
Abstract
Bangpungtongseong-san (BT), an oriental herbal medicine, is used to treat obesity in Korea and East Asia and its antiobesity effects have been examined by several researchers. However, the molecular mechanisms of the antihepatic steatosis effects of BT are unclear. In this study, we examined the effects of BT on obesity, particularly nonalcoholic fatty liver disease, by analyzing metabolic and transcriptional responses using mRNA-sequencing profiles. C57BL/6J mice were fed a high-fat diet (HFD) or HFD + BT (1.5%, w/w, BT) for 12 weeks. Phenotype characteristics were estimated, and the antiobesity mechanism was examined using mRNA sequencing transcriptomic profiles in HFD-induced obese mice. BT treatment ameliorated dyslipidemia and hepatic steatosis in HFD-induced obese mice and reduced body weight gain. The levels of hepatic lipotoxicity markers were significantly decreased, while hepatic antioxidant enzyme activities were augmented by BT compared with in the HFD group. BT attenuated HFD-induced fatty liver through transcriptional changes in the liver. BT treatment downregulated mitochondrial oxidative phosphorylation-related genes in the liver, suggesting improved mitochondrial function. BT treatment also decreased the hepatic fibrosis-related transcriptome. Our findings provide insight into the antiobesity effects of BT, an alternative oriental medicine, for treating obesity-related conditions. Metabolic and transcriptional responses to diet-induced obesity with BT treatment improved liver function.
Introduction
The liver is a central organ in metabolic homeostasis and a main site for the metabolism, synthesis, storage, and redistribution of macronutrients such as carbohydrates, proteins, and lipids. 1 It acts as a hub to metabolically connect various tissues, including skeletal muscle and adipose tissue. 2 Obesity promotes alterations in hepatic lipid and glucose metabolism and is linked to the pathophysiology of nonalcoholic fatty liver disease (NAFLD). 1 NAFLD is a type of fatty liver characterized by lipid accumulation within hepatocytes because of causes other than excessive alcohol use and may progress to nonalcoholic steatohepatitis. Hepatic steatosis, the hallmark of NAFLD, occurs when the rate of hepatic fatty acid (FA) uptake from plasma and de novo FA synthesis exceeds the rate of mitochondrial FA oxidation and export as triglycerides within very low-density lipoprotein. 3,4 Therefore, excessive amounts of intrahepatic triglyceride cause metabolic imbalance and steatosis results in adverse alterations in glucose, FA, and lipoprotein metabolism. 4 NAFLD has become a major public health problem in many industrialized countries because of its high prevalence, potential progression to severe hepatic disease, and association with serious cardiometabolic abnormalities, including metabolic syndrome, type 2 diabetes mellitus, and coronary heart disease. 5 Therefore, therapeutic agents to prevent or treat NAFLD are needed.
Bangpungtongseong-san (BT), an oriental herbal medicine that comprised 18 plants, has been used in the treatment of obesity in Korea and the East Asian region. The antiobesity effects of BT have been studied by several researchers. BT was reported to have an antiobesity effect in obese patients as well as in various animal models of obesity. 6 –11 However, the molecular mechanism involved in the antihepatic steatosis effects of BT is not completely understood. In this study, we researched the antihepatic steatosis mechanisms of BT by focusing on its transcriptional and phenotypic responses in an obesogenic animal model. This is the first study to integrate the efficacy of BT with a liver tissue transcriptome by mRNA sequencing (mRNA-seq).
Materials and Methods
Animals
Twenty male C57BL/6J mice (4-week-old) were obtained from the Jackson Laboratory (Bar Harbor, ME). All mice were singly housed under a constant temperature (24°C) and 12-h light/12-h dark cycle. After 1 week of acclimatization on normal standard chow diet, at 5 weeks of age, the mice were randomly divided into 2 groups with 10 mice per group and fed either an HFD or HFD +1.5% (w/w) BT for 12 weeks. The HFD (TD06414; Harlan, Madison, WI) contained 21.3 kcal% carbohydrate, 18.4 kcal% protein, and 60.3 kcal% fat. BT extracts were obtained from Sungil Bioex (Hwaseong, Republic of Korea). The dose of BT was established based on the Ministry of Food and Drug Safety (MFDS) guidelines. The human BT dose (2850 mg/day for adults) was translated to a mouse dose using the body surface area normalization method. 12 The mice were given free access to diet and distilled water. All mice were anesthetized with isoflurane after a 12-h fasting at the end of the diet period. Blood was collected from the inferior vena cava to measure plasma lipid, glucose, and hormone concentrations. The adipose tissue and organs were isolated, rinsed with physiological saline, weighed, immediately frozen in liquid nitrogen, and stored at −80°C until further analysis. The animal study protocols were approved by the Ethics Committee of Kyungpook National University (KNU 2012-136).
Analysis of plasma and hepatic lipids
The assays for the plasma triglyceride and total cholesterol (TC) levels were performed using commercial kits (Asan Pharm Co., Seoul, Republic of Korea). Hepatic lipids were extracted as described by Folch et al. 13 ; both triglyceride and cholesterol levels were measured using the same enzymatic kit as used for plasma analyses. The plasma-free FA (FFA) and hepatic FA levels were determined using an enzymatic kit (Wako Pure Chemicals, Osaka, Japan).
Plasma glutamic oxaloacetic transaminase and glutamic pyruvic transaminase activities
Glutamic oxaloacetic transaminase (GOT) and glutamic pyruvic transaminase (GPT) activities were quantified using commercial kits (Asan Pharm Co., Seoul, Republic of Korea).
Plasma hormones, adipokines, and proinflammatory cytokines
Plasma insulin and adipokine (leptin and resistin) concentrations were measured with a multiplex detection kit (171-F7001M; Bio-Rad, Hercules, CA).
Hepatic lipid-regulating enzyme activity
FA β-oxidation and carnitine palmitoyltransferase activities were analyzed according to previously described protocols. 14,15 Microsomal HMG-CoA reductase activity was estimated using [14C]-HMG-CoA and [14C]-Oleoyl CoA as substrates. 16
Hepatic antioxidant enzyme activity
Superoxide dismutase activity was estimated as described by Marklund, which is based on the auto-oxidation of pyrogallol. 17 Catalase activity was analyzed using Aebi's method. 18 Glutathione peroxidase activity was measured as described by Paglia and Valentine with slight modification. 19 The glutathione reductase activity was analyzed by monitoring the oxidation of NADPH at 340 nm. 20
Histological analysis of liver
The histological analysis of the liver was performed as described previously. 21 The liver was isolated from each mouse, fixed in 10% (v/v) neutral-buffered formalin, and embedded in paraffin for staining with hematoxylin and eosin or Masson's trichrome dye.
RNA preparation, library preparation, and mRNA-seq
The liver was harvested from three mice randomly selected from each of the HFD and BT groups. The extraction of total RNA was performed using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA). The qualities of the cDNA libraries were estimated using an Agilent 2100 Bioanalyzer (Agilent, Santa Clara, CA). To construct cDNA libraries with the KAPA Library Quantification Kit (Kapa Biosystems, Wilmington, MA), 1ug of total RNA was used. After cluster amplification of the denatured templates, samples were sequenced as paired-end polymers (2 × 100 base pairs) using an Illumina HiSeq2500 (Illumina, San Diego, CA). Preprocessing of mRNA-seq data was analyzed according to previous protocols. 22,23
Real-time quantitative polymerase chain reaction
One microgram of total RNA was reverse-transcribed into cDNA using the QuantiTect® Reverse Transcription Kit (Qiagen, Hilden, Germany). The SYBR Green PCR Kit (Qiagen, Hilden, Germany) and the CFX96TM real-time system (Bio-Rad, Hercules, CA) were used for quantification of real-time quantitative polymerase chain reaction (RT-qPCR). The primers for RT-qPCR in this study are shown in Table 1. The 2−▵▵Ct method was used to calculate the relative gene expression level using glyceraldehyde-3-phosphate dehydrogenase. 24
Primer Sequences Used for Real-Time Quantitative Polymerase Chain Reaction
Fgf21, fibroblast growth factor 21; Gapdh, glyceraldehyde 3-phosphate dehydrogenase; Gdf15, growth/differentiation factor 15; Ndufa2, NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, 2.
Differential transcriptome and pathway analysis
The data for gene level counts were generated using HTSeq-count v0.5.4p3 for differential expression analysis. 25 Differentially expressed genes (DEGs) were determined based on a q value threshold less than 0.05 using the TCC, R software package. 26 The raw data files for the RNA-seq experiment were deposited in the Gene Expression Omnibus (GEO) database (GEO accession no.: GSE126604). Ingenuity Pathway Analysis software (IPA, Ingenuity® Systems; Qiagen, Redwood City, CA) was used for molecular pathway analysis of DEGs. We uploaded the genes with a q value threshold <0.05 and a fold change in expression ≥1.5, as well as the associated expression value from the mRNA-Seq data, to the IPA website.
Statistical analysis
Phenotypic data were presented as the mean ± standard error of the mean. Significant differences between BT and HFD groups were evaluated using unpaired Student's t-test in SPSS software (SPSS, Inc., Chicago, IL). Differences were considered statistically significant with P < .05.
Results
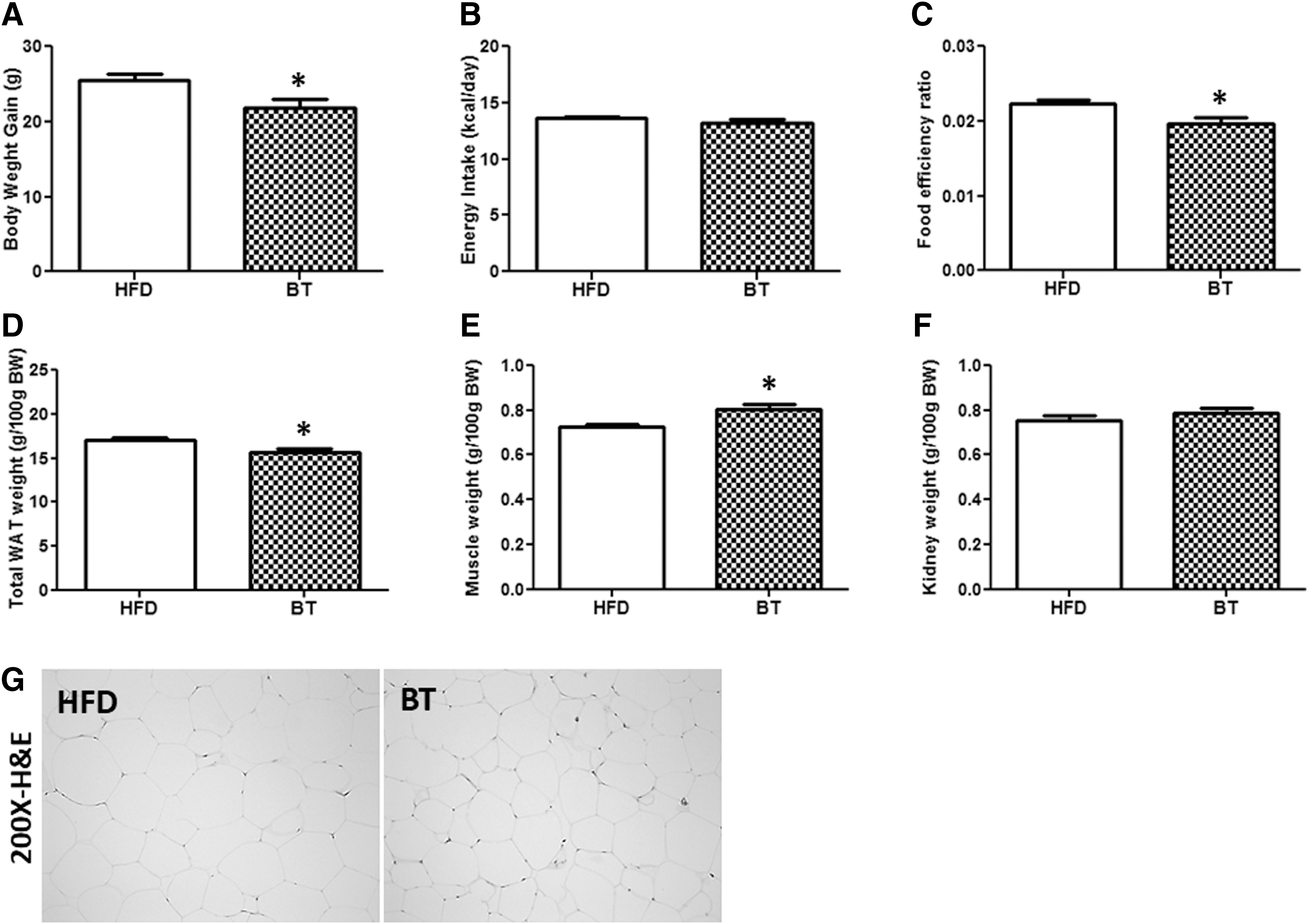
BT lowered both body weight gain and white adipose tissue mass, while augmenting muscle weight in diet-induced obese mice
Body weight gain was significantly reduced in the BT group compared with the HFD group after 12 weeks (Fig. 1A). Energy intake was not significantly different between the HFD and BT groups (Fig. 1B). Therefore, the food efficiency ratio in BT group was significantly lower compared with the HFD group (Fig. 1C). Total white adipose tissue (WAT) weight in the BT group was significantly decreased compared with that in the HFD group, while muscle weight was increased by BT treatment (Fig. 1D–F). Furthermore, epididymal WAT size in the BT group was visibly smaller than that in HFD-fed mice (Fig. 1G).
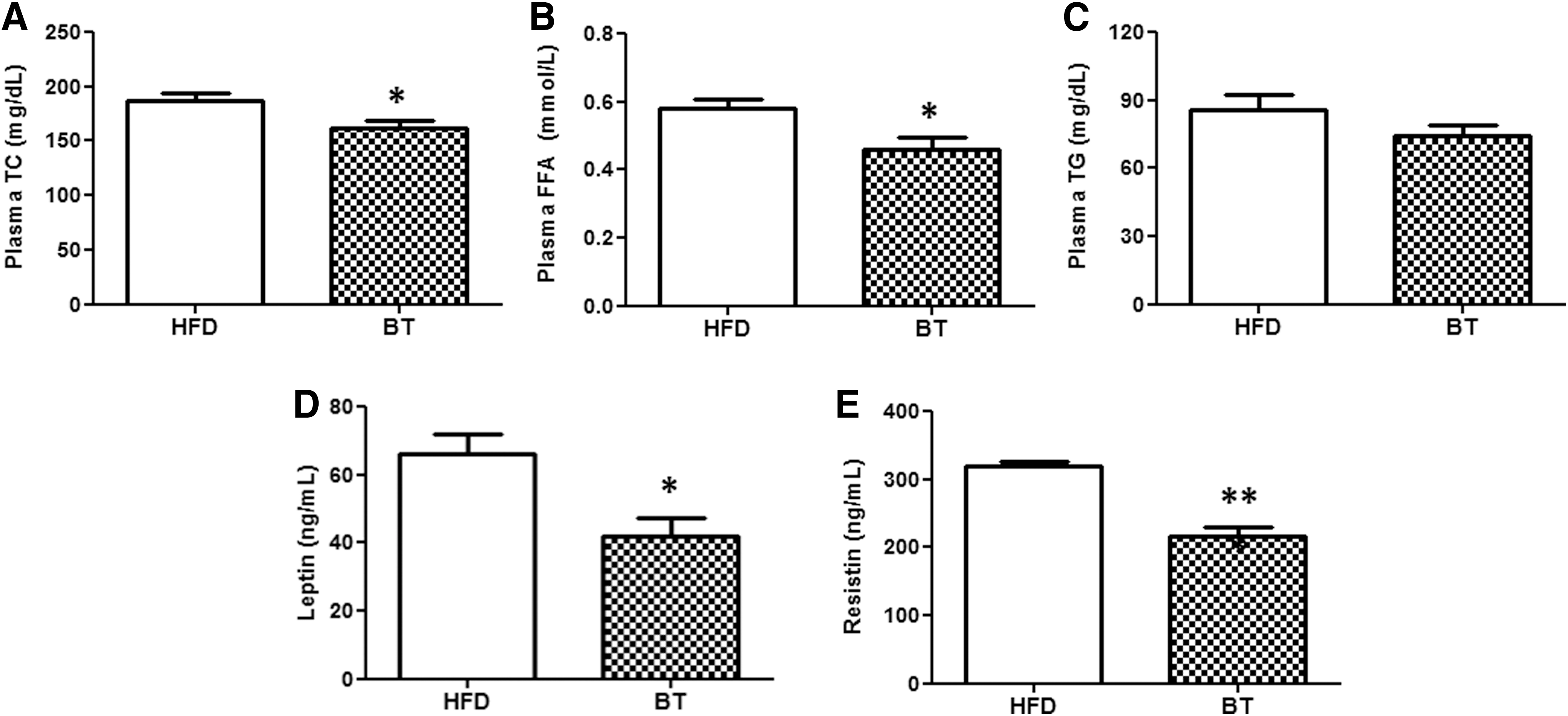
BT improved plasma lipid and adipokine levels in DIO mice
The plasma TC and FFA concentrations were significantly lower in the BT group than in the HFD group (Fig. 2A, B). There was no statistically significant difference in the plasma triglyceride between the two groups (Fig. 2C). The elevated levels of plasma leptin and resistin in the HFD group were decreased by BT treatment (Fig. 2D, E).
BT improved hepatic lipid profiles, hepatic tissue morphology, while altering hepatic lipid regulating enzyme activities in DIO mice
When liver weight was expressed as g/100 g body weight, this value was significantly lower in the BT group than in the HFD group (Fig. 3A). Hepatic FA and cholesterol contents in the BT group were significantly lower than those in the HFD group (Fig. 3B, C). However, hepatic triglyceride content did not differ between the HFD and BT groups (Fig. 3D). Hepatic tissue morphology also showed decreased accumulation of lipid droplets in the BT group (Fig. 3E). The HMG-CoA reductase activity, hepatic cholesterol-regulating enzyme, was significantly reduced in the BT group compared with the HFD group (Fig. 3F). The activities of the two enzymes, associated with FA oxidation, β-oxidation, and carnitine palmitoyl-CoA transferase, did not significantly differ between the HFD and BT groups (Fig. 3G, H). The levels of plasma GOT and GPT were markedly attenuated by BT treatment (Fig. 3I, J).

BT partly improved hepatic antioxidant enzyme activities in DIO mice
The hepatic enzyme activities of superoxide dismutase and glutathione peroxidase did not differ between the HFD and BT groups (Fig. 4A, B). In contrast, the activities of catalase and glutathione reductase were significantly higher in BT-treated mice than in HFD mice (Fig. 4C, D).

BT modulates transcriptional responses of liver in DIO mice
We performed mRNA-Seq on liver samples harvested from mice and systematically analyzed the results as follows to identify the global transcriptomic profiles related to hepatic steatosis. We identified 523 DEGs (169 up- and 354 downregulated genes) between the BT and HFD groups using a cutoff of |fold change| ≥ 2 and q-value <0.05. The BT phenotype was underpinned by a set of 523 DEGs (in the liver) that showed expression changes only in the BT group (Fig. 5A–C). To validate the mRNA-seq data, we performed RT-qPCR. We selected Gdf21, Fgf21, and Ndufa2 from the DEG list (BT vs. HFD). The results showed good agreement between the mRNA-seq- and RT-qPCR-determined expression levels (Fig. 5D). The top-ranked canonical pathways in the liver are shown in Figure 6A as a comparison of BT versus HFD. DEGs were involved in canonical pathways, including oxidative phosphorylation, JAK/Stat signaling, IGF-1 signaling, GADD45 signaling, ATM signaling, IL-12 signaling, and production in macrophages, hereditary breast cancer signaling, mitochondrial dysfunction, hepatic fibrosis/stellate cell activation, and glutathione-mediated detoxification pathway (Fig. 6A). Particularly, BT led to downregulation of oxidative phosphorylation (OXPHOS), mitochondrial dysfunction, and hepatic fibrosis compared with the HFD group (Fig. 6B). We also found that hepatic fibroblast growth factor 21 (FGF21) and growth/differentiation factor 15 (GDF15), biomarkers for NAFLD, were significantly attenuated in BT mice compared with HFD-fed mice (Fig. 6C).


Discussion
In this study, we investigated the effect of BT and possible mechanisms of the fundamental metabolic regulation using transcriptomic profiles in a diet-induced obesity (DIO) model. BT treatment improved HFD-induced dyslipidemia, hepatic steatosis, and obesity and augmented the antioxidant activities in obese mice. Significant improvements in adipokine secretion, including leptin and resistin, were observed in BT mice, compared with HFD mice. Leptin, the satiety hormone, regulates appetite and energy balance. However, leptin resistance is a hallmark of DIO. Obese rodents have elevated leptin levels but blunted leptin sensitivity and such rodents have apparent central and peripheral leptin resistance. 27 Resistin boosts both insulin resistance and inflammation in animal models. 28,29 Therefore, it is considered that BT treatment effectively improves HFD-induced adipokine imbalance.
The antiobesity action of BT was consistent with that described previously using other models of obesity. 6 –11 In a previous study, chronic treatment with BT (4.7% BT, more than threefold the experimental intake used in this study) in KKAy mice decreased adipocyte hypertrophy in the WAT by activating brown adipose tissue thermogenesis and ameliorating adipocytokine dysregulation. 6 Similar to our findings, Yoshikawa and colleagues reported that BT treatment significantly reduced the WAT/body weight ratio and WAT size in HFD mice but did not affect dietary intake. 11 While previous studies have focused primarily on the role of BT in adipose tissue, our study focused on its action in liver tissue. Particularly, this is the first report on the efficacy of BT using liver tissue transcriptomes obtained from mRNA-seq.
The primary factor promoting hepatic steatosis is an imbalance in lipid flux in the liver. 30 Various factors result in lipid accumulation in the liver, for example, increased lipolysis in adipose tissues and/or high dietary fat intake, increased de novo hepatic lipid synthesis, decreased fat oxidation, and decreased hepatic very low-density lipoprotein secretion. 30,31 Enlarged adipose tissues release more FFA into the circulation, which results in greater infusion into the liver.
In this study, HFD feeding for 12 weeks successfully induced liver injury and fatty liver. In addition, BT significantly reversed hepatic lipid accumulation by HFD, as well as elevated plasma lipids. Furthermore, BT treatment reduced plasma GPT and GOT levels, indicating that attenuated liver damage was induced in the obesogenic environment. Possible explanations for the lower hepatic lipid levels in the BT group include decreased adiposity, reduced flux in FFA via the portal vein from the visceral fat to the liver, and downregulation of hepatic cholesterol-regulating enzymes.
HFD and dysregulated lipid metabolism cause hepatic steatosis, and under these conditions, a metabolic shift is caused to resist the hepatic lipid burden. This shift includes augmented mitochondrial FA oxidation, tricarboxylic acid cycle induction, and OXPHOS stimulation. 32 Mitochondria play a critical role in generating energy via nutrient oxidation. 33 In addition to the role of mitochondria in the oxidation of fat and glucose to generate energy, mitochondria produce reactive oxygen species (ROS). 34,35 Mitochondrial dysfunction is induced via altered OXPHOS and ROS overproduction. 36 Moreover, mitochondrial dysfunction leads to the pathogenesis of NAFLD, as it promotes ROS production and affects hepatic lipid homeostasis and cytokine release, and lipid peroxidation. 33,36 Excessive ROS exceeding the cell's antioxidant capacity can damage cellular components such as nucleic acids (particularly mitochondrial DNA), proteins, and lipids, leading to oxidative stress and ultimately apoptosis. 37,38 In animal models of NAFLD, mitochondrial dysfunction is characterized by alterations in the abundance and activity of OXPHOS. 32 According to IPA, OXPHOS is the most significant pathway among the 53 canonical pathways (P-value <.05) in the liver. A majority of the OXPHOS-related genes, including NADH dehydrogenase (complex I), cytochrome c oxidase (complex IV), and ATP synthase, were decreased by BT treatment. Therefore, it is likely that BT supplementation can partially contribute to improvements in mitochondrial function, especially OXPHOS transcriptional response, through enhanced antioxidant effect and reduced hepatic FFA burden.
Meanwhile, FGF21 and GFD15 have been suggested as markers of mitochondrial respiratory chain deficiency and NAFLD. 39 –44 FGF21 is a hepatic protein that plays a central role in metabolism, stimulating FA oxidation in the liver, and glucose uptake in fat. 45 The consistent increase in FGF21 observed in human obesity may reflect the presence of fatty liver disease, indicating that FGF21 can be used as a biomarker of NAFLD. 40 GDF15 is an endocrine hormone belonging to the transforming growth factor β superfamily, and elevated expression of GDF15 occurs following diet-induced hepatic endoplasmic reticulum stress. 46 Kim et al. reported that GDF15 can serve as a novel biomarker of advanced fibrosis in NAFLD. 47 Consistent with previous studies, marked decreases in hepatic Fgf21 and Gdf15 mRNA expression were observed after BT treatment.
Of top ranked canonical pathways, “JAK/STAT signaling” was the second rated pathway. The activation of the JAK/STAT signaling pathways plays a complex role in regulation of liver fibrogenesis, especially STAT1 acts as an antifibrotic signaling molecule via the induction of hepatic stellate cell apoptosis and cell cycle arrest. 48 In this study, BT regulated mRNA expression of JAK/STAT signaling pathway-related genes, including Stat1, Fos, Jun, Cdkn1a, Cish, Socs2, Cebpb, and Atm. Furthermore, the regulation of IGF-1 signaling, including IGF binding protein-1 (Igfbp-1), Igfbp-3, Igfbp-6, and Igfbp-7, was also observed in transcriptional response by BT treatment. In addition, there were substantial transcriptional differences between HFD and BT mice in molecular pathways, including GADD45 signaling, ATM signaling, IL-12 signaling, and production in macrophages, hereditary breast cancer signaling, mitochondrial dysfunction, and hepatic fibrosis/hepatic stellate cell activation in liver.
In summary, our data indicate that BT treatment can suppress or improve DIO and obesity-associated metabolic disorders, particularly NAFLD. BT attenuated hepatic lipid accumulation and decreased the transcriptional response of OXPHOS in the liver, suggesting improved mitochondrial function after BT treatment. Moreover, BT was an effective antioxidant supplement for NAFLD. Taken together, our findings provide an important mechanistic insight into the anti-NAFLD effects of transcriptional regulation of mitochondrial function and antioxidant defense in the liver. These results suggest that dietary BT may delay NAFLD via maintaining liver physiology and thereby normal function. Therefore, BT may be an effective treatment for pathological alterations in NAFLD by targeting mitochondrial dysfunction in NAFLD.
Footnotes
Acknowledgment
This work was supported by a National Research Foundation of Korea (NRF) grant, funded by the Ministry of Science and ICT (NRF-2012M3A9C4048818, NRF-2016R1C1B1014680, NRF-2019R1C1C1009530), and the BK21 Plus Program (Department of Food Science & Nutrition, Kyungpook National University, 22A20130000161), funded by the Ministry of Education (MOE, Korea) and NRF.
Author Disclosure Statement
No competing financial interests exist.