
Abstract
Tendinopathy, a common disorder in man and horses, is characterized by pain, dysfunction, and tendon degeneration. Inflammation plays a key role in the pathogenesis of tendinopathy. Tendon cells produce proinflammatory molecules that induce pain and tissue deterioration. Currently used nonsteroidal anti-inflammatory drugs are palliative but have been associated with adverse side effects prompting the search for safe, alternative compounds. This study determined whether tendon-derived cells' expression of proinflammatory cyclooxygenase (COX)-2 and production of prostaglandin E2 (PGE2) could be attenuated by the combination of avocado/soybean unsaponifiables (ASU), glucosamine (GLU), and chondroitin sulfate (CS). ASU, GLU, and CS have been used in the management of osteoarthritis-associated joint inflammation. Tenocytes in monolayer and microcarrier spinner cultures were incubated with media alone, or with the combination of ASU (8.3 μg/mL), GLU (11 μg/mL), and CS (20 μg/mL). Cultures were next incubated with media alone, or stimulated with interleukin-1β (IL-1β; 10 ng/mL) for 1 h to measure COX-2 gene expression, or for 24 h to measure PGE2 production, respectively. Tenocyte phenotype was analyzed by phase-contrast microscopy, immunocytochemistry, and Western blotting. Tendon-derived cells proliferated and produced extracellular matrix component type I collagen in monolayer and microcarrier spinner cultures. IL-1β-induced COX-2 gene expression and PGE2 production were significantly reduced by the combination of (ASU+GLU+CS). The suppression of IL-1β-induced inflammatory response suggests that (ASU+GLU+CS) may help attenuate deleterious inflammation in tendons.
Introduction
Tendinopathy is a common musculoskeletal disorder characterized by pain, dysfunction, tendon extracellular matrix (ECM) degeneration, and rupture. 1 –6 Both man and horses suffer from this disorder as they share similar tendon structure, cellular-molecular characteristics, and biomechanical function. 1 –6 The prognosis for severe tendinopathy is typically poor, and effective strategies for treatment are limited. 7 –16 It has been suggested that the pathology is triggered by microdamage that accumulates in tendon ECM due to overuse, overload, and aging. 4,8,16 This accumulation could induce the upregulation of inflammatory and degradative pathways. How these processes contribute to tendon degeneration and rupture is not known. While inflammatory cell infiltrates such as macrophages are not a prominent feature in tendinopathy, the presence of inflammatory mediators, including prostaglandin E2 (PGE2), its cyclooxygenase (COX)-2 regulatory enzyme, cytokines, and metalloproteinases, has been documented. 10 –13
Inflammation has been proposed to play a role in tendon pathology as well as in healing and repair. 17 –21 In the early stage of naturally occurring equine tendon injury, inflammation at the cellular level has been identified to be important in healing. 17 –21 Similar observations during early-stage human tendon injury also implicate the role of inflammatory response in tendon repair. 17 –21 Increased levels of PGE2 and cytokines, such as interleukin (IL)-1β, have also been shown in the early response to tendon injury. In dogs, flexor tendon healing was characterized by dramatic upregulation of cytokine IL-1β expression. 20 However, persistent, unresolved inflammation with sustained excessive production of mediators, degradative enzymes, and growth factors could result in further damage and aberrant fibrotic repair. 8 –21 An imbalance in the levels of inflammatory mediators and proteolytic enzymes could lead to progressive tendon ECM deterioration, impaired healing, and rupture.
The prominent role of COX-2 and PGE2 in tendon pathology led to the widespread use of analgesic and nonsteroidal anti-inflammatory drugs (NSAIDs). However, safety concerns have been raised regarding an increased risk of re-injury with prolonged use of NSAIDs. There is evidence suggesting that NSAIDs that act as COX-2 inhibitors could hinder the healing process in tendons. 14–15,22 These drugs have also been associated with serious side effects, including gastrointestinal, renal, and liver pathologies. 23 The drawbacks of NSAIDs have prompted a search for effective alternatives to either replace or supplement these agents.
A number of nonpharmacological agents have shown considerable anti-inflammatory potency along with a good safety profile. Among these products are avocado/soybean unsaponifiables (ASU), glucosamine (GLU), and chondroitin sulfate (CS) used separately or in combination. GLU has been shown to effectively reduce pain and increase mobility. 24 CS has shown efficacy in long-term treatment of inflammation and pain. 25,26 In vitro, the combination of CS and GLU effectively suppressed inflammation in equine tendon and tendon cells. 27,28 ASU, GLU, and CS have been demonstrated to have anti-inflammatory activity in vitro. 27 –39 Clinical studies in man and animals support the beneficial and safe administration of these agents. 38 –49 A recent review reported that nonpharmacologic compounds, including GLU and CS, are commonly used as support therapy for tendinopathies. 48 Preclinical results on the use of these agents are encouraging, and further confirmation by clinical studies is being pursued. 46,50,51 To what extent these agents may be used as substitutes or supplements of NSAIDs for the management of tendinopathies remains to be explored.
The search for alternative compounds to replace or supplement NSAIDs for the management of tendinopathy would be facilitated by the availability of culture methods that recapitulate repetitive mechanical stress that tenocytes encounter in vivo. Static monolayer culture techniques do not expose tendon-derived cells to mechanical forces with associated changes in cell markers. 52,53 To address this unmet need, we used dynamic microcarrier spinner culture method to help identify potential therapeutic agents. The microcarrier spinner culture method has been employed successfully to propagate chondrocytes, osteoblasts, and tenocytes for cell response analyses and tissue engineering applications. 39,53 –60 Using both the monolayer and the microcarrier spinner culture methods, we found that the combination of (ASU+GLU+CS) attenuated cytokine IL-1β-induced COX-2 expression and PGE2 production in tendon-derived cells.
Materials and Methods
Isolation and propagation of tendon-derived cells
Digital flexor tendons were obtained from three adult mares (8–24 years), scheduled for the Institutional Animal Care and Use Committee. Cells were isolated by tendon digestion with type II collagenase (110 U/mL; GIBCO®; Invitrogen Corporation, Carlsbad, CA, USA) at 37°C, 5% CO2 as previously described. 30 The viability of retrieved cells was assessed using the trypan blue exclusion method. Tendon-derived cells were propagated to confluency in monolayer for 2–4 weeks and harvested using trypsin EDTA (Invitrogen). Cells (4 × 106) were re-seeded in microcarriers and cultured in spinner flasks (Bellco Glass, Vineland, NJ, USA) as previously described. 55
Characterization of tendon-derived cells in monolayer and microcarrier spinner cultures
Cell imaging
Cells were visualized by phase-contrast microscopy, and images were captured using an Olympus C-7070 Wide Zoom digital camera mounted on an Olympus CKX41 inverted phase-contrast microscope.
Immunocytochemistry
Cells were detached from microcarriers by digestion with collagenase as previously described. 54 Retrieved cells were resuspended at 1 × 105 cells/mL and re-plated on Thermanox™ discs for 24 h. Cells were fixed in 10% formalin for 15 min, washed three times with phosphate buffered saline (PBS) and were incubated overnight at 4°C with a 1:1000 dilution of goat anti-bovine type I collagen (Southern Biotechnology Associates, Birmingham, AL, USA) in PBS containing 0.1% Triton-X-100. After washing four times with PBS, cells were incubated with a 1:100 dilution of Alexa-Fluor 488 donkey anti-goat IgG (Invitrogen) overnight at 4°C, washed with PBS, and viewed using a TX200 Nikon fluorescence microscope equipped with a Nikon digital camera.
Western immunoblot analysis
Equal amounts of proteins (10 μg) from culture supernatants were run on 4–15% polyacrylamide gels under reducing conditions and were transferred electrophoretically onto nitrocellulose membranes (BioRad Laboratories, Hercules, CA, USA) in Tris-glycine buffer containing 20% methanol. The membranes were washed twice with deionized water and probed using a chromogenic western blot immunodetection kit (Invitrogen Corporation) containing a blocking solution to eliminate nonspecific staining. Membranes were incubated with goat anti-collagen type I antibody (1:1000; Southern Biotechnology Associates) followed by alkaline phosphatase-labeled rabbit anti-goat antibody and BCIP/NTB as substrate (Invitrogen Corporation).
Treatment design
ASU (NMX1000®; Nutramax Laboratories, Inc., Edgewood, MD, USA) formulation was dissolved in 200 proof ethanol (Sigma Aldrich) to obtain the desired concentration. The clinically relevant concentrations of ASU used for this study were selected following testing on articular chondrocytes. 30,31 ASU composition analysis was performed using gas chromatography/mass spectrometry and high-pressure liquid chromatography. 31 GLU (FCHG49®; Nutramax Laboratories, Inc.,) and CS (TRH122®; Nutramax Laboratories, Inc.,) were dissolved in Hanks balanced salt solution (ATCC) to obtain the desired working concentration. The concentrations of ASU (8.3 μg/mL), CS (20 μg/mL), and GLU (11 μg/mL) selected were drawn from previous in vivo and in vitro studies. 24 –50 Detectable concentrations of GLU and CS in plasma after oral administration showed biologic effects. Although the bioavailability of ASU is unknown, oral doses used in man and horse have exhibited anti-inflammatory and disease-modifying effects. Thus, reported physiologically relevant concentrations of ASU, GLU, and CS were used in the present study. 24 –50 There were no toxic effects of the (ASU+GLU+CS) concentrations used in vitro and in vivo. 24 –50
Cells in 24-well monolayer cultures (seeded 1 × 105/mL overnight) and cell-seeded microcarriers in 10 mL of fresh medium in minispinner flasks were used for treatment experiments. Both sets of tendon cultures were incubated with medium alone, or a combination of ASU (8.3 μg/mL), CS (20 μg/mL), and GLU (11 μg/mL) for 24 h at 37°C, 5% CO2. Two sets of monolayer and microcarrier cultures were next incubated with media alone, or stimulated with IL-1β (10 ng/mL; R&D systems). One set was incubated for 1 h to analyze COX-2 expression, while the second set was incubated for another 24 h to measure PGE2 production. Culture supernatants from monolayer and microcarrier cultures were collected after 24 h for PGE2 immunoassay. Cells from monolayer cultures were retrieved by trypsinization, while pelleted cell-seeded microcarriers were enzyme-digested after 1 h for real-time polymerase chain reaction (PCR) analysis as previously described. 30
COX-2 expression analysis using quantitative PCR
Cells were lysed using TRizol® reagent (Invitrogen), followed by RNA extraction using chloroform (Sigma-Aldrich) as previously described. 30 The synthesis of cDNA from 1 μg of total tenocyte RNA was performed using Moloney-Murine Leukemia Virus reverse transcriptase from the Advantage RT-for-PCR kit (BD Biosciences Clontech, Mountain View, CA, USA). 30 Primers specific to equine COX-2 (forward, ATA CCA AAA CCG CAT TGC CG; reverse, TCT AAC TCC GCA GCC ATT TC) and equine GAPDH (forward, GTT TGT GAT GGG CGT GAA CC; reverse, TTG GCA GCA CCA GTA GAA GC) as the housekeeping gene were used for gene expression analysis. 30 PCR products were generated using 2 μg of cDNA sample template and iQTM SYBR Green Super Mix, 2 × mix for real-time PCR (BioRad, Hercules, CA, USA). Denaturation of cDNA was performed as previously described. 30 Incorporated dye was detected using the BioRad iQ5 real-time detection system to determine the threshold cycle (CT), as described by the iQ5 manual (BioRad). ΔΔCT values were calculated from CT values, and a graphical representation of the data was generated to observe fold differences in copy number as previously described. 30
Measurement of PGE2 production using high-sensitivity immunoassay
A commercial PGE2 immunoassay was used to quantify secreted PGE2 levels in the cellular supernatant, according to the manufacturer's instructions (R&D). A PGE2 standard was run in parallel to the supernatant samples. Briefly, 100 μL of each supernatant sample was assayed in triplicate on a 96-well microplate coated with a goat anti-mouse polyclonal antibody as previously described. 30
Statistical analysis
The SigmaStat statistical program (Windows version 3.11; Systat Software Inc., Point Richmond, CA) was used for data analysis, with multiple comparisons by one-way analysis of variance (ANOVA) using the Tukey post hoc test. Differences of P < .05 were considered statistically significant.
Results
Characterization of tenocytes in monolayer and microcarrier spinner culture
Tendon-derived cells plated in monolayer culture were mostly spindle-shaped displaying elongated morphology. Doubling times were slow in early passage (∼7–14 days) and increased (∼5–7 days) by passage three (Fig. 1A). Tenocytes retrieved from monolayer culture after three to four passages readily attached to microcarrier beads during the initial intermittent stirring phase of the seeding process. By day 7, most microcarrier beads exhibited adherent cells on their surface. These cells proliferated reaching confluency in 10–14 days (Fig. 1B). Tenocyte-seeded microcarriers began to aggregate within 5 days of incubation and exhibited ECM-like material between adjacent beads. Empty unseeded microcarriers did not form aggregates. Tenocytes on the surface of microcarriers displayed heterogeneous morphology with a fibroblastoid or spherical shape. By 2 weeks, larger aggregates were formed and ECM-like material appeared to bridge large clusters together (Fig. 1B).
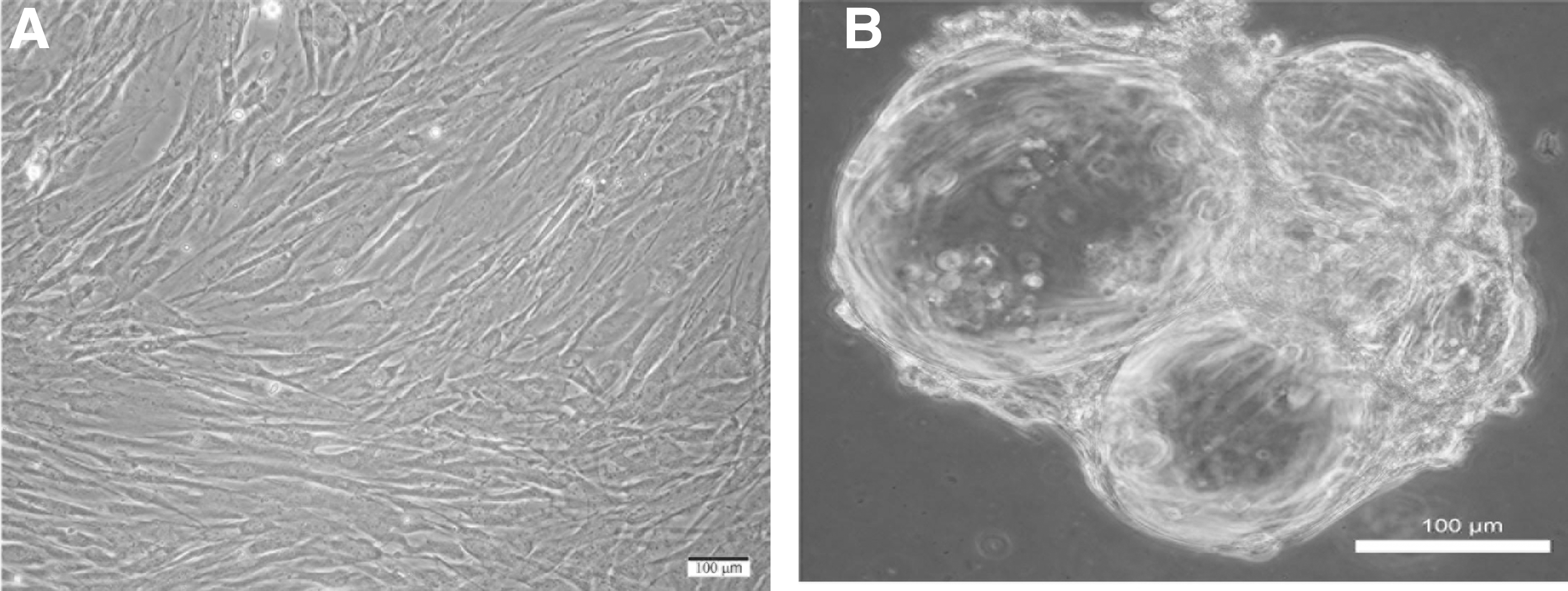
Representative phase-contrast micrographs:
Tenocytes retrieved from microcarriers remained viable and attached with ease when re-plated onto Thermanox discs. Cells in monolayer and those recovered from microcarriers displayed immunostaining for type I collagen (Fig. 2A, B). Immunostaining revealed distinct localization of the collagen protein in the cytoplasm indicated by green fluorescence, while the nucleus displayed blue DAPI staining (Fig. 2A, B). The specificity of immunostaining was verified when the primary or secondary antibody was omitted, or when the primary antibody was substituted with normal immunoglobulin. In addition, the antibody against type I collagen did not stain other cell types such as monocytes-macrophages that do not produce type I collagen.
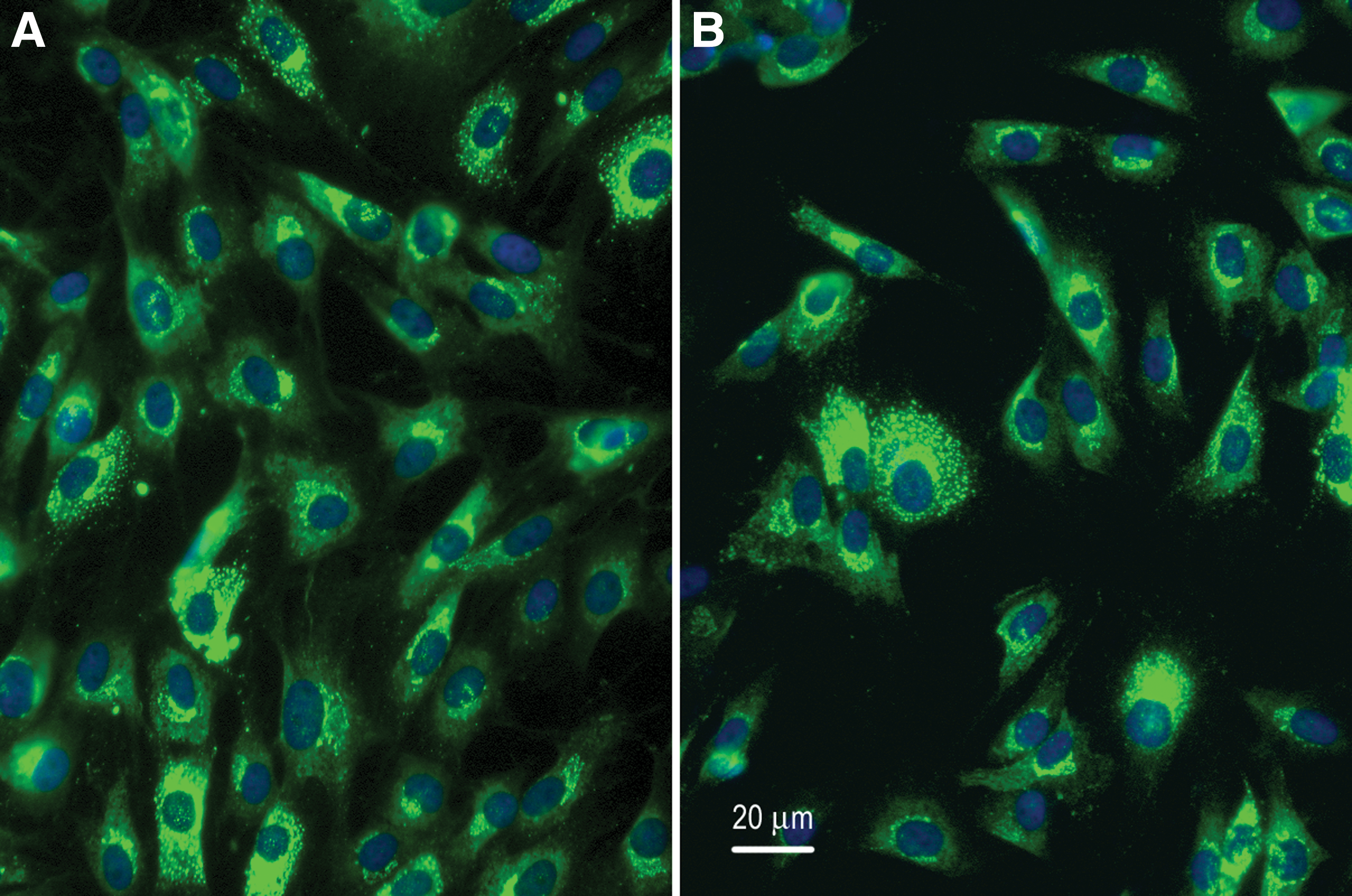
Representative photomicrographs of tenocytes retrieved from monolayer and microcarrier spinner cultures immunostained with antibody against type I collagen. Type I collagen stained green and nuclei stained blue with DAPI. Scale bar = 20 μ.
The production and secretion of type I collagen by tenocytes grown on monolayer and microcarrier cultures were further verified by western blot analysis. Supernatants from tenocytes in microrcarrier culture showed continued production of type I collagen (Fig. 3, left panel) with similar patterns to supernatant from monolayer culture (Fig. 3, right panel). Immunoreactive protein bands were seen between 115 and 202 Kd (Fig. 3). There was no detectable immunostaining of type I collagen in supernatants from microcarrier cultures without cells.
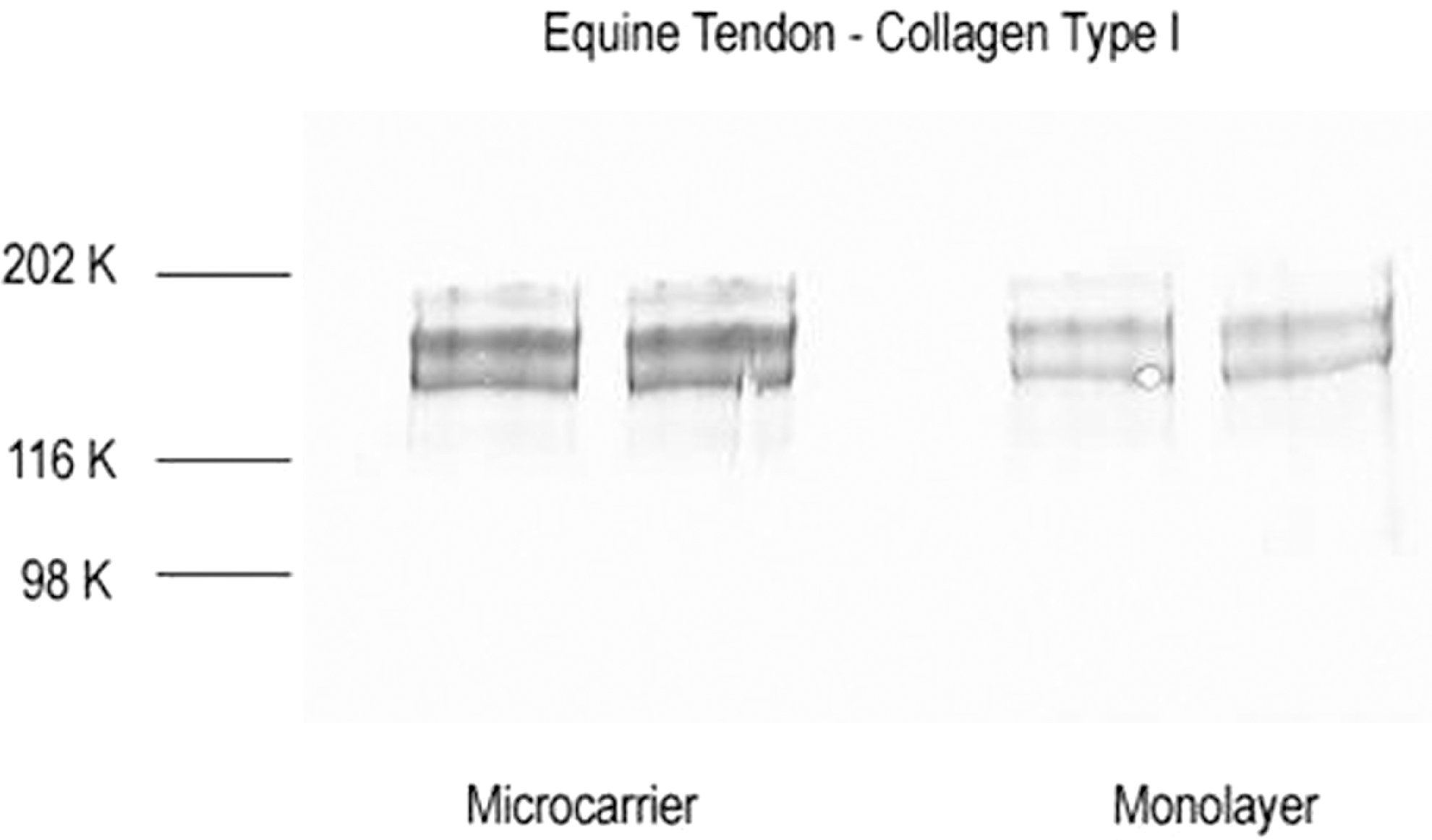
Representative western blots immunostained for type I collagen of supernatants from monolayer and microcarrier spinner cultures. Duplicate samples from microcarrier culture (left panel) and monolayer culture (right panel) were used for immunoblots. Immunoreactive collagen bands were seen between 115 and 202 Kd.
Effect of (ASU+GLU+CS) combination on COX-2 gene expression and PGE2 production in tenocyte monolayer and microcarrier spinner culture
Nonactivated tenocytes in monolayer and microcarrier cultures incubated for 1 h with media alone (C) expressed lower levels of COX-2 mRNA, whereas cells activated with IL-1β showed greater COX-2 expression (Fig. 4A, B). For ease of comparison, results were expressed as a percentage of IL-1β-activated COX-2 expression (100%). Pretreatment with the combination of (ASU+CS+GLU) profoundly reduced the induction of COX-2 expression by IL-1β in both monolayer and microcarrier spinner cultures (Fig. 4A, B). Table 1 shows significant increase in COX-2 expression upon activation of monolayer cultures with IL-1β (P < .0001). (ASU+CS+GLU) treatment significantly reduced COX-2 expression in IL-1β-activated cells (P < .0001). In contrast, COX-2 expression in control nonactivated microcarrier cultures displayed wide variability and did not significantly increase with IL-1β activation (P > .05, Table 1). In spite of the variability, the (ASU+CS+GLU) combination significantly reduced COX-2 expression in IL-1β-activated cells (P < .03).

Representative effect of (ASU+GLU+CS) on IL-1β-induced COX-2 gene expression in tenocyte monolayer and microcarrier spinner cultures. Data taken from one tenocyte cell line. Gene expression analysis in equine tenocytes shows a profound elevation in COX-2 levels with IL-1β stimulation. (ASU+GLU+CS) dramatically reduced COX-2 gene expression in
Effect of (ASU+GLU+CS) on IL-1β-induced COX-2 Gene Expression in Monolayer and Microcarrier Spinner Cultures
Data represent the mean ± standard deviation (n = 3 per group).
(ASU+GLU+CS) treatment significantly reduced IL-1β-induced COX-2 gene expression in monolayer and microcarrier spinner cultures.
ASU, avocado/soybean unsaponifiables; COX, cyclooxygenase; CS, chondroitin sulfate; GLU, glucosamine; IL, interleukin.
Tenocytes incubated for 24 h with control media alone (C) showed low levels of PGE2, whereas activation with cytokine IL-1β significantly increased PGE2 production in monolayer and microcarrier cultures (P < .001, Fig. 5A, B). Pretreatment of tenocytes in both monolayer and microcarrier spinner cultures with the combination of (ASU+GLU+CS) significantly inhibited IL-1β-induced PGE2 production (P < .001; Fig. 5A, B). The combination product suppressed but did not totally block PGE2 in both monolayer and microcarrier cultures (P < .001; Fig. 5A, B).

Effect of (ASU+GLU+CS) on IL-1β-induced PGE2 production in tenocyte monolayer and spinner cultures. Results are presented as mean ± 1 standard deviation, n = 3. Tenocyte PGE2 production increased with IL-1β stimulation and was significantly reduced with (ASU+GLU+CS) treatment in both
Discussion
The key finding of the present study is that tendon-derived cells in monolayer culture respond to IL-1β stimulation with enhanced COX-2 expression and robust production of PGE2. This inflammatory response is significantly inhibited by the combination of (ASU+GLU+CS). Although, there is wide variability in COX-2 expression in tenocyte microcarrier spinner culture, (ASU+GLU+CS) also significantly reduced both COX-2 expression and PGE2 production. Tendon is comprised mostly of collagen type I ECM infiltrated with tenocytes and fibroblast-like cellular elements. It has been thought that tenocytes have the capacity to heal the tendon intrinsically, and much effort has been devoted to study the behavior of these cells. 1–2,18,21,52,53 Tenocytes propagated in monolayer culture are commonly used for such studies, but cells retrieved from tendon are limited in number. They also undergo phenotypic drift with increasing passage, raising doubts as to whether cells propagated in monolayer represent an appropriate in vitro model for studying tendon biology. 52,53 Tendon explant cultures have also been used as models, but their utility is hampered by limited survival times and problems with storage. 27
Since biomechanical forces influence tenocyte behavior, culture techniques that apply mechanical forces, such as strain and fluid shear, would facilitate the analysis of their function and metabolism. 1 –4 In this study, we used microcarrier spinner cultures to monitor the response of tenocytes exposed to fluid shear with respect to their expression of COX-2 and production of PGE2 under dynamic conditions. This culture method facilitates the attachment of cells onto a constrained microcarrier surface and exposes cells to biomechanical fluid shear forces during suspension culture. 54 –60 Thus, the microcarrier spinner culture provides an environment that more closely reflects the conditions found in situ. The current study confirms that equine tenocytes grow with ease in collagen microcarrier beads, produce ECM type I collagen, and can be activated to express COX-2 and produce PGE2 (Figs. 1–5, Table 1). These results indicate that the dynamic tenocyte spinner culture may be a useful in vitro model for screening and evaluating agents that could modulate deleterious proinflammatory events associated with tendinopathy.
PGE2 and COX-2, its key regulatory enzyme, were used in this study as well-defined markers of inflammation and documented mediators in the pathogenesis of tendonitis. 8 PGE2 is known to induce the production of degradative enzymes as well as suppress the production of extracellular matrix components. In addition, PGE2 induces the synthesis of other proinflammatory molecules, thus exacerbating the inflammatory response and damage to tendon. 8 –13 The elevated production of PGE2 has been shown to affect cell proliferation and the production of ECM, including catabolism by matrix metalloproteinases, a family of enzymes responsible for degrading ECM, and the synthesis of type I collagen. 8 –13 The pathologic effect of these inflammatory molecules is associated with localized degenerative changes in tendons. 8 –13 Collectively, these studies indicate a relationship between inflammatory mediators, degradative enzymes, induction of pain, and pathologic changes in tendon. 5 –13
The observation that (ASU+GLU+CS) significantly suppressed but did not totally block COX-2 expression and PGE2 production supports the notion that this combination could tone down inflammation without abrogating their physiologic functions (Figs. 4, 5, and Table 1). We have earlier reported similar inhibition of cytokine-induced PGE2 production in equine monolayer and canine microcarrier chondrocyte cultures by (ASU+GLU+CS) combination. 37,39 PGE2 with other inflammatory molecules are reported to be critical for healing and tissue repair. 17 –21 (ASU+GLU+CS) could spare the physiologic functions of PGE2 while attenuating excessive and extended production of PGE2. The inhibition of PGE2 would attenuate the production of degradative enzymes and maintain the synthesis of tendon ECM components. (ASU+GLU+CS) could also reduce sensitization of pain nociceptors by PGE2.
NSAIDs are widely used to counteract the increased production of PGE2 and associated biological molecules in patients suffering from tendinopathy. 14 –25 However, there is concern that these drugs may impede the early phases of the healing process and inhibit collagen synthesis. Our results show that PGE2 production by tenocytes in monolayer and spinner culture is significantly reduced by pretreatment with (ASU+GLU+CS). This reduction is most likely linked to the inhibition of COX-2 expression since mRNA levels of this enzyme are also reduced. In conclusion, the finding that COX-2 expression and PGE2 production are attenuated by (ASU+GLU+CS), but were not totally abrogated, is clinically important since inflammatory molecules such as PGE2 are thought to play a role in tendon healing and repair. 17 –21 (ASU+GLU+CS) may mitigate deleterious inflammation without ablating PGE2 synthesis needed for physiologic tendon healing and repair. Further in vivo studies would help clarify the effects of (ASU+GLU+CS) and assess its clinical efficacy.
Footnotes
Acknowledgments
This study was supported by Nutramax Laboratories Veterinary Sciences, Inc. We would like to thank Dr. Reinhard Grzanna for designing the immunohistochemistry and western blot experiments as well as for constructive review of the article.
Author Disclosure Statement
C.G.F., R.Y.A., A.Y.A., and M.W.G. are former employees of Nutramax Laboratories Veterinary Sciences, Inc., but do not hold stocks or royalties. A.M.R. does not have a conflict of interest.