
Abstract
Previous studies have proven that polysaccharide obtained from the seeds of Plantago asiatica L. (PLCP) could induce maturation of murine dendritic cells, promote defecation, and possess antioxidant activity in vitro. However, the effect of PLCP on lipopolysaccharide (LPS)-induced liver injury in mice has been rarely reported. In this study, we investigated the anti-inflammatory effect of PLCP on LPS-induced liver injury. Mice were pretreated orally with different dose of PLCP for 3 weeks. On day 22, they were injected intraperitoneally with LPS and sacrificed 12 h later. The results showed that PLCP inhibited the excessive production of tumor necrosis factor-α, interleukin (IL)-6, IL-10, IL-2, and IL-1β in mouse serum and liver. PLCP also improved glutathione peroxidase and total antioxidant capacity activities in mouse liver. In addition, PLCP inhibited nitric oxide, inducible nitric oxide synthase, and cyclooxygenase-2 expression, and increased metallothionein production in mouse liver. Consequently, PLCP may possess protective effects on inflammatory associated liver injury.
Introduction
Appropriate inflammatory response is beneficial for killing bacteria. However, unsuitable or excessive inflammation is harmful and sometimes even associated with fatal diseases, such as fever, inflammation, sepsis, and endotoxin shock. 1 Inflammation involved in the liver wound healing response is referred to as toxic hepatitis. Unless exposure is stopped, toxins may lead to chronic disease and even cirrhosis.
Lipopolysaccharide (LPS) is a stable macromolecule, and it is the toxic component of the outer cell wall of Gram-negative bacteria. LPS exposure occurs not only in bacterial infection but also from the organisms itself. After exposure to LPS, organisms will induce a series of host inflammatory response. Liver is an important organ to remove intestinal bacteria and the LPS endotoxin. It is considered to be the final barrier to stop LPS from entering the systemic bloodstream. 2 LPS-induced liver injury has been universally used as a model to explore the mechanism of endotoxin-induced liver injury and evaluate anti-inflammatory activities of drugs or compounds. 3,4 Kupffer cells, as resident macrophages in liver, are activated by bacteria, endotoxin, and microbial debris by producing inflammatory and fibrogenic cytokines, as well as reactive oxygen species (ROS). 5 Their activation is necessary to the response of the liver to infection or injury. It has been demonstrated that LPS-induced liver injury often leads to overproduction of proinflammatory cytokines and ROS. 6 High expression of interleukin (IL)-6 after LPS challenge has also been considered a sign of liver injury. 7 As is well known, oxidative damage plays a remarkable role in hepatic injury mediated by poisons and drugs. Inflammation induced by LPS also results in oxidative stress by producing oxidants. 8 Thus, antioxidant enzymes may play central roles in the defense of body against oxidative stress. Glutathione peroxidase (GSH-Px) is one of the important peroxide-degrading enzymes in the body. Its biochemical function is reducing lipid hydroperoxides to alcohols and free hydrogen peroxide to water. It was reported that mice injected with LPS showed a marked increase in ROS production evidently by low GSH-Px activities. 9 Inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) are two inflammatory mediators stimulated by oxidative stress or injury. Overexpression of iNOS and COX-2 is harmful to the liver by increasing nitric oxide (NO) and prostaglandin productions.
Plantago asiatica L. is a perennial plant considered to be both medicinal and edible. It is famous for its diuretic, anti-inflammatory, and antiasthmatic functions. 10 It also has been traditionally used in Asian pharmacopoeia for the treatment of liver disease. In the past few decades, many plant polysaccharides have been widely reported for their chemical properties and bioactivities. 11,12 Recently, our group has extracted and purified a polysaccharide from the seeds of P. asiatica L. (PLCP). Arabinoxylan is considered to be its main component because of its high content of arabinose and xylose. The main linkage type was β-1,4-linked Xylp, with highly branched structure of β-T-linked Xylp, α-1,3-linked Araf, and α-T-linked Araf. 13 –15 Moreover, we demonstrated that PLCP possesses antioxidant activities in vitro, 16 induces maturation of murine dendritic cells, 17 and promotes defecation. 18 As lots of researches have demonstrated the tight connection between antioxidative capacity and anti-inflammatory activity, the effects of PLCP on liver of LPS-induced mice were observed in this article.
Materials and Methods
Reagents
The seeds of P. asiatica L. were obtained from Ji'an (Jiangxi, China). LPS (Escherichia coli 0127:B8) was purchased from Sigma-Aldrich (St. Louis, MO, USA). Tumor necrosis factor-α (TNF-α), IL-6, IL-10, and IL-1β enzyme-linked immunosorbent assay (ELISA) kits were purchased from Boster (Wuhan, China) for serum and Cloud-Clone (Wuhan, China) for liver homogenates. IL-2 ELISA kit was purchased from Cloud-Clone. NO assay kit, BCA protein assay kit, GSH-Px, and total antioxidant capacity (T-AOC) kits were obtained from Beyotime Biotechnology (Shanghai, China). Mouse metallothionein (MT) ELISA kit was purchased from Bogoo (Shanghai, China). Whole protein extraction kit was obtained from KeyGEN BioTECH (Jiangsu, China). Manganese superoxide dismutase (MnSOD), iNOS, and COX-2 antibodies were purchased from Cell Signaling Technology (Beverly, MA, USA).
Preparation of PLCP
Preparation of PLCP was carried out according to our previous report. 16 In brief, the seeds of P. asiatica L. (100 g) were soaked in ethanol (80%, v/v) for 24 h, dried naturally, and then extracted with 10-fold boiling distilled water for 3 h. The residue was repeated for three times. The combined aqueous extract was centrifuged, filtered, concentrated, and deproteinized. The resulting aqueous solution was separated from the organic matter, dialyzed, and precipitated with ethanol (4:1) overnight. After centrifugation, the precipitate was redissolved in water and lyophilized to obtain polysaccharide (PLCP).
Animals
Sixty BALB/c mice (specific pathogen free, 18–22 g weight) were purchased from Hunan SJA Lab Animal Ltd. (certificate number SCXK [xiang] 2011-003, Hunan, China). The mice were maintained on a 12/12 h of light–dark cycle with 55% ± 10% relative humidity at 23°C ± 2°C temperature for 1 week of adaptation. The mice were fed ad libitum. All experiments followed NIH rules for the care and use of laboratory animals and the study protocol was approved by the Nanchang University Animal Ethics Review Committee.
Animal experiment
The mice were randomly divided into 5 groups (12 for each group): control group, model group, and PLCP treatment group. Mice in the PLCP treatment group were intragastrically administered with low, middle, and high dose of PLCP (50, 100, and 200 mg/kg body weight) continuously for 21 days, whereas mice in control and model groups were orally administrated with equal volume of normal saline. Then mice in model and PLCP treatment groups were intraperitoneally injected with LPS (10 mg/kg) on the 22nd day for 12 h, whereas mice in the control group received an equal volume of normal saline simultaneously. Twelve hours later, all mice were killed after bleeding by eyeball enucleation, and the livers were removed immediately.
Measurement of cytokine levels in serum and liver
The effects of PLCP on the levels of cytokines in serum and liver homogenates were measured. For serum, TNF-α, IL-6, IL-10, and IL-1β production was determined directly by using commercially available ELISA kits. Livers were homogenized in precooled phosphate-buffered saline (PBS) and centrifuged to obtain supernatants, which were then assayed according to the directions for the ELISA kits.
Assays for antioxidant enzyme activities in liver
The activities of GSH-Px and T-AOC in liver homogenate were detected by commercial kits. In addition, protein concentrations of the supernatant were measured by BCA protein assay kit. The experimental data were presented as the enzymatic activity per unit mass of the protein.
NO assay
To detect the concentration of NO in the liver, supernatant of liver homogenates were obtained as described in the procedure of cytokine measurement in liver. Then, 50 μL of Griess Reagent I and II was added, respectively, to 50 μL of supernatant, and the absorbance at 540 nm was determined by using a microplate reader.
Evaluation of MT expression
MT is considered to be an important free radical scavenger 19 ; thus, MT expression in liver was determined. The supernatant from liver homogenates was obtained by homogenizing liver with equal volume of PBS (1 mg/mL) and centrifuging for 20 min at 827 g. Then MT was quantified using mouse MT ELISA kit.
Western blotting analysis
Liver proteins were extracted according to the operating instructions of whole protein extraction kit, and protein concentration was measured by BCA protein assay kit. The protein lysate was mixed with sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) protein loading butter and denatured at 100°C for 5 min. Then, they were separated by 10% SDS-PAGE and transferred to PVDF membrane. The membrane was incubated with primary antibodies against MnSOD, iNOS, and COX-2 (1:1000 dilution) overnight at 4°C after blocking for 1 h with 5% bovine serum albumin at room temperature. Then, the membrane was incubated with horseradish peroxidase-conjugated secondary antibody (1:10,000 dilution) for 1 h at room temperature. The intensity images were enhanced by reacting with BeyoECL Plus and exposure in Bio-Rad Chemi XRS+ gel image system.
Statistical analysis
IBM SPSS statistics 19.0 software was used to analyze the data. The results are expressed as mean ± standard deviation, and significant differences between groups were calculated with least-significant difference of one-way analysis of variance. A value of P < .05 indicated statistical significance.
Results
Effects of PLCP on cytokine levels in serum of LPS-challenged mice
The levels of TNF-α, IL-6, IL-10, and IL-1β in serum of LPS-challenged mice were assayed in this study. The results are shown in Figure 1. Twelve hours after LPS challenge, there was a significant increase in the expression of TNF-α (from 58.56 to 2354.35 pg/mL [P < .01]), IL-6 (from 50.60 pg/mL to 44.52 ng/mL [P < .01]), IL-10 (from 56.14 to 1256.55 pg/mL [P < .01]), and IL-1β (from 112.89 to 1522.68 pg/mL [P < .01]) compared with that of the control group. However, pretreatment with PLCP could greatly suppress LPS-induced TNF-α, IL-6, IL-10, and IL-1β production.

Effects of PLCP on cytokine levels in LPS-challenged mouse serum. Mice were orally administered with PLCP (50, 100, and 200 mg/kg body weight) for 21 days, then intraperitoneally injected with LPS (10 mg/kg) on the 22nd day. Serum levels of TNF-α, IL-6, IL-1β, and IL-10 were measured by ELISA 12 h after intraperitoneal injection. Data are shown as mean ± SD. Significant differences were ▴ P < .01 as compared with the control group; *P < .01 as compared with the model group. ELISA, enzyme-linked immunosorbent assay; IL, interleukin; LPS, lipopolysaccharide; PLCP, polysaccharide obtained from the seeds of Plantago asiatica L.; SD, standard deviation; TNF-α, tumor necrosis factor-α.
Effects of PLCP on antioxidant enzyme activities and T-AOC in livers of LPS-challenged mice
The activities of GSH-Px, T-AOC, and MnSOD in liver were also determined (Fig. 2). Results showed that the mice challenged with LPS alone had significantly lower GSH-Px (from 783.59 to 466.68 U/g protein [P < .01]) and T-AOC (from 37.22 to 23.28 μmol/g protein [P < .01]) activities in the liver compared with that of the control group. In addition, results of western blotting showed that LPS challenge also decreased liver MnSOD activity by 38.79 ± 0.01% (P < .01), whereas after being pretreated with PLCP, GSH-Px, T-AOC, and MnSOD activities improved significantly.
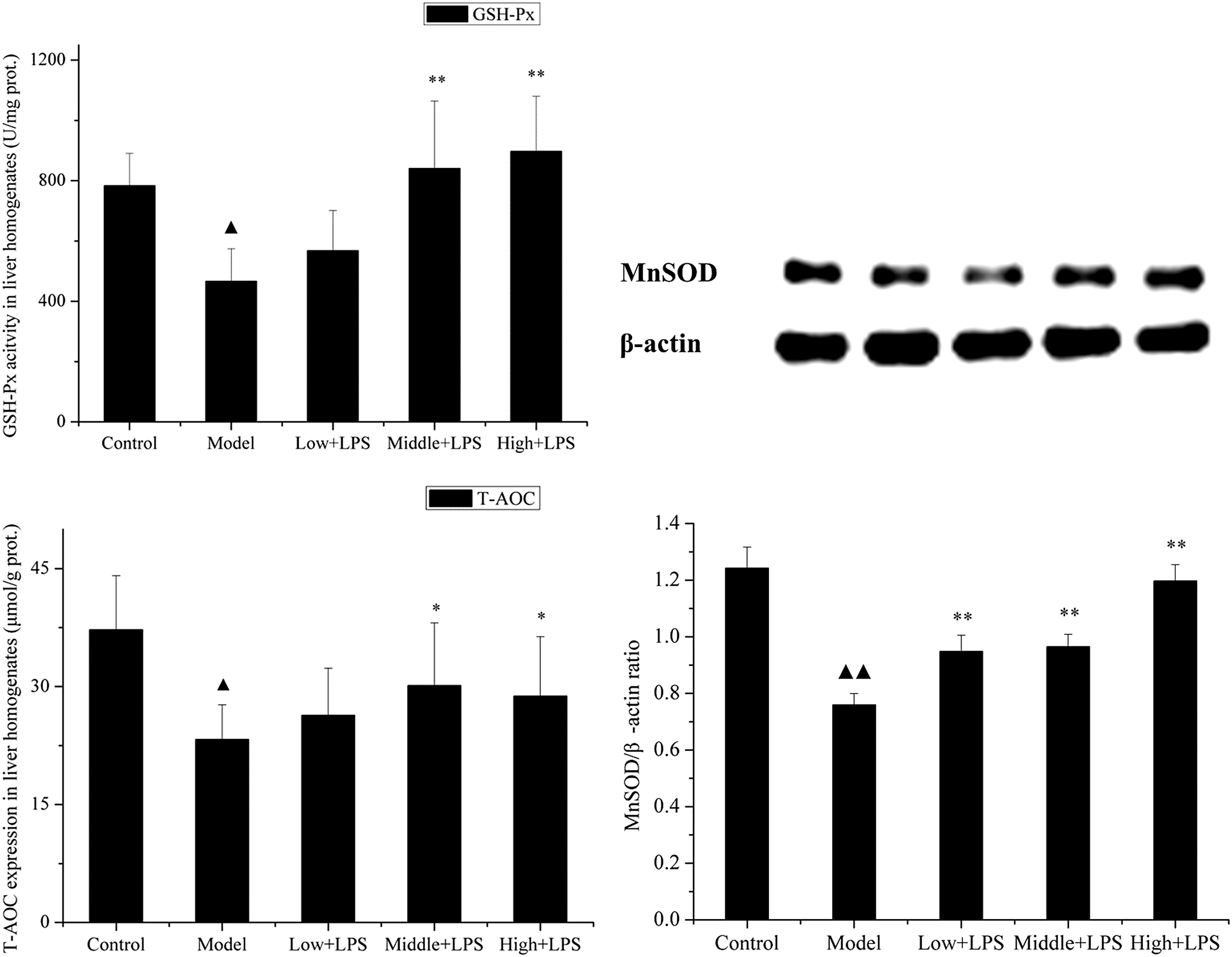
Effects of PLCP on antioxidant enzyme activities and T-AOC in liver homogenates of LPS-challenged mice. Mice were orally administered with PLCP (50, 100, and 200 mg/kg body weight) for 21 days, then intraperitoneally injected with LPS (10 mg/kg) on the 22nd day. Liver tissues of each group were collected 12 h after intraperitoneal injection. GSH-Px activity and T-AOC were determined according to commercial kits, whereas MnSOD activity was confirmed by Western blotting and displayed by MnSOD/β-actin ratio. Data are shown as mean ± SD. Significant differences were ▴ P < .05 versus control, ▴▴ P < .01 versus control; *P < .05 versus model, **P < .01 versus model. GSH-Px, glutathione peroxidase; MnSOD, manganese superoxide dismutase; T-AOC, total antioxidant capacity.
Effects of PLCP on cytokine levels in liver of LPS-challenged mice
The levels of TNF-α, IL-6, IL-10, IL-1β, and IL-2 in liver homogenate of LPS-challenged mice were also assayed in this study. The results are shown in Figure 3. Twelve hours after LPS challenge, there was a significant increase in the expression of TNF-α, IL-6, IL-10, IL-1β, and IL-2 compared with that of the control group from 137.52, 287.33, 22.81, 301.46, 728.13 to 358.80 ng/mL (P < .01), 388.10 ng/mL (P < .01), 27.65 ng/mL (P < .05), 336.19 ng/mL (P < .05), 1224.14 ng/mL (P < .01), respectively. Pretreatment with PLCP greatly suppressed LPS-induced TNF-α, IL-6, IL-10, IL-1β, and IL-2 production. The results were in accordance with that of serum.
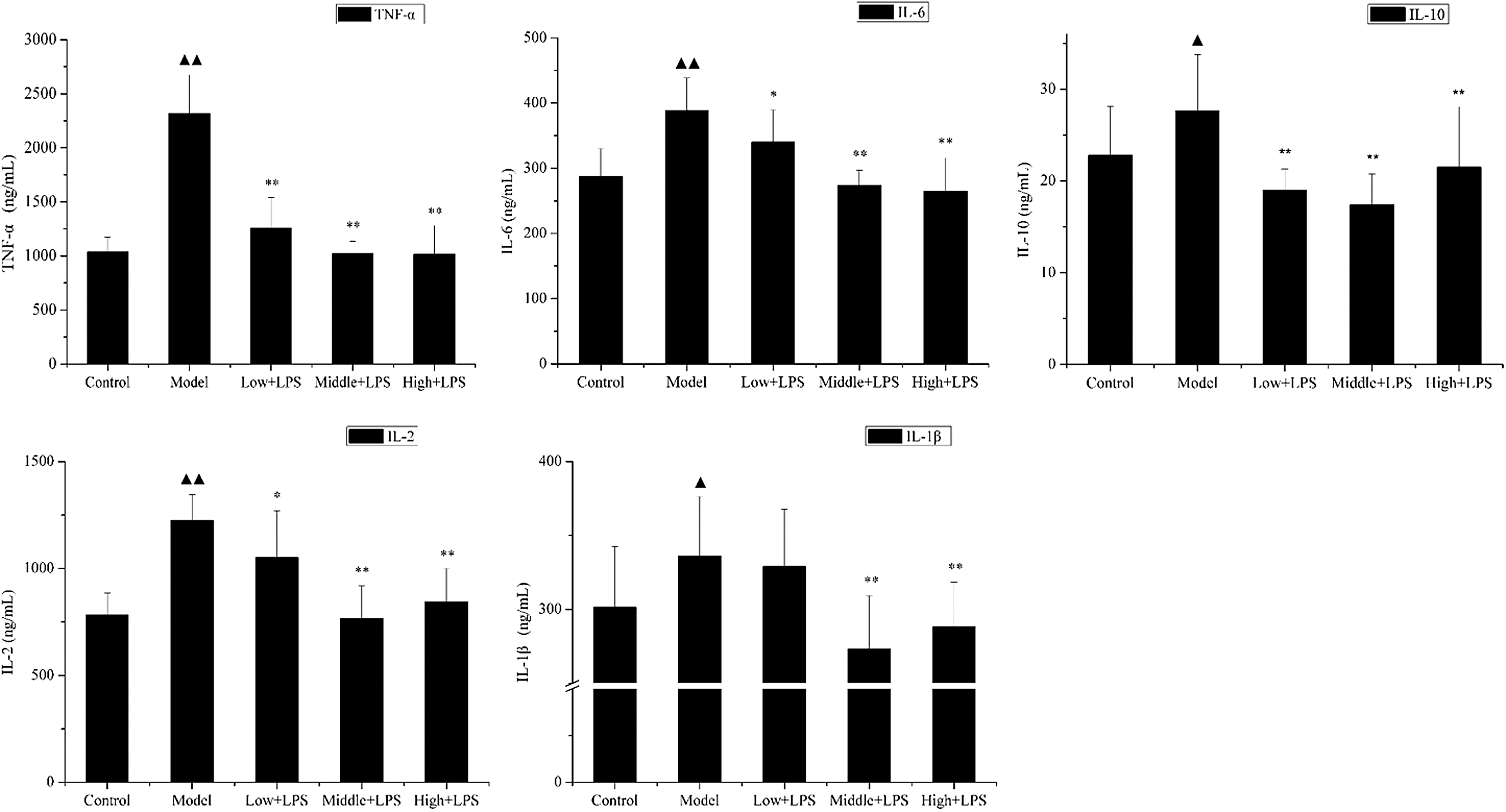
Effects of PLCP on cytokine levels in liver homogenates of LPS-challenged mice. Mice were orally administered with PLCP (50, 100, and 200 mg/kg body weight) for 21 days, then intraperitoneally injected with LPS (10 mg/kg) on the 22nd day. Liver tissues of each group were collected 12 h after intraperitoneal injection. Cytokine levels of liver homogenates were determined according to ELISA kits. Data are shown as mean ± SD. Significant differences were ▴ P < .05 versus control, ▴▴ P < .01 versus control; *P < .05 versus model, **P < .01 versus model.
Effects of PLCP on MT content in liver of LPS-challenged mice
The MT expression in liver is shown in Figure 4. The level of MT expression in the control group was 192.91 ± 28.11 pg/mL. However, it was dramatically decreased to 143.29 ± 28.11 pg/mL (P < .01) 12 h after LPS challenge. In contrast, middle and high dose of PLCP pretreatment improved this expression significantly (178.62 ± 43.57 and 179.26 ± 43.13 pg/mL, respectively [P < .05]).

Effects of PLCP on MT content in LPS-challenged mouse liver homogenates. Mice were orally administered with PLP (50, 100, and 200 mg/kg body weight) for 21 days, then intraperitoneally injected with LPS (10 mg/kg) on the 22nd day. MT expression of liver homogenates was determined 12 h after intraperitoneal injection. Data are shown as mean ± SD. Significant differences were ▴ P < .01 as compared with the control group; *P < .05 as compared with the model group. MT, metallothionein.
Effects of PLCP on NO, iNOS, and COX-2 expressions in the liver of LPS-challenged mice
Furthermore, we observed the effect of PLCP on LPS-induced changes in hepatic NO expression. The NO expression in liver homogenate of LPS-challenged mice was dramatically increased compared with the control group (from 14.52 to 27.72 μM; P < .05). However, pretreatment with PLCP dose-dependently decreased the level of NO to 22.30, 21.59, and 19.64 μM in low, middle, and high dose of PLCP, respectively (Fig. 5). In addition, there was a dramatical increase in the expressions of hepatic COX-2 and iNOS proteins by 0.77% ± 0.06% (P < .01) and 0.30% ± 0.09% (P < .01), respectively, 12 h after LPS challenge. In contrast, PLCP pretreatment showed a significant decrease in expressions of COX-2 and iNOS compared with the model group (Fig. 5).
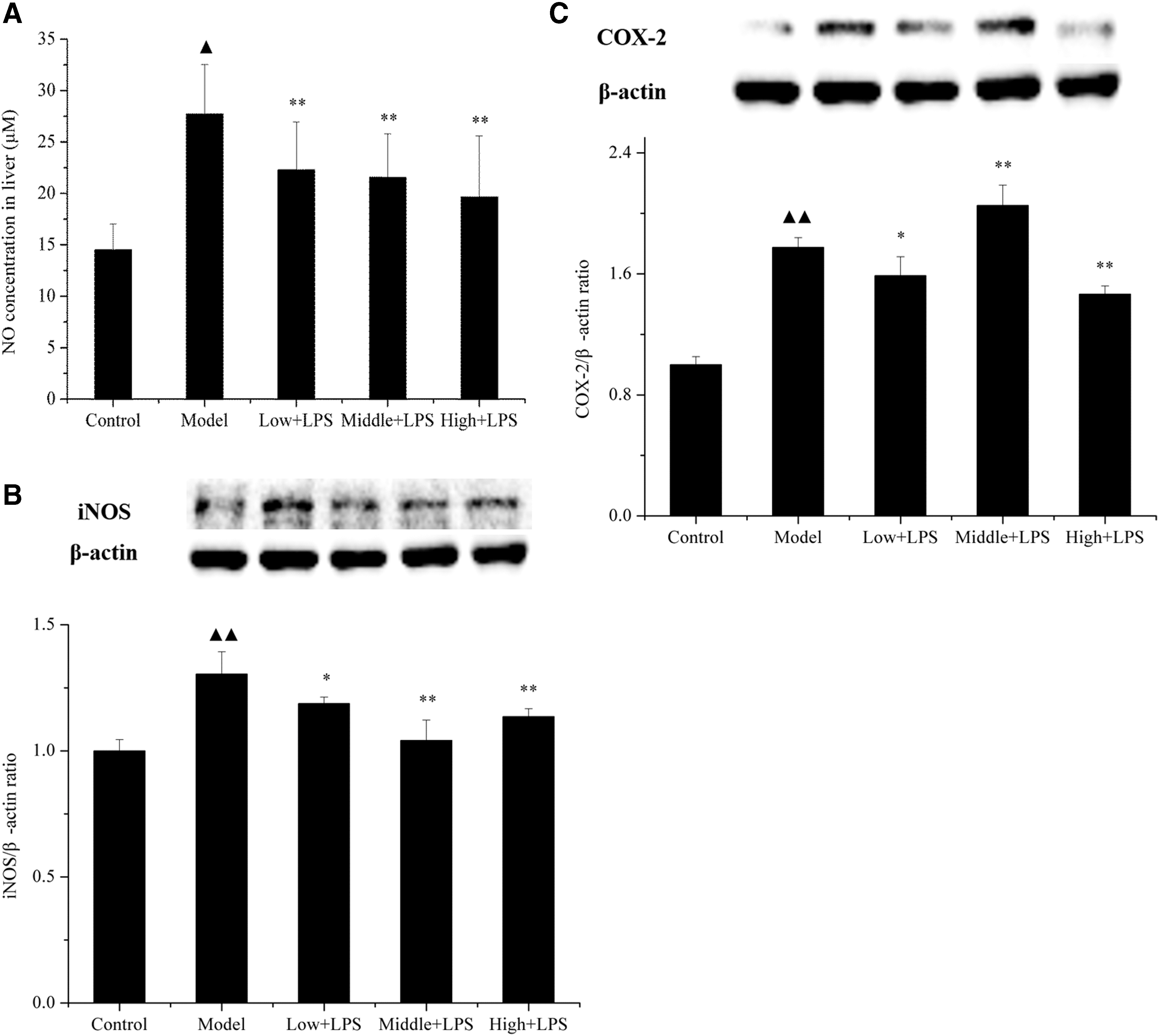
Effects of PLCP on NO
Discussion
Intraperitoneal injection of LPS to induce systemic inflammation is well documented in many studies. 20,21 It has been shown in mice that LPS increases the level of inflammatory mediator IL-6 systemically. 7 In our study, the level of IL-6 in serum and liver was dramatically increased after LPS injection (Figs. 1 and 3), which means that successful liver injury model was established.
It is well known that LPS is recognized by binding Toll-like receptor 4 (TLR4) on the cell surface and then initiates cytokines secretion. 22,23 LPS can stimulate cells to release TNF-α, which in turn triggers a cascade of other cytokines that cooperate to kill hepatocytes, recruit inflammatory cells, and initiate a wound healing response. 24 LPS interferes with cytokines' usual protective responses and cause liver injury. 25 TNF-α and IL-1β are two early and important mediators produced by Kupffer cells in liver injury. 26 Our results showed that LPS challenge led to abnormal increase of TNF-α, IL-6, and IL-1β expression in serum and liver compared with the control group. This indicated that serious injury had occurred. However, when the mice were pretreated with PLCP, the decrease in cytokines secretion was found in serum and liver 12 h after LPS challenge. IL-10 is considered to be a unique cytokine due to its anti-inflammatory and immunosuppressive functions. 27 Overexpression of IL-10 can inhibit the proinflammatory response to bacteria, which causes pathogens to escape from immune control. 28 It was reported that LPS-induced acute lung injury caused a high expression of IL-10, which was associated with NF-κB signaling pathways. 29 Thus, the balance between proinflammatory and anti-inflammatory cytokines is of vital importance. Our results showed that PLCP could control this balance by downregulating LPS-induced IL-10 overexpression (Figs. 1 and 3), which was consistent with our previous study in vitro. 30 The release of IL-10 secreted by Kupffer cells was maximal 12–24 h after LPS challenge, whereas IL-6 and TNF-α release were inhibited by IL-10 6 h later. 31 LPS exposure activated the three mitogen-activated protein kinase (MAPK) pathways: p38, extracellular signal-regulated kinases (ERK) 1 and 2, and c-Jun N-terminal kinase (JNK), which in turn activated various transcription factors that coordinated the induction of many genes encoding inflammatory mediators as well as anti-inflammatory cytokines. 32 Some researchers have demonstrated that the transcription factor Sp1 is vital in IL-10 transcription. 33 –35 Therefore, we speculate that overexpression of IL-10 in our study may be the results of concomitant activation of p38, ERK, and JNK MAPKinases, which control downstream signaling to Sp-1 transcription factor. IL-2 is also a proinflammatory cytokine, mainly produced by T lymphocytes. It markedly increased after LPS challenge. 36 PLCP was proved to downregulate the high expression of IL-2 caused by LPS. All these results indicated that the protective effects of PLCP on the liver of LPS-challenged mice were partly attributed to inhibition of TNF-α, IL-6, IL-10, IL-2, and IL-1β.
The serious damage induced by endotoxin to various organs is caused by the high production of reactive oxygen intermediates such as O2 −, peroxides, and NO. ROS are important signaling molecules in the progression of inflammatory disorders. An increased ROS generation at the site of inflammation causes tissue injury. Once exposure to LPS, neutrophils and Kupffer cells would accumulate in injury site and release ROS. 5 Low level of ROS may function as an intracellular messenger through modulating NF-κB signaling pathways, 37 whereas too much ROS leads to cell death. 9 NO is a kind of free radical that may cause severe body damage. It is reported that high NO expression produced by iNOS appears in liver after LPS challenge. 9 In this study, nitrite level in the liver was significantly increased after LPS injection. Mice with PLCP pretreatment successfully ameliorated this rise showing the potential protection of PLCP against nitrosative stress (Fig. 5). In addition, iNOS and COX-2 are involved in promoting inflammatory processes. LPS has been shown to induce iNOS expression in Kupffer cells and hepatocytes. 9 iNOS then synthesizes large amounts of NO, which can induce oxidant stress. Inhibition of iNOS and COX-2 activity may become an effective therapeutic strategy for prevention of inflammation. In this research, as shown in Figure 5, there was a dramatical increase in the expression of hepatic COX-2 and iNOS proteins 12 h after LPS challenge. In contrast, PLCP pretreatment showed a significant decrease in expression of COX-2 and iNOS proteins compared with the model group. These results indicated that anti-inflammatory activities of PLCP were attributed to the inhibition of iNOS that decreased NO expression, as well as the inhibition of COX-2.
The imbalance between ROS and antioxidant defense is a well-established cause of various diseases. MT was found to be an effective antioxidant against ROS and reactive nitrogen species (RNS). It could be rapidly induced in vitro and in vivo by many stimuli, such as hormones, cytokines, and oxidative stress. MT is currently known as the most effective free-radical scavenger. 19 Its ability to scavenge free radicals (•OH) is about 340-fold higher than that of GSH. In the human body, large quantities of MT are synthesized primarily in the liver and kidneys. Thus, we determined the MT expression in liver. Results showed that LPS challenge significantly decreased MT expression as compared with the control group (192.91 vs. 143.29 pg/mL), whereas pretreatment with PLCP enhanced the reduction caused by LPS intraperitoneal injection (149.82, 178.62, and 179.26 pg/mL in low, middle, and high doses of PLCP group, respectively) (Fig. 4). Accordingly, our results indicated that PLCP enhanced antioxidant activities through increasing MT expression.
Antioxidant enzymes also play central roles in the defense against oxidative stress. GSH-Px is one of the potential antioxidant enzymes that defend against ROS. In our study, mice in LPS group showed a low activity of GSH-Px compared with the control group. PLCP significantly restored the activity of GSH-Px in LPS-administered mice (Fig. 2). In addition, mitochondria are the main site of ROS synthesis, which makes mitochondria very sensitive to oxidative stress. MnSOD, distributed in mitochondria, is one subpopulation of superoxide dismutase (SOD). It was reported that overexpression of mitochondrial RNS inhibits MnSOD activity. 38 It played an important role in liver tissue damage of rats with rheumatoid arthritis. 39 Zhang 40 demonstrated that LPS dramatically decreased the activity of MnSOD. In our study, a decreased MnSOD activity after LPS intraperitoneal injection was found, whereas PLCP administration significantly improved this decline (Fig. 2). These results suggested that after LPS challenge there may be increased ROS in the liver, whereas PLCP maintained the level of endogenous antioxidants in the liver of LPS challenged mice possibly by its antioxidant activity. These results were consistent with that of our previous research, which demonstrated that PLCP was able to significantly scavenge DPPH radicals, superoxide radicals, and hydroxyl radicals. 16 Thus, the protection shown by PLCP in this study might be attributed to its property of free radical scavenging and improvement of the endogenous antioxidant enzymes.
In conclusion, our results indicated that administration of PLCP downregulated overexpression of inflammatory cytokines (TNF-α, IL-6, IL-10, IL-2, and IL-1β) in serum and liver, decreased NO expression, and improved antioxidant activities in liver by improving GSH-Px and MnSOD activity. In addition, we demonstrated that PLCP inhibited iNOS and COX-2. Consequently, PLCP may act as a potential agent to prevent liver injury. Of course, further and comprehensive researches are needed to explore its mechanisms.
Footnotes
Acknowledgments
This study was supported by the National Key R&D Program of China under Grant No. 2017YFF0207800, National Natural Science Foundation of China under Grant Nos. 31860471 and 31260364, Natural Science Foundation of Jiangxi Province under Grant No. 20161BAB204171, and the Project of College Student Innovation of Nanchang University under Grant No. cx2016007.
Author Disclosure Statement
No competing financial interests exist.