
Abstract
Moringa oleifera is a natural plant with high nutritional and pharmacological value. Leaves of M. oleifera contain a variety of active substances. In our previous research, we had obtained a polysaccharide separated from M. oleifera leaf, namely MOs-2-a (1.35 × 104 Da). In this study, this polysaccharide was administrated daily to 6 week-old ICR mice for 4 weeks. Then, the body weight, immunity, intestinal digestion, and intestinal microenvironment of Institute of Cancer Research (ICR) mice were investigated. After 4 weeks of feeding intervention with the polysaccharide, the immune and intestinal digestive ability of the ICR mice were significant as shown by the organ index, digestive enzymes, and reduction of serum tumor necrosis factor-alpha and diamine oxidase levels. The polysaccharide could regulate the microbial composition of the intestinal tract in mice by increasing the bacteria that have been reported for antiobesity effects, short chain fatty acid production, and lactic acid production. These findings indicate that the polysaccharide of M. oleifera leaf might be a promising prebiotic that exhibits health promotion effects.
INTRODUCTION
The importance of gut microbiota in human health has been revealed by numerous studies in recent years. 1 –3 Many chronic diseases and symptoms such as type 2 diabetes, gout, and cardiovascular system disease are proved to be highly correlated with gut microbiota composition and function. 4 –6 For the treatment of these chronic diseases, in addition to insisting on correct long-term medication, proper dietary habits should be adopted to improve the balance of nutritional intake. Many studies have shown that dietary prebiotics, such as plant polyphenol, polysaccharide or oligosaccharide, and plant fiber can selectively stimulate the growth of the beneficial bacteria and adjust the body's physiological functions. 7,8
Oligo- and polysaccharides in plants have been found to exert many important gut health functionalities including prebiotic functions. 9 –11 Inulin and galacto-oligosaccharides could selectively promote the growth of beneficial bacteria in the human colon and regulate the inflammatory markers in adult patients. 12 Most of these oligo- and polysaccharides cannot be digested and absorbed in the upper gastrointestinal tract. However, they can be fermented or metabolized by the intestinal bacteria and stimulate the growth of bacterial groups that produce metabolites such as short chain fatty acids (SCFAs) that are helpful for host physiology, particularly gastrointestinal health. 13 Furthermore, some prebiotics such as lentinan and Ganoderma lucidum polysaccharides have demonstrated immunity enhancement capacity. 2,14
Moringa oleifera is native to northern India, and belongs to the family Moringaceae, a single family of shrubs. The tree is distributed in many tropical and subtropical countries and is now widely cultivated in the south China. 15 M. oleifera is rich in nutrients and has many pharmacological activities. 16 –18 M. oleifera leaves are rich in protein, vitamins, calcium, and iron, and fresh M. oleifera leaves contain higher concentration of calcium, potassium, iron, vitamins A and C, and protein than common food and fruits such as milk, yoghurt, oranges, and bananas. 19 The M. oleifera seed contains phytochemicals such as alkaloids, flavonoids, and phenolic glycosides that are reported to exhibit hypoglycemic and antiphlogistic activities. 20 –22 The extract of M. oleifera leaf was shown to have antioxidant and antidiabetic activities both in vitro and in vivo, which could be used as food by diabetic patients to protect them against oxidative damage. 23 Besides, previous studies have found that M. oleifera leaf powder-supplemented diet could affect the growth of animals. 24 In our previous research, we had obtained a polysaccharide separated from M. oleifera leaf, namely MOs-2-a (1.35 × 104 Da) that is composed by mannose, rhamnose, glucose, and galactose with molar ratio of 0.49:3.65:0.63:1.27. 25 In this study, this polysaccharide was administered daily to 6-week-old Institute of Cancer Research (ICR) mice for 28 days. The beneficial effect of MOs-2-a on the intestinal microecology was assessed to reveal the potential prebiotic effects of MOs-2-a.
MATERIALS AND METHODS
Preparation of MOs-2-a
The M. oleifera leaves were obtained from Yunnan Biotechnology Co., Ltd (Yunnan, China). The preparation and characteristics of MOs-2-a were performed as previously described. 25 In brief, defatted samples were extracted by 80% ethanol and the residues were extracted by deionized (DI) water for 2 h. The extracting solution was then passed over a microporous resin column and the H2O effluent was concentrated in vacuum to a proper volume (500 mL). Ethanol was added slowly into the concentrated solution until the alcohol content reached 80% and was then stored at 4°C overnight. The solution was centrifuged and sediment was dried to obtain crude polysaccharide. The crude polysaccharide aqueous solution was then separated by diethylaminoethylcellulose-52 column chromatography and the subfraction was further fractionated on a Glucan-200 gel column and eluted with 0.1 mol/L NaCl solution and then dialyzed to obtain MOs-2-a.
Animals and treatments
Thirty male ICR mice (aged 6 weeks, 18–22 g) were obtained from the Laboratory Animal Center in Nanjing University of Chinese Medicine. All the animals were housed with a 12 h light/dark cycle at 22°C and 55% ± 5% relative humidity and had free access to food and water. Animal experiment protocols in this study were carried out according to the guidelines of the Animal Care Committee of Nanjing University of Chinese Medicine, the ethics number is SCXK2016-0010.
Thirty ICR mice were randomly divided into three groups (n = 10). Subsequently, they were treated with MOs-2-a (50 mg/kg for MOs-2-a-H group and 10 mg/kg for MOs-2-a-L group) or DI water for 28 consecutive days.
The body weight of each mouse was measured at days 0, 7, 14, and 28. The blood samples were collected by carotid puncture from each mouse before the laparotomy. Serum samples were obtained by centrifugation of the collected blood (1789 g for 10 min).
The duodenum, ileum, and colon of each mouse were dissected, a portion of the tissue was placed in bottles with 4% neutral-buffered paraformaldehyde in physiological saline for pathological evaluation. Other parts were stored at −80°C for measurement of enzyme activity of intestinal tissue. Mice were sacrificed with pentobarbital sodium.
Pathological section, staining, and pathological evaluation of intestinal tissue
About 2 cm of duodenum, ileum, and colon was cut from the mice, respectively. These intestinal tissues were formalin fixed and embedded in paraffin. Sections (4-μm thick) were stained with hematoxylin-eosin (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) and examined using a light microscope (Olympus Medical Systems Corp., Tokyo, Japan). The villus height, crypt depth, and mucosal thickness of each segment were measured, and the ratio of villus height and crypt depth (V/C) value was calculated by ImagePro.
Measurement of biochemical indicator
Serum was obtained through centrifugation (1006 g, 10 min, 4°C) for the detection of biochemical indicator. Serum levels of diamine oxidase (DAO),
Determination of the enzyme activity in the intestine
The intestinal segment was homogenized in ice-cold physiological saline. The homogenate was collected and centrifuged (4025 g, 10 min, 4°C) to absorb the supernatant. The activity of amylase (Amy), trypsin (Try), lipase (Lip), and intestinal alkaline phosphatase (ALP) in supernatant was determined using the kits following the manufacturers' instructions.
Sample collection and fecal DNA extraction
Fecal samples were collected from each individual mouse at the end of the experiment and stored at −80°C for later DNA preparation. DNA extraction was performed using an E.Z.N.A.® Soil DNA kit (Omega Bio-Tek, Georgia State, USA). The concentration of bacterial DNA was measured using Nanodrop 2000 (Thermo Scientific, USA).
16S ribosomal RNA gene sequencing
The V3–V4 region of the bacteria's 16S ribosomal RNA (rRNA) gene was amplified by polymerase chain reaction with barcode-indexed primers (338F and 806R), using FastPfu polymerase. Amplicons were then purified by gel extraction (AxyPrep DNA GelExtraction Kit; Axygen Biosciences, Union City, CA, USA) and were quantified using QuantiFluor-ST (Promega, USA). The purified amplicons were pooled in equimolar concentrations and paired-end sequencing was performed using an Illumina MiSeq PE 30 instrument (Illumina, San Diego, CA, USA).
Microbial analysis
The 16S rRNA sequencing data were processed using the Trimmomatic software. Sequencing reads were demultiplexed and filtered. Operational taxonomic units were picked at 97% similarity cutoff, and the identified taxonomy was then aligned using the Greengenes database (V.13.8). Chimeric sequences were identified and deleted. Ribosomal Database Project (RDP) classifier (
Statistical analysis
Results were presented as mean ± standard deviation. Experiments were conducted separately at least three times. Data were analyzed by one-way analysis of variance (ANOVA). Tukey's multiple comparison test was made for multiple comparisons among all groups if overall P < .05. P < .05 was considered statistically significant. R (pheatmap package) was used to perform a Spearman correlation analysis of the factors associated with intestinal mucosal integrity and the diversity indices of bacterial communities. The Spearman correlation coefficient of the top 30 abundant bacteria at family level and the factors associated with intestinal mucosal integrity were calculated, and displayed on the heat map.
RESULTS
MOs-2-a altered body weight and viscera index of ICR mice
After 7 days of polysaccharides administration, the body weight in the MOs-2-a group was increased compared with control mice (Fig. 1A). During the experimental period, the mice treated with MOs-2-a kept growing, and at the end of this research, body weight in the MOs-2-a group was significantly higher than in mice fed standard chow (P < .05).
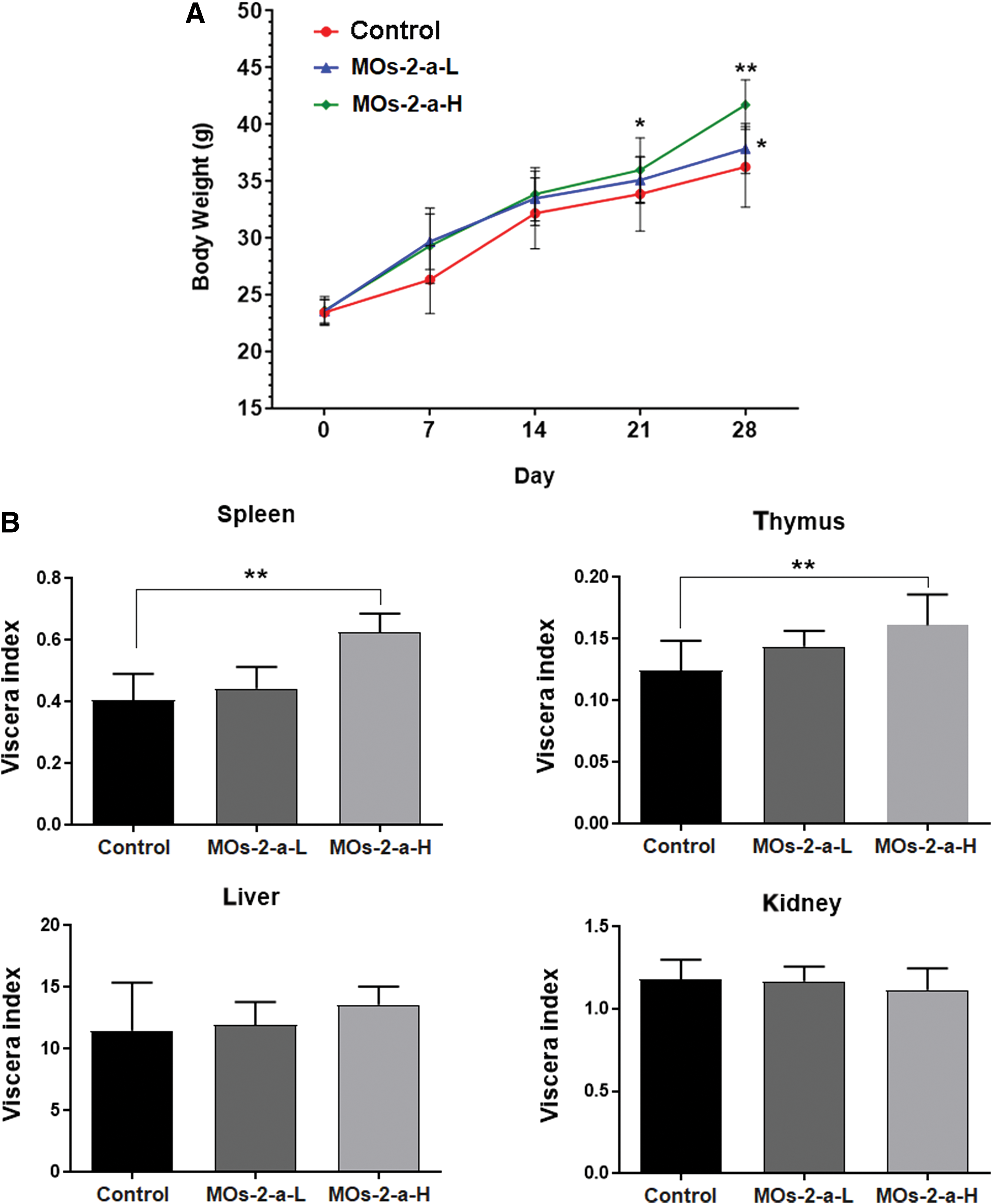
Body weight
MOs-2-a treatment could significantly increase the viscera index of spleen and thymus (Fig. 1B). As the main immune organ in the body, the increase of spleen and thymus index indicates that MOs-2-a could improve the differentiation of immune organs and enhance the immune function.
Changes in intestinal morphology
The integrity of intestinal tissue is a prerequisite for maintaining favorable intestinal microenvironment and ensuring good digestive activity. Serum ET, TNF-α, DAO, and
Effects of MLP Treatment on Biochemical Parameters in Serum of ICR Mice
Values are presented as mean ± SD, n = 4, * P < .05, ** P < .01 versus control.
DAO, diamine oxidase;

Effect of MOs-2-a on intestinal morphology.
Changes in digestive ability
MOs-2-a administration could enhance the activity of various digestive enzymes. The Amy activity in the MOs-2-a group was much higher than the control group (28.5% higher of MOs-2-a-L group, P < .01; 54.9% higher of MOs-2-a-H group, P < .01). Correspondingly, the activities of ALP (65.3% higher than control group, P < .01), Try (38.9% higher than control group, P < .05), and Lip (29.9% higher than control group) were improved by MOs-2-a treatment (Fig. 3).
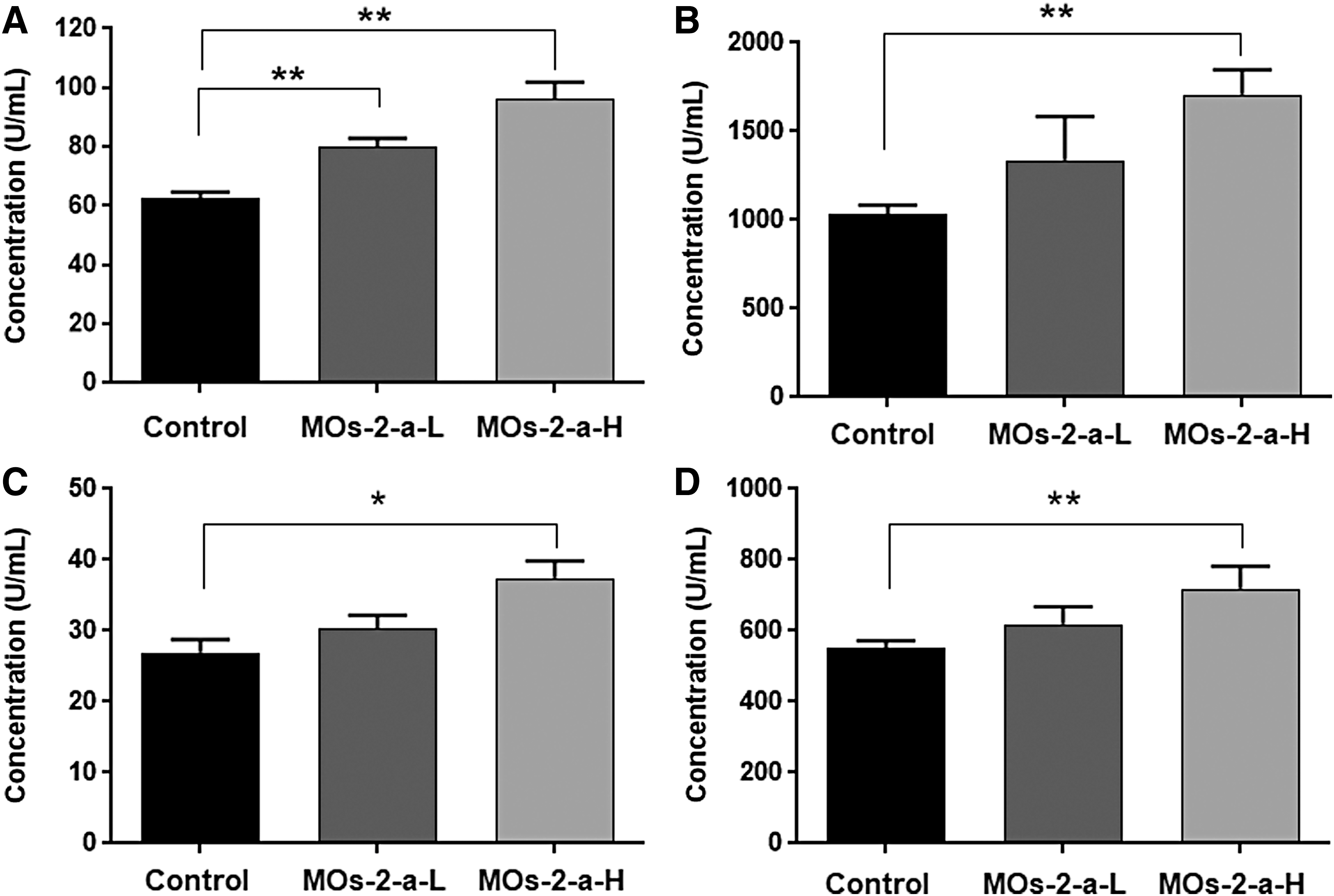
Effect of MOs-2-a on digestive enzyme activity.
Physical and chemical properties of feces
The nitrogen content and pH value of the feces are shown in Table 2. Compared with the control group, MOs-2-a treatment could significantly reduce the nitrogen content of feces (P < .01) and keep the pH at faintly acid.
Physical and Chemical Properties of Feces
Values are presented as mean ± SD, n = 10, * P < .05, ** P < .01 versus control.
MOs-2-a induces core changes in composition and diversity of gut microbiota
Alfa diversity refers to the diversity of a region or ecosystem that is an ecological indicator of how many taxonomic groups are present within each sample. 26 Compared with control group, MOs-2-a–treated groups were significantly reduced in alpha diversity (Fig. 4A), using either the sobs diversity metric or the Shannon entropy metric (Student's t-test, all P values <.05).

MLP induces core changes in composition and diversity of gut microbiota.
After feeding with MOs-2-a, gut microbiota composition was restored. Analysis of average relative percentage abundance and hierarchal clusters at phylum level showed that MOs-2-a treatment could upregulate Firmicutes and Proteobacteria, whereas downregulate Bacteroidetes (Fig. 4B). PERMANOVA test was assessed at genus level, and all the groups exhibited significantly different microbial compositions (P < .01). Furthermore, principal co-ordinates analysis based on weighted UniFrac distances and unweighted UniFrac distances (Fig. 5A, B) between different samples revealed an obvious difference between MOs-2-a treatment group and control group, which is further proved by non-metric multidimensional scaling analysis based on Bray–Curtis similarity distance (Fig. 5C).
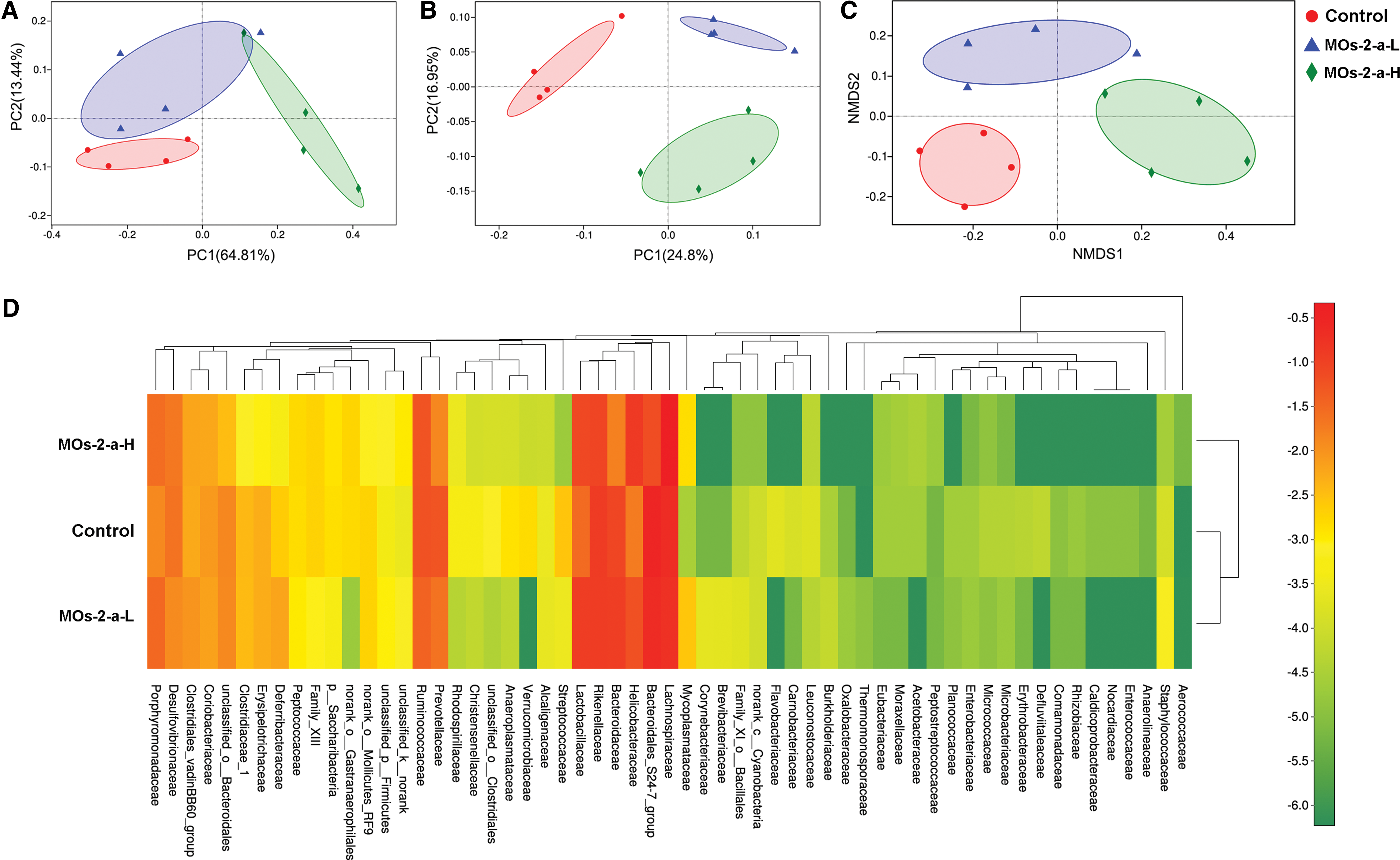
GM clustering of three experimental groups displayed by PCoA plots of the unweighted UniFrac matrix
The heat map graphically showed that at the family level, Lachnospiraceae, Bacteroidaceae, Helicobacteraceae, Rikenellaceae, and Lactobacillaceae composed the dominant bacterial community and the result of cluster analysis shows that the microbial composition between control group and MOs-2-a-L group was similar but there was significant difference between control group and MOs-2-a-H group (Fig. 5D).
MOs-2-a induces major changes of taxa
The taxa with abundance >0.01% were isolated and analyzed among the detected genus (Fig. 6A). Bacteroidales-S24-7, Bacteroides, Lachnospiraceae-NK4A136, and Rikenellaceae-RC9 were dominating in the control group. MOs-2-a treatment modulated the abundance of these genera. Bacteroidales-S24, Bacteroides, and Rikenellaceae-RC9 were reduced in MOs-2-a group, and an increase of Lachnospiraceae-NK4A136 was observed in MOs-2-a-H group. Furthermore, some beneficial bacteria such as Lactobacillus, Lachnospiraceae, Rikenella, and Roseburia were markedly enriched in the MOs-2-a group (Fig. 6B). All the detected genera were analyzed for linear discriminant analysis score to determine the significantly abundant genus in each group. The result showed that Parabacteroides was most prevalent in the MOs-2-a-L group. MOs-2-a-H group was enriched in Lachnospiraceae and Eubacterium-xylanophilum.

Correlation analysis between biochemical factors and bacteria
The correlation heat map showed that the relationship between bacterial phyla and biochemical factors was different. As for the factors associated with digestion, Lachnospiraceae, Helicobacteraceae, Deferribacteraceae, Lactobacillus, Helicobacter, and Peptostreptococcaceae showed a significant positive correlation, whereas for the factors associated with intestinal mucosal integrity, these bacteria showed significant negative correlations (Fig. 7A, B).
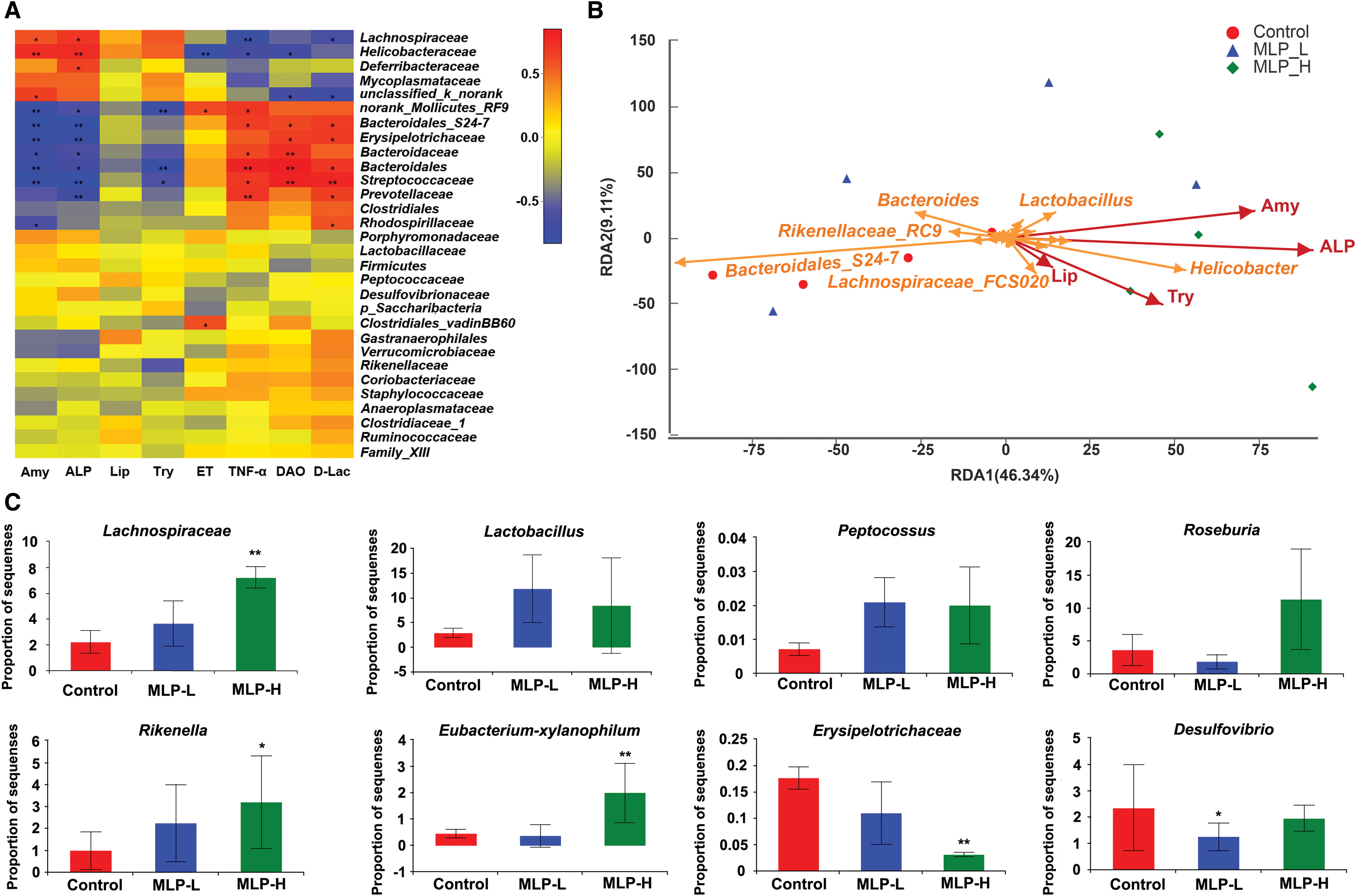
The influence of MOs-2-a on beneficial and pathogenic bacterial
The abundance of beneficial bacteria and pathogenic bacteria among three groups was compared (Fig. 7C). We found that the abundance of pathogenic bacteria such as Erysipelotrichaceae and Desulfovibrio was decreased in MOs-2-a–treated mice. Moreover, an evident increase of beneficial bacteria was observed in the MOs-2-a group compared with the control group. The increase was related to the expansion of Lachnospiraceae, Eubacterium-xylanophilum, Lactobacillus, Peptococcus Roseburia, and Rikenella. Some of these categories were highly related to the SCFAs production and digestion. 27,28
DISCUSSION
M. oleifera leaf has long been used in Asian and African countries as a food supplement because of its high-nutritional value. 29,30 Although M. oleifera leaf has been previously studied for its nutrition and medicine activities, 23,31,32 however, for the first time, in this study, we evaluated the beneficial effect on the gut microecology by MOs-2-a in mice, and the results indicated that MOs-2-a exhibited promising probiotic effects by regulating the gut microecology.
The intestinal integrity is very important for normal physiological activities. DAO is an important indicator of the integrity of intestinal epithelial cells and intestinal mucosal barrier. In the early stage of intestinal injury, the level of DAO in serum increases rapidly. Therefore, it is commonly used to detect intestinal barrier function.
33
Many kinds of bacteria in the gut can produce
The integrity of the structure and function of the small intestine and the activities of digestive enzymes are the prerequisite for the digestion and absorption of nutrients. The increase of villus height and mucosal thickness can enlarge the surface area of villus and increase the absorption of nutrients. 35 Paneth cells at the bottom of crypt play a key role in lipid digestion. The depth of crypt determines the rate of mitosis of intestinal villi to produce epithelial cells, which reflects the rate of cell formation, whereas the shallowness of crypt indicates the increase of cell maturation rate and the enhancement of secretory function. The increase of V/C value indicates the enhancement of digestive function, conversely, the decrease of V/C value indicates compromised performance of digestion and nutrients absorption. 36,37 ALP is a characteristic enzyme in digestion and absorption of small intestine. It participates in the absorption of many substances such as lipids, vitamins, calcium, and amino acids. The activity of ALP reflects the strength of digestion and absorption in small intestine. 38 Amy, Try, and Lip are related to the digestion of starch, protein, and fat, respectively. Their activity also directly reflects the function of digestion. 39 Our results found that MOs-2-a could significantly increase the thickness of mucous membrane and the value of V/C. These results suggested that feeding with MOs-2-a can improve the tissue surface area of the ileum as well as the activities of various digestive enzymes to degrade starch, protein, and lipid in food.
The gut microbiota is closely related to the various physiological activity states of the human body, including the digestion of food and the absorption of nutrients. Our results revealed that MOs-2-a also changed the composition of gut microbiota. Lachnospiraceae, Eubacterium-xylanophilum, and Peptococcus are important phyla of normal gut flora that participate in the synthesis of vitamins and proteins in the intestine, assist digestion and absorption, and stimulate immune function. 40 –42 In the MOs-2-a group, we found significant enrichment of the abundance of these bacteria. In addition, Lactobacillus can promote the secretion of digestive enzymes. Its metabolites produced can inhibit the growth of pathogenic bacteria in the intestinal tract, maintain the integrity of the intestinal mucosal barrier, and enhance the immunity of the body by activating the function of the intestinal autoimmune system. Therefore, Lactobacillus is beneficial bacteria in the intestine and plays an important role in maintaining intestinal health. MOs-2-a could also improve the Lactobacillus level. In contrast, MOs-2-a treatment can reduce the number of pathogenic bacteria in the intestinal tract and inhibit its growth. For example, the abundance of Desulfovibrio in the MOs-2-a group was significantly lower than that in the control group. Desulfovibrio is closely related to the occurrence of intestinal polyps and ulcerative colitis. Reducing the number of Desulfovibrio in the intestine is important for maintaining intestinal health. 43 In conclusion, MOs-2-a can significantly improve the diversity of gut microbiota in mice and improve the flora structure by increasing the number of beneficial bacteria and reducing the number of pathogenic bacteria. It also helps to maintain the integrity of intestinal mucosa, improve digestion, and promote intestinal health.
We conclude that the MOs-2-a exhibits prebiotic effect through maintaining of intestinal mucosa integrity and modulation of gut microbiota composition. It can improve the intestinal mucosal barrier function and the activities of various digestive enzymes to degrade starch, protein, and lipid in food.
Footnotes
ACKNOWLEDGMENTS
This study was financially supported by the National Natural Science Foundation of China (Grant Nos. 31800280 and 21476103), the Natural Science Foundation of Jiangsu Province of China (Grant No. BK20180817), the Natural Science Foundation of the Jiangsu Higher Education Institutions of China (Grant No. 18KJB550006), the 2015 Jiangsu University Outstanding Science and Technology Innovation Team, and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
AUTHORS' CONTRIBUTIONS
F.W. and ZB.W. conceived and designed the experiments. YF.B. and Y.D. performed the experiment and analyzed the data. F.W., ZB.W., YF. B., and Y.D. prepared the article. XC.S. and ZB.W. reviewed the final article.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.