogosa, and Sharpe (MRS) broth (BD Biosciences, Franklin Lakes, NJ, USA). The cells were harvested by centrifugation (3000 g for 15 min at 4°C), washed twice with phosphate-buffered saline (PBS), and preserved as a glycerol stock (molecular biology grade, 25% w/w; Merck, Darmstadt, Germany) in a −70°C ultrafreezer and used as an inoculum as needed. Frozen stocks of L. plantarum LM1004 were propagated in a 300-L working volume/500-L stirred tank reactor (STR) batch fermenter through a series of steps that included activation and seed culturing. The cells were cultured at 37°C, and the pH in the STR was maintained at 6.0 with ammonia solution (20% v/v) throughout the production of HT-LM1004. The growth medium for batch culture contained 40 g/L glucose, 10 g/L soy peptone, 10 g/L yeast extract, 0.02 g/L
L. plantarum LM1004 was cultured in an MBR to generate MHT-LM1004. Bacteria can acclimatize to environmental stress for survival; in particular, cell wall shear stress caused by membrane geometry, and rotating pump and quorum sensing caused by extremely high cell density during culture in the MBR, 26 can lead to a reduction in cell size 27,28 but also large-scale production, which may be advantageous for enhancing the host immune system.
L. plantarum LM1004 was propagated in the MBR by seed cultivation of an 18-L working volume in a 20-L culture bottle purged with N2 gas from a frozen stock; the seed culture was inoculated into the MBR. The pH was maintained at 6.0 with ammonium solution (20% w/w). The same growth medium used for batch culture was used for the 18-L seed culture; the feed medium during culture in the MBR contained 50 g/L glucose, 10 g/L soy peptone, 13 g/L yeast extract, 0.02 g/L
MBR design
The design of the MBR is shown in Figure 1. The total capacity of the MBR used in this study was 15 L. Cells circulated in culture broth through the MBR, which consisted of a reactor (5.0 L), co-centric hygienic centrifugal pump, membranes, and pipelines. A ceramic membrane (3.5 m2 effective area, 0.22 μm pore size, Al2O3; Pall Corp., Port Washington, NY, USA) fitted with a medium-mixing tank (100 L) was used to sterilize fresh feeding medium during culturing by filtering out particles or contaminants before the medium was fed into the MBR. Another membrane with the same specifications was fitted externally onto the MBR to remove harmful waste material from the culture broth. The dilution rate for replacing the harmful wastes with fresh medium was controlled from 0 to 0.99/h by a pump and level gauge fitted in the bioreactor (5.0 L). The membranes were fitted in the stainless steel housing (SUS 304; Pall Corp.). The MBR was sterilized at 120°C for 15 min before cultivation. At the end of the culture period, cells were harvested in a presterilized bottle (20 L) and concentrated by centrifugation.

Schematic illustration of the MBR (15 L). The MBR was designed for pure monocultures of anaerobes. Growth medium and cell-filtering membranes (3.5 m2, 0.22 μm pore size, Al2O3) were externally fitted onto the MBR in the stainless steel housing. MBR, membrane bioreactor.
Measurement of live and HT cells
The numbers of live LM1004 (1 × 1011 colony-forming units per gram of freeze-dried lactobacilli) were quantified by counting cells on MRS plates using an automated cell counter (TC20; Bio-Rad Laboratories, Hercules, CA, USA). For both HT-LM1004 and MHT-LM1004, dead cells were counted with a hemocytometer under a microscope (BX53F; Olympus, Tokyo, Japan). Cell density during culturing in either the batch culture or MBR was analyzed by real-time measurement of optical density at 660 nm (G10S UV-Vis; Scinco Co., Gangnam, Korea).
Cell culture and sample treatment
YAC-1 cells, a mouse lymphoma cell line, and RAW 264.7 cells, a mouse macrophage cell line, were obtained from ATCC (Manassas, VA, USA; No. ATCC®TIB-160™ and ATCC®TIB-71™). YAC-1 cells, lymphocytes isolated from mouse spleen, and RAW 264.7 macrophages were cultured in RPMI 1640 and Dulbecco's modified Eagle's medium (DMEM) containing 10% FBS, penicillin, and streptomycin sulfate at 37°C and 5% CO2. Cells were treated with various concentrations (0.5–20 × 105 cells/mL) of HT-LM1004 or MHT-LM1004, LPS (1 μg/mL), and ConA (1 μg/mL) for various times.
Measurement of cell cytotoxicity
Cell viability was assessed with the MTT assay or the LDH cytotoxicity assay. RAW 264.7 mouse macrophage cells (1 × 106 cells) were seeded in 96-well plates containing 100 μL RPMI 1640 medium and 10% FBS and incubated overnight. HT-LM1004 (0.5, 1, 5, or 20 × 105 cells/mL) was added, followed by incubation for 24 h. MTT solution (5 mg/mL in PBS) was added for 4 h at 37°C and 5% CO2; the medium was then discarded, and formazan crystals were dissolved with 200 μL DMSO and the absorbance of each well at 550 nm was measured with a microplate reader (Varioskan; Thermo Fisher Scientific, Waltham, MA, USA).
Animals
Four-week-old female BALB/c mice (20 ± 2 g) were obtained from Samtako (Osan, Korea) and housed in a room with controlled temperature (22°C ± 2°C) and humidity (50% ± 5%) on a 12:12-h light/dark cycle with free access to food and water. The mice used in this study were handled in accordance with the Guidelines for the Care and Use of Laboratory Animals published by the U.S. National Institutes of Health (NIH Publication No. 85-23, 1996), and all experimental procedures were approved by the Committee on Ethics of Animal Experiments of International University of Korea (Ethics No. IUK-M-1710/01).
Preparation of the immunosuppressed mouse model
BALB/c mice were divided into seven groups (n = 8) each consisting of normal, CTX, CTX +0.1 mg/kg HT-LM1004, CTX +1 mg/kg HT-LM1004, CTX +10 mg/kg HT-LM1004, CTX +100 mg/kg HT-LM1004, and normal +100 mg/kg HT-LM100. Forty mice were injected intraperitoneally (i.p.) with CTX in sterile saline (60 mg/kg) for 2 consecutive days to establish immunosuppressed models. Another 16 mice (normal and normal +100 mg/kg HT-LM100 group) injected with normal saline served as the control group. CTX-immunosuppressed mice were randomly divided into five groups (n = 8 each) and HT-LM1004 in saline was administered intragastrically at 0.1, 1, 10, and 100 mg/kg once daily for 2 weeks. For the measurement of body and organ weights, mice were weighed on days 0 and 14. At the end of the experiment, the mice were sacrificed by injection of 200 mg/kg pentobarbital, and organs including the spleen were immediately removed and weighed. The immune organ index (%) was calculated according to the formula: index = organ weight (mg)/body weight (g).
Serum biochemistry
Alanine aminotransferase (ALT) and aspartate aminotransferase (AST) serum activities were measured to assess hepatotoxicity. ALT and AST activities were measured using spectrophotometric diagnostic kits (Sigma Chemical Co). Briefly, the samples were centrifuged at 1000 g for 10 min within 1 h after collection. The sera were stored in the −80°C freezer until they were analyzed. Enzyme activities of ALT and AST in blood serum were evaluated with an autoanalyzer (Varioskan; Thermo Electron Co.).
Assay for macrophage phagocytosis
The phagocytosis assay was performed as previously described. 29 Briefly, 100 μL fluorescein-5-isothiocyanate (FITC)-labeled Escherichia coli (Molecular Probes, Eugene, OR, USA) was added to the wells of a 96-well plate containing RAW 264.7 macrophages. The plate was incubated for various times at 37°C in a humidified atmosphere of 5% CO2. Extracellular fluorescence was quenched by adding 100 mL Trypan blue. After 1 min, FITC-labeled bacteria that had not been phagocytosed by macrophages were washed away, and the macrophages were rinsed twice with PBS and then lysed with lysis buffer (10 mM Tris-HCl [pH 7.5], 130 mM NaCl, 1% Triton X-100, 10 mM Na2HPO4, and 10 mM Na4P2O7). The relative fluorescence intensity of bacteria inside the macrophages was determined at excitation and emission wavelengths of 480 and 520 nm, respectively, using a microplate reader. The relative phagocytic activity was calculated as the percent fluorescence intensity of sample-supplemented versus unsupplemented (control) FITC-labeled bacteria.
Assay of NK cell activity
The spleen was aseptically removed from each mouse and ground into a single-cell suspension using sterile gauze and washed three times with RPMI 1640 medium. Cells were centrifuged at 1000 g for 10 min at room temperature, and 100 μL spleen cell suspensions (1 × 106 cells) were seeded in a 96-well cell culture plate with 1 × 105 NK-sensitive YAC-1 cells (Korean Cell Line Bank, Seoul National University, Korea) at an effector cell:target cell ratio of 10:1, while 100 μL RPMI 1640 medium was used as a control. After 4 h incubation at 37°C (5% CO2), the plate was centrifuged at 800 g for 5 min and the culture supernatant (100 μL per well) was mixed with LDH solution (Promega, Madison, WI, USA); the absorbance of each well was measured at 490 nm. NK cell cytotoxicity was calculated using the following formula: cytotoxicity (%) = ([experimental release − spontaneous release]/[maximum release − spontaneous release]) × 100.
Lymphocyte proliferation assay
Cell cytotoxicity was examined with a water-soluble tetrazolium (WST)-1 assay kit according to the manufacturer's instructions. Briefly, mouse spleen cells (1 × 106) were seeded in 96-well plates in DMEM with 10% FBS. After mixing with 10 μL RPMI 1640 medium (negative control) or 10 μL of 200 μg/mL ConA (positive control), the cells were incubated at 37°C and 5% CO2 for 48 h. The supernatant was removed, and the cells were used in the WST-1 assay. Relative cytotoxicity was quantified by measuring the absorption at 550 nm with a microplate reader.
Measurement of cytokine levels
Cells were cultured for 6 and 24 h at a density of 1 × 106 cells per well in 48-well plates. At the end of the experiment, blood samples were collected from the caudal vein of mice and centrifuged at 3000 g for 10 min at room temperature. The supernatant was collected and stored at −80°C. IFN-γ, IL-2, IL-6, IL-12p70, and TNF-α levels were quantified using ELISA kits.
Statistical analysis
Results are expressed as mean ± standard deviation (SD) of triplicate experiments. Data from the animal study are expressed as mean ± SD (n = 8). Mean differences were evaluated by analysis of variance followed by Dunnett's post hoc test, and P-values <.05 were considered statistically significant.
Results
L. plantarum LM1004 culture in the MBR media
L. plantarum LM1004 was grown in the MBR media for large-scale production (Table 1). L. plantarum LM1004 grew with no feeding for 12 h in batch cultures, but in the MBR, the cells grew for 19 h with a fresh medium feeding rate of 0–0.99/h. At the end of cultivation, the cell mass produced was 15.2 g/L by batch culture; a 11.2-time higher mass (170.4 g/L) was obtained with the MBR. During culture in the MBR, the spent medium containing harmful wastes that could inhibit growth was continuously removed through a cell-filtering membrane, thereby increasing the final cell density. Indeed, the cell production rate (g/h·L) was increased from 1.27 in batch culture to 9.47 in the MBR (7.46 times higher). On the contrary, the mean cell size was smaller in the MBR (1.08–1.12 μm) than in batch culture (1.74–2.58 μm) (Fig. 2). Heat treatment (110°C, 20 min) after MBR culture further reduced cell size to 0.82–0.9 μm. The size distribution of cells cultured in the MBR showed a sharper peak than those grown in the STR.

Size distribution of cells cultured in the STR and MBR.
Comparison of Cell Productivity Between Batch and Membrane Bioreactor Cultures of Lactobacillus plantarum HT-LM1004
L. plantarum LM1004 was cultivated in a stirred tank reactor (300-L working volume/500-L pilot scale fermenter) to generate HT-LM1004 or in an MBR (15 L working volume/15 L MBR) to generate micronized/HT-LM1004. The culture pH was maintained between 5.8 and 6.0. Data are expressed as an average (n = 3) and the error rate was ±0.05.
CFU, colony-forming unit; HT, heat-treated; MBR, membrane bioreactor.
HT-LM1004 increases phagocytic activity and NO production in RAW 264.7 macrophages
To obtain a suitable concentration range to investigate the effects of HT-LM1004 on cell viability in RAW 264.7 cells, we treated cells with HT-LM1004 concentrations ranging from 0.5, 1, 5, or 20 × 105 cells/mL for 24 h. The toxicity and cell viability of HT-LM1004 to RAW 264.7 cells were evaluated with the MTT (Fig. 3A) and LDH (Fig. 3B) assays. HT-LM1004 had no effect on cell viability even at concentrations up to 20 × 105 cells/mL. Therefore, we treated cells with HT-LM1004 concentrations ranging from 0.5 to 5 × 105 cells/mL in subsequent experiments.
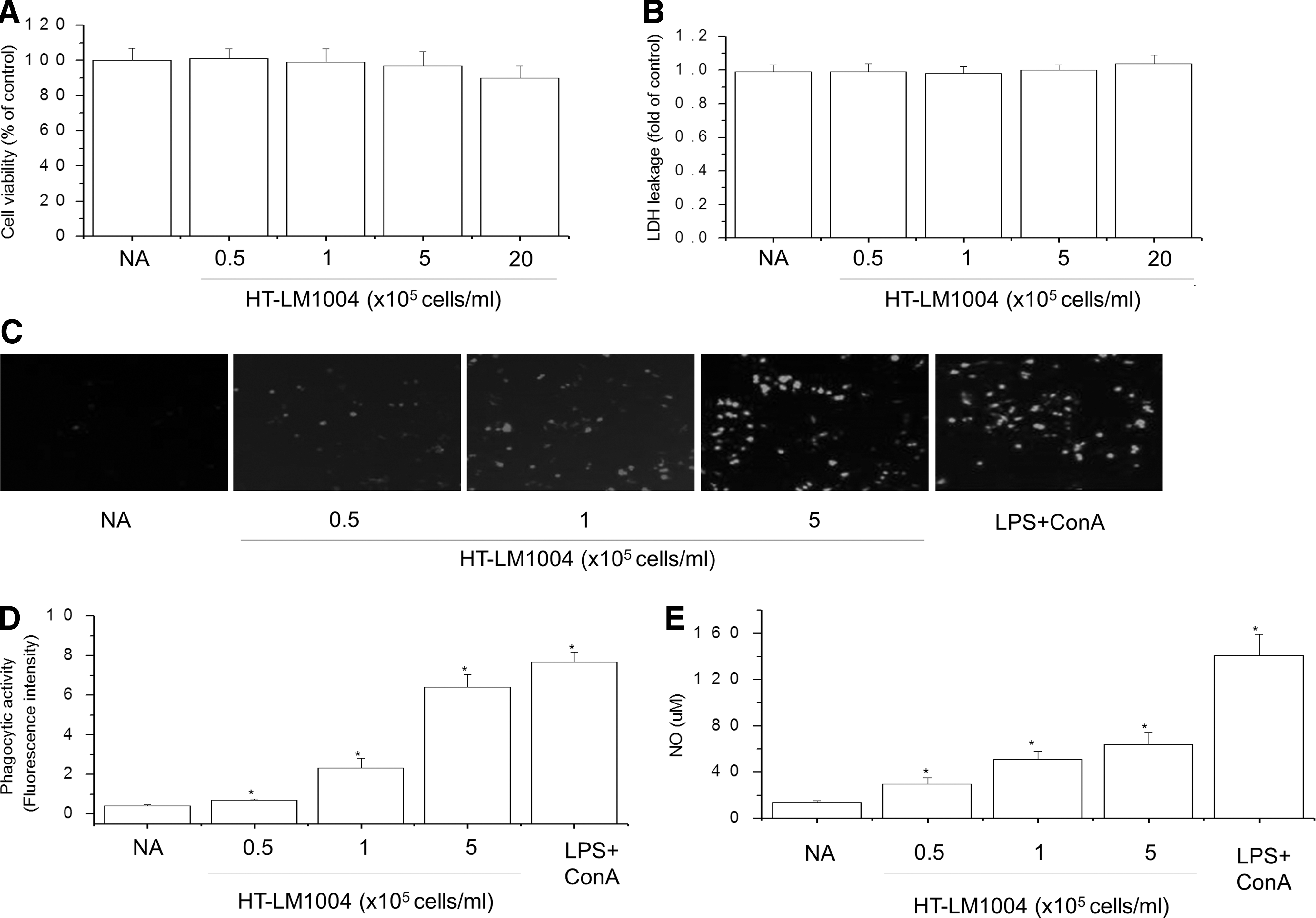
Effects of HT-LM1004 on phagocytosis and NO production in mouse splenocytes.
To determine the effects of HT-LM1004 on the phagocytic activity of RAW 264.7 macrophages, we compared the uptake of FITC-labeled E. coli between HT-LM1004-treated and untreated macrophages. The stimulatory effect of HT-LM1004 on macrophage phagocytosis was evaluated based on the internalization of FITC-labeled E. coli cells, with mean fluorescence intensity estimated from fluorescence spectrometer reading. Macrophage phagocytosis was increased in a dose-dependent manner by HT-LM1004 treatment (Fig. 3D). E. coli uptake was confirmed by confocal laser scanning microscopy (Fig. 3C). These results demonstrated that HT-LM1004 enhances phagocytosis by macrophages, which is one of the major functions of these immune cells.
To further investigate whether HT-LM1004 activates macrophages, we measured NO production in mouse splenocytes incubated with HT-LM1004 for 24 h by measuring NO concentrations in the culture supernatant by the Griess reaction. HT-LM1004 (0.5–5 × 105 cells/mL) increased NO levels in a concentration-dependent manner (Fig. 3E), but was not toxic to the RAW 264.7 cells in the range of tested concentrations, as determined with the MTT assay (data not shown).
HT-LM1004 increases TNF-α, IFN-γ, IL-2, and IL-12 levels in mouse splenocytes
Activated macrophages release proinflammatory cytokines such as TNF-α, IFN-γ, IL-2, and IL-12p70, which are potent immune modulators. 15 To examine the effect of HT-LM1004 on cytokine production, we measured TNF-α, IFN-γ, IL-2, and IL-12p70 levels in splenocytes culture supernatant with ELISA. HT-LM1004 (0.5, 1, or 5 × 105 cells/mL) stimulated the production of all of the tested cytokines in a concentration-dependent manner (Fig. 4A–D), with LPS and ConA serving as positive controls. These data demonstrate that HT-LM1004 is a potent inducer of cytokine secretion.

Effects of HT-LM1004 on TNF-α, IFN-γ, IL-12, and IL-12p70 production by mouse splenocytes.
Effects of HT-LM1004 on cytokine levels, NK cell activity, and splenocyte proliferation in a mouse model of CTX-induced immunosuppression
To investigate the immune-enhancing effects of HT-LM1004 under physiological conditions, we carried out an in vivo study. CTX suppresses the immune system by depleting lymphoid cells, thereby suppressing the hematopoietic function of the medulla and causing leukopenia. HT-LM1004 was orally administered daily to CTX immune-suppressed mice, which were injected (i.p.) with CTX or HT-LM1004 twice over 14 days. The spleen was examined for morphological changes to assess the effects of HT-LM1004 on immune organs, and body and liver weight were monitored throughout the study. CTX treatment did not cause any changes in body or liver weight compared with mice with no CTX treatment. Moreover, the spleen indices of CTX-treated mice (60 mg/kg per day, i.p.) were reduced compared with untreated controls. On the contrary, the spleen indices of CTX+HT-LM1004-treated mice (0.1, 1, 10, or 100 mg/kg/day, oral administration) were dose-dependently increased relative to the CTX-only group (Table 2). The effects of HT-LM1004 on the CTX-induced elevation of serum ALT and AST activities are shown in Table 3. HT-LM1004 prevented the CTX-induced elevation in the ALT and AST serum levels (Table 3).
Regulatory Effects of HT-LM1004 on Body Weight and Organ Indices
Female Balb/C mice were administered 0.1–100 mg/kg HT-LM1004 (LM) for 14 days. CTX was intraperitoneally injected as a single dose of 60 mg/kg on day 13 before sacrifice. Data are expressed as mean ± SD (n = 8).
P < .05 versus normal group; * P < .05 versus CTX-treated group.
CTX, cyclophosphamide; SD, standard deviation.
Effects of HT-LM1004 on Alanine Aminotransferase and Aspartate Aminotransferase Activity in Cyclophosphamide-Treated Mice
Female Balb/C mice were administered 0.1–100 mg/kg HT-LM1004 (LM) for 14 days. CTX was intraperitoneally injected as a single dose of 60 mg/kg on day 13 before sacrifice. Data are expressed as mean ± SD (n = 8).
P < .05 versus normal group; * P < .05 versus CTX-treated group.
ALT, alanine aminotransferase; AST, aspartate aminotransferase.
We also measured serum levels of TNF-α, IL-2, IL-12p70, and IFN-γ in mice with CTX-induced immunosuppression treated with different concentrations of HT-LM1004 and found that the levels of all of these cytokines were reduced compared with those in mice treated with CTX only (Fig. 5A–D). These results suggest that HT-LM1004 restores cytokine production that was abolished by CTX. NK cells are the first line of defense against tumor cells and cells infected with virus. 30 Here we found that CTX inhibited NK cell activity (Fig. 5E) as well as splenocyte proliferation (Fig. 5F); these effects were reversed in a dose-dependent manner by HT-LM1004 treatment compared with treatment with CTX only.

Effects of HT-LM1004 on inflammatory cytokine levels, NK cell activity, and splenocyte proliferation in CTX-treated mice.
Comparison of the immune-enhancing effects of MHT-LM1004 and HT-LM1004
Micronized and heat-treated LM1004 (MHT-LM1004) produced in the MBR and HT-LM1004 produced in batch cultures were compared in terms of the production of cytokines TNF-α, IFN-γ, IL-6, and IL-12p70 (Fig. 6). Cytokine release, especially of IL-12 p70 in splenocytes, was strongly stimulated in a dose-dependent manner by treatment with MHT-LM1004. IL-12 p70 is associated with IFN-γ production by macrophages and activation of phagocytic activity by NK cells. In addition, MHT-LM1004 treatment in splenocytes was found to have a higher NK cell toxicity to YAC cells compared with HT-LM1004 treatment (Fig. 6F).

Comparison of immune-enhancing effects of MHT-LM1004 and HT-LM1004 using mouse splenocytes. Mouse splenocytes (1 × 106 cells per well) were cultured for 3 h (TNF-α) or 24 h (IFN-γ, IL-12p70, and IL-6) in medium without or with 0.5–5 × 105 cells/mL HT-LM1004, MHT-LM1004, or LPS+ConA (1 μg/mL).
Discussion
The immunostimulatory potential of probiotics makes them promising prophylactic and therapeutic agents for immune diseases. 10 Several Lactobacillus strains have been proposed to have beneficial health effects, including enhancement of the immune response. 13 In addition, HT probiotics have been reported to promote host defense against pathogenic infection and stimulate innate immunity. 31 In the present study, we evaluated the immunostimulatory activity of HT-LM1004 isolated from Korean kimchi (fermented Napa cabbage) using mice splenocytes and a mouse model of CTX-induced immunosuppression.
The immunostimulatory mechanisms of Lactobacillus include the modulation of T cell effector subsets to support humoral immunity in conjunction with macrophages and dendritic cells.
32
Innate immunity is the first line of defense in the immune system. Macrophages play key roles in host defense mechanisms, and many immune regulators potentiate innate immune responses by activating phagocytosis in macrophages in response to invading pathogens.
32
In the present study, we found that HT-LM1004 enhanced the phagocytic activity of macrophages. During phagocytosis, activated macrophages release NO and cytokines such as TNF-α, IFN-γ, IL-2, and IL-12p70. NO, which is synthesized from
Immunosuppression induced by chemotherapy limits the therapeutic efficacy of cancer treatment. As such, there is a need for new strategies to prevent or reduce the toxicity of chemotherapeutic agents at the level of hematopoiesis, which could improve patient prognosis. CTX is the most widely used alkylating agent in chemotherapy; in addition to its clinical effect, it can also inhibit cell proliferation and reduce the levels of inflammatory cytokines. 4 We examined the immune-enhancing effect of HT-LM1004 in a mouse model of CTX-induced immunosuppression. CTX reduced spleen indices, which reflect nonspecific immunity, 34 in mice. Several studies have reported that immune-potentiating agents can restore spleen weight in animals with CTX-induced immunosuppression. 25,34 Similarly, we found that HT-LM1004 orally administered for 14 consecutive days to CTX-treated mice increased spleen indices compared with mice treated with CTX only, suggesting that HT-LM1004 has protective effects in lymphoid tissues and can counter immunosuppression.
Previous studies have shown that LAB increase lymphocyte proliferation in response to mitogens in mice, and stimulate the production of proinflammatory cytokines such as TNF-α, IFN-γ, IL-1β, and IL-6 by immune cells. 22,23 Our results demonstrated that HT-LM1004 induced the secretion of TNF-α, IFN-γ, IL-6, and IL-12p70 in mice with CTX-induced immunosuppression. Furthermore, HT-LM1004 administration increased splenocyte proliferation in these mice. NK cells are critical for host defense since they exert cytotoxic activity against viruses, bacteria, and even cancer cells. 35 We found that although CTX suppressed NK cell activity, this was alleviated in a dose-dependent manner by LM1004. Thus, HT-LM1004 can enhance cell-mediated and humoral immunity under conditions of immunosuppression. Previous studies have shown that the increases in NO and cytokines are associated with the NF-κB and MAPK pathways. The transcription factor NF-κB and activated MAPK pathways are the key regulators of the expression of the genes encoding macrophage-related cytokines. We need to further experiment with the effects of LM1004 on NF-κB and MAP kinase activity. 36,37
We also found that MHT-LM1004 produced in the MBR activated NK cell cytotoxicity against NK-sensitive YAC-1 cells to a greater extent compared with HT-LM1004 produced in batch culture and live HT-LM1004. Thus, both MHT-LM1004 and HT-LM1004 can successfully enhance the host immune response. Furthermore, MHT-LM1004 was nontoxic in vivo in the oral toxicity test using rats or in the single oral dose toxicity test using beagle dogs (data not shown), which were performed at GLP Laboratory in accordance with the Organization for Economic Co-operation and Development guidelines (Korea Testing & Research Institute, Gwacheon, Korea). In both experiments, there were no signs of inflammation or tissue necrosis and no mortality, even when administered at a high dose of 5000 mg/kg body weight.
In summary, we showed that the immune-stimulatory effect of HT-LM1004 was greater when it was manufactured in the MBR than in batch culture, indicating that the effects of probiotics can vary depending on the production method. HT-LM1004 stimulated phagocytosis by NK cells and the production of immune modulators such as NO and proinflammatory cytokines. Importantly, HT-LM1004 improved spleen indices in a mouse model of CTX-induced immunosuppression. Our findings provide a basis for the use of MHT-LM1004 or HT-LM1004 as agents that can counter the immunosuppressive effects of chemotherapeutic drugs used for cancer, and thereby improve treatment outcome.
Footnotes
Acknowledgments
This work was supported by a 2017 research grant from LactoMason Co., Ltd., and also financially supported by the Ministry of Trade, Industry, and Energy, Korea, under the “Regional Specialized Industry Development Program” supervised by the Korea Institute for Advancement of Technology (R0005616).
Author Disclosure Statement
No competing financial interests exist.