
Abstract
Opuntia ficus indica has been an important dietary source and a traditionally used medicinal plant. Given the promising health-promoting properties of this plant, a comparative toxicological assessment and antioxidant bioevaluation of extracts from different parts of the plant were carried out in relation to their chemical profile. Toxicity was examined at multiple endpoints using the 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT), Comet and the γH2AX In-Cell Western Assay, while hyphenated ultra-high-performance liquid chromatography-high-resolution mass spectrometry (UHPLC-HRMS) analysis was carried out to identify main constituents. None of the extracts showed any cytotoxic and genotoxic effect on cell lines used, apart from the flower extract in HepG2 cells at the highest concentration tested (2.5 mg/mL). Both fruit flesh and seed extracts demonstrated a prominent protective effect against H2O2-induced genotoxicity in almost all concentrations tested, while extracts originated from flowers and cladodes were effective only at the low non-cytotoxic (0.312 and 0.625 mg/mL) and high (1.25 and 2.5 mg/mL) concentrations, respectively. In total, 2 phenolic acids, 12 flavonoids, along with 3 feruloyl derivatives and the plant pigment indicaxanthin, were tentatively identified by UHPLC-HRMS analysis. Phenolic acids (compounds
Introduction
Opuntia ficus indica is a tropical and subtropical plant that belongs to the Cactaceae family. 1 It grows in arid and semiarid climates with a global distribution, including Mexico, Latin America, South Africa, and the Mediterranean. 2 O. ficus indica (OF) has been a traditionally used herbal medicine around the world, with parts of the cactus plant being used for their emollient, moisturizing, cicatrizing, hypocholesterolemic, antiulcerogenic, and hypoglycemic properties. 3 –5 As a diet component, cactus pear fruit and its products have been long consumed for their considered high nutritional value in Latin America and worldwide later. 1
The last decades, OF has drawn considerable attention regarding the health-promoting and pharmacological activities that it presents. 1,2,6 Numerous studies have supported this notion by determining the presence of significant amounts of phenolic acids and several flavonoids, including isorhamnetin, kaempferol, and quercetin glycosides identified in extracts obtained from flowers, cladodes, fruits, and seeds. 6 –15 Beside phenolics, substantial amounts of vitamin E, 16 ascorbic acid, 8 betalains, 17 and taurine 6,18 have been reported as well.
The antioxidant potential of OF is widely supported by several in vivo and clinical studies. Extracts from cladodes have shown to prevent oxidative damage endpoints induced by the mycotoxin zearalenone in the liver and kidney in Balb/C mice in vivo and to decrease hepatic oxidative stress biomarkers in obese Zucker rats after being fed a cladode-containing diet. 19,20 Similarly, fruits of OF have shown promising results against the ethanol-induced gastric ulcer in rats, a protection partially attributed to the antioxidant activity of isorhamnetin derivatives. 21 Moreover, a short-term dietary supplementation with fruit flesh positively affected the body's redox balance, decreased lipid oxidation, and improved antioxidant status of healthy volunteers. 22
Several other beneficial proprieties have been reported for OF in in vivo and in vitro conditions, including a hypoglycemic, hypolipidemic, anticancer, and neuroprotective effect with the underlying mechanisms of action still being under investigation. 23 –27
Given the promising biological activities that all parts of OF have, mostly associated with their strong antioxidant activity, there is a growing interest for their medicinal and food supplementation use nowadays. In this work, a comparative study was carried out on the chemical content of five aqueous extracts obtained from the different aerial parts of the plant. In respect to their profile, the extracts were comparatively examined for cytotoxic and genotoxic effects, as well as for their potential protective activity against H2O2-induced DNA damage using a selected human cell line.
Materials and Reagents
Chemicals and reagents
All chemicals and reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA), unless stated otherwise. Triton X-100 was obtained from Serva, propidium iodide from Research Organics, ethanol from Merck, SYBR Gold from Life Technologies, and hydrogen peroxide solution from Carlo Erba. PHOSSTOP for the γH2AX In-Cell Western assay was purchased from Roche, while MAXBlock Blocking Medium from Active Motif. Primary rabbit monoclonal anti-γH2AX (Clone 20E3) was supplied by Cell Signaling, while secondary goat anti-Rabbit IgG CF770 and RedDot2 were purchased from Biotium. LC-MS grade acetonitrile (ACN), water (H2O), and formic acid were purchased from Fisher Scientific Co.
Extraction of plant material
Plant material was obtained from local producers of the southern Peloponnese (municipality of East Mani, Laconia, Greece) in June 2014. Five different extracts were obtained from the aerial parts of OF. Namely, OF1, OF2, OF3, OF4, and OF5 extracts were received from OF flower, cladode, fruit peel, fruit seed, and fruit flesh, respectively. Extractions were carried out in H2O using a Branson 2510 Ultrasonic bath. Originally, 1 g of dried and finely chopped peels, seeds, and flowers and 5 g of the fresh flesh and dried cladodes were extracted with 50 mL of distillated water each. Following the end of the 45-min extraction, samples were centrifuged at 3000 rpm for 15 min using a Heraeus Multifuge 3S centrifuge and filtered through a Buckner funnel using filter paper. Obtained aqueous extracts were then stored at −20°C for further UHPLC-HRMS analysis and biological evaluation.
Chemical analysis
Identification and comparative analysis of secondary metabolites were performed on an ultra-high-performance liquid chromatography system hyphenated to a hybrid LTQ Orbitrap Discovery XL high resolution mass spectrometer (UHPLC-HRMS) as previously described. 28 Before injection, each extract was adjusted to the final concentration of 50 μg/mL with H2O. Comparative analysis was based on EIC (extracted ion chromatogram) areas. All experiments were performed in triplicate.
Determination of total phenolic and total flavonoid content
The total phenolic content (TPC) was determined based on the Folin-Ciocalteu colorimetric method with few modifications. 29 Briefly, 25 μL of each extract in DMSO (1 g/L) was mixed with 125 μL of Folin-Ciocalteu's reagent (10% v/v). After mixing, 100 μL of aqueous sodium bicarbonate (75% v/v) was added, and the mixture was allowed to stand for 30 min in dark. The absorbance was measured at 765 nm. TPC was expressed in mg gallic acid equivalents (GAE)/g of dried extract. Determination of total flavonoid content (TFC) was based on the aluminum chloride colorimetric method. 30 TFC was expressed in mg quercetin equivalents (QE)/g of dried extract. All assays were carried out in triplicate.
Cell culture
HepG2 and HeLa cells were cultured in Minimum Essential Medium Eagle and Dulbecco's modified Eagle's medium, respectively, supplemented both with 10% v/v FBS and antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin). Both cell lines were maintained at 37°C in a humidified incubator with 5% CO2 and 95% air.
Tetrazolium-based colorimetric 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide assay
The 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay was performed as previously described with some modifications. 31 Briefly, HepG2 and HeLa cells were seeded in 96-well plates and incubated at 37°C. After 24 h, cells were exposed to serum-free medium containing the OF1-OF5 extracts at concentrations ranging from 0.005 to 2.5 mg/mL for 24 h at 37°C. Five percent DMSO and H2O were used as positive and negative controls, respectively. Before the end of the exposure phase, the medium was removed and replaced with 1 mg/mL MTT solution. After 4 h of incubation, the absorbance was measured colorimetrically at 550 nm using the plate reader BioRad i-Mark.
Single cell gel electrophoresis assay (Comet assay)
The single cell gel electrophoresis assay was performed as previously described with some modifications. 32 –34 Briefly, HepG2 cells were seeded in a 12-well plate. After 24 h, cells were treated with OF1–OF5 extracts at a concentration of 2.5 mg/mL for 24 h. H2O and H2O2 were used as negative and positive controls, respectively. After trypsinization, cells were mixed with 1% low melting point agarose in PBS and pipetted onto glass microscope slides. Two gels were prepared for each extract. The slides were immersed in prechilled lysis solution for 1 h at 4°C and placed in the electrophoresis buffer (300 mM NaOH and 1 mM Na2EDTA, pH ≥13) for 30 min. Neutralization in Tris 0.5 M (pH 7.4) and dehydration with absolute ethanol were followed. Slides were allowed to dry, stained with SYBR Gold, and examined with a LEICA DM2500 fluorescence microscope equipped with an excitation filter BP 515–560 nm. The incidence of DNA single-strand breaks in HepG2 cells was evaluated using the parameter “%DNA in the comet tail” analyzed with TriTek CometScore™ software v1.5.
γH2AX In-Cell Western assay
The γH2AX In-Cell Western (γH2AX ICW) assay was performed according to Khoury et al. 35 with some modifications. Briefly, HepG2 cells were seeded in a 96-well plate. After 24 h, cells were treated with the OF1–OF5 extracts at concentrations of 0.312, 0.625, 1.25, and 2.5 mg/mL for 24 h at 37°C. H2O and H2O2 were used as negative and positive controls, respectively. Cells were subsequently treated with H2O2 for 20 min when the potential protective effect of extracts on H2O2-induced genotoxicity was examined. After treatments, cells fixed with 4% paraformaldehyde in PBS and neutralized with 20 mM NH4Cl. Cells were then permeabilized, blocked with MAXblock blocking medium supplemented with phosphatase inhibitor PHOSSTOP and RNase A, and incubated with primary rabbit monoclonal anti-γH2AX in PBS-SFB-Triton X-100 buffer for 2 h.
After PST wash, cells were incubated with the secondary goat anti-Rabbit IgG-conjugated with an infrared fluorescent dye and the DNA labeling dye RedDot 2 in PST for 1 h. Determination of the fluorescence intensity for DNA and γH2AX was carried out by using the Odyssey Infrared Imaging Scanner (LiCor) at 700 and 800 nm. Genotoxicity was considered positive when the extracts produced a statistically significant 1.3-fold induction of γH2AX at a level of cytotoxicity below 50% compared to the negative control. These parameters were based on previous studies by Khoury et al. 31,35,36
Statistical analysis
GraphPad Prism 7.0 was used for the statistical analysis of both phytochemical and biological data. Results generated by the comparative analysis of different phytochemicals were evaluated by applying Tukey's multiple range test, and analyzed as mean ± standard deviation and fold change. Statistical significance of results was considered at P-values ≤0.05.
Results generated by the MTT and Comet Assay were analyzed with the nonparametric Kruskal–Wallis test followed by Dunn's post hoc test. For the γH2AX In-Cell Western assay, statistically significant increases in γH2AX phosphorylation after treatment were compared to the negative controls using Student's t-test with SPSS 16.0 for Windows (SPSS BI Greece SA) to estimate the genotoxic potential of the tested extracts. For the assessment of the protective effects of extracts against H2O2-induced genotoxicity, the levels of γH2AX phosphorylation were compared to the positive controls (H2O2-treated cells). P ≤ 0.05 was considered statistically significant.
Results
Phytochemical analysis and comparative investigation
Results of the UHPLC/HRMS analysis are shown in Table 1. Among identified phenolic acids, cladodes extract (OF2) showed the highest level of piscidic (

Structure of representative metabolites identified in different plant tissues of Opuntia ficus indica. Eucomic acid (
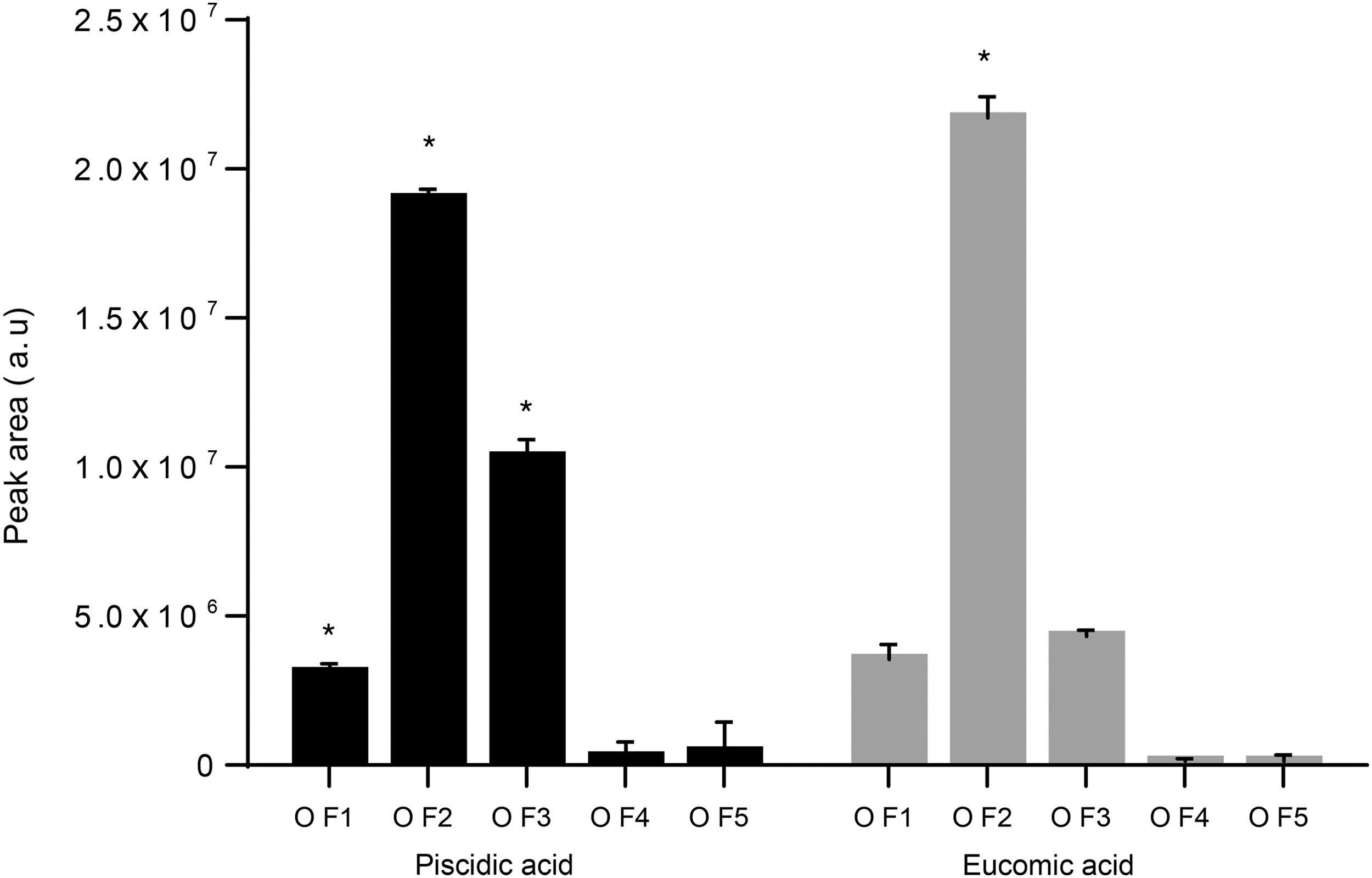
Distribution of phenolic acids in different plant tissues of O. ficus indica. Asterisk represents a significant different value of a specific compound among the different extracts (P < 0.05). OF1: flower, OF2; cladode, OF3; fruit peel, OF4; fruit seed, and OF5: fruit flesh.

Pie charts show the relative proportion of main phenolic acids (compounds
UHPLC-HRMS Analysis of Opuntia ficus indica: A Comparative Investigation of the Different Parts (Flower, Cladode, Fruit peel, Fruit seed, and Fruit Flesh)
The different plant parts were arranged according to the relative abundance of each compound.
Ara, arabinose; Glu, glucose; Rha, rhamnose; Xyl, xylose; ISO, isorhamnetin; K, kaempferol; Q, quercetin; RDB, ring double bond equivalent; OF1, flower; OF2, cladode; OF3, fruit peel; OF4, fruit seed; OF5, fruit flesh.
Among identified flavonoids, 81.8% of the total amount (compounds

Distribution of flavonoids in the different O. ficus indica plant tissues. Asterisk represents a significant different value of a specific compound among the different plant tissues (P < 0.05). OF1; flower, OF2; cladode, OF3; fruit peel.

UHPLC-(−)ESI/HRMS chromatogram of high abundant flavonoids identified in the flower extract of O. ficus-indica. Ion chromatogram extracted at m/z corresponding to the monoisotopic mass of detected flavonoids.
Total phenolic and flavonoid contents of the aerial parts
The total phenolic and flavonoid content of the aerial parts of OF, as defined by colorimetric methods, are presented in Figure 6. TFC ranged from 2.20 ± 0.024 mg of QE equiv/g in the peel (OF3) to 22.76 ± 0.015 mg of QE equiv/g in the flower extract (OF1). Flavonoids have not been detected in the seeds and in the flesh extracts. TPC varied from 5.03 ± 0.001 mg GAE/g in the flesh (OF5) to 61.44 ± 0.213 mg GAE/g in cladodes (OF2) and to 62.80 ± 0.009 mg GAE/g in the flower (OF1).
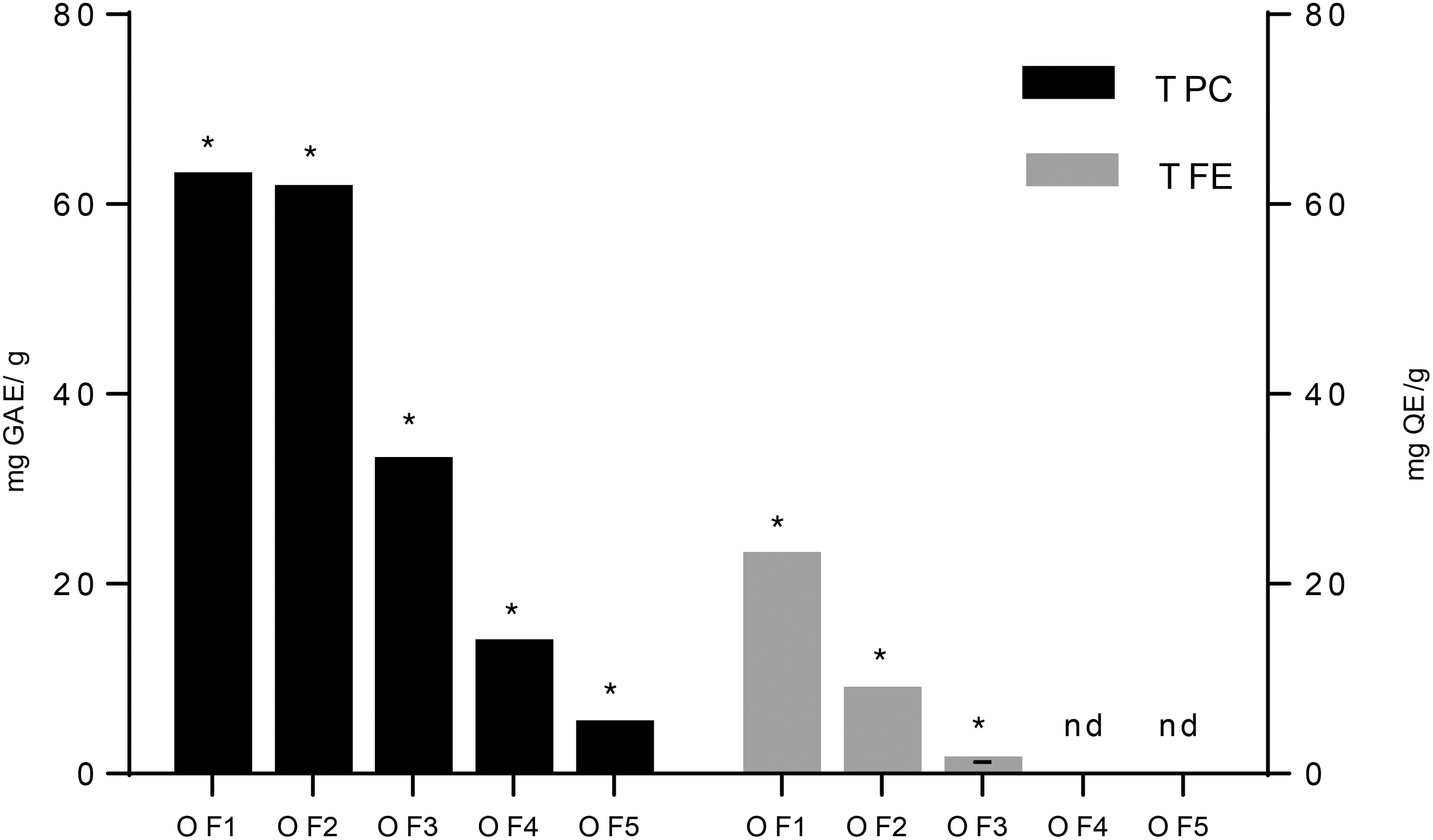
Total phenolic and flavonoid contents of O. ficus indica. (OF1: flower, OF2; cladode, OF3; fruit peel, OF4; fruit seed, and OF5; fruit flesh). Asterisks represent a significant different value.
In vitro assessment of cytotoxicity
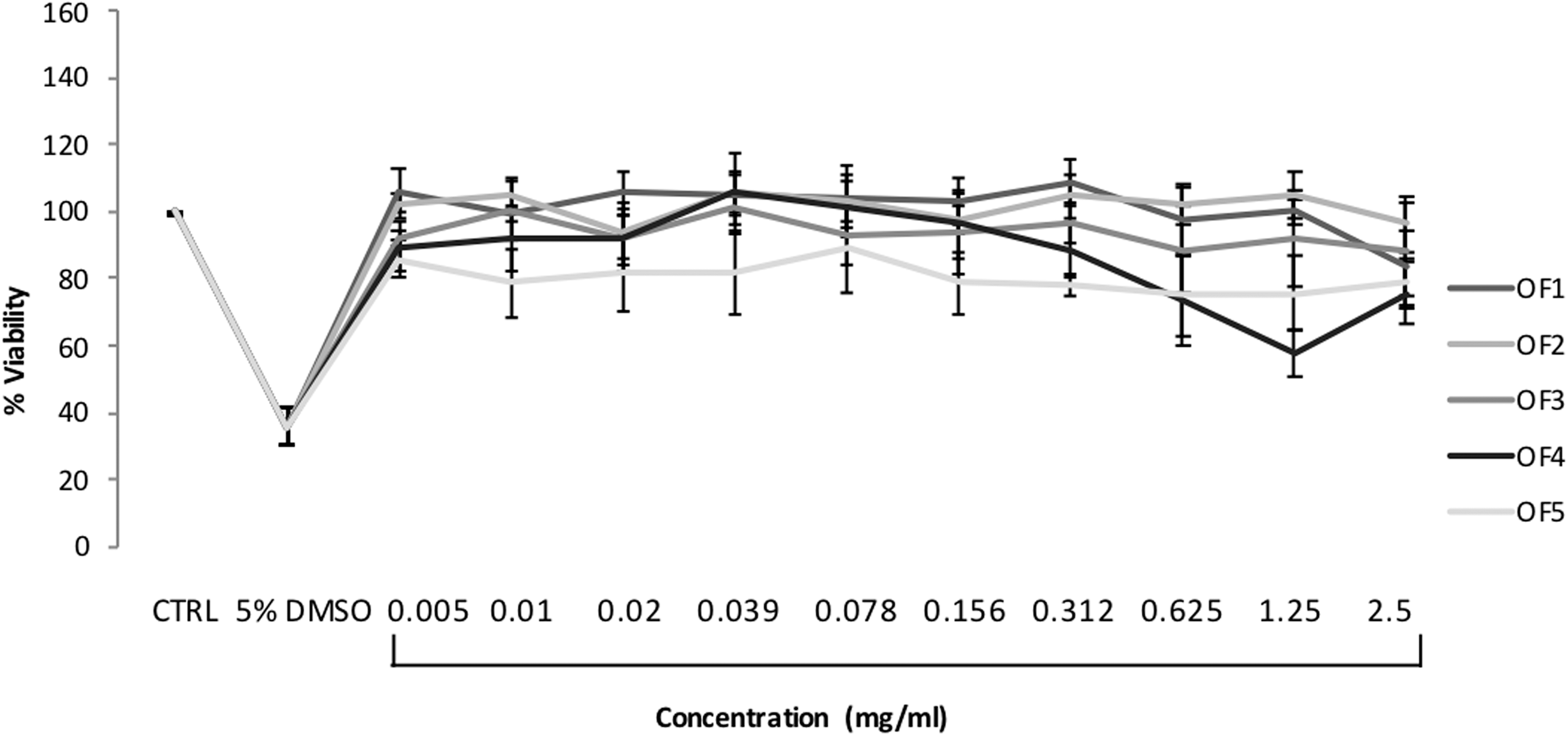
Cytotoxicity of the OF extracts was initially examined in HepG2 and HeLa cells with the MTT assay, and subsequently in HepG2 cells with the γH2AX In-Cell Western assay. When the MTT assay was used, none of the extracts significantly decreased HepG2 (Fig. 7) and HeLa (Fig. 8) viability at the range of concentrations tested compared to control (P > 0.05 for all tested concentrations of the OF extracts). In a similar way, no significant cytotoxicity was observed in the γH2AX-ICW assay, except for an approximate 50% statistically significant cytotoxicity exerted by the flower (OF1) extract at the highest tested concentration of 2.5 mg/mL (Fig. 10a).

HepG2 cell viability after 24-h treatment with the O. ficus indica extracts (OF1: flower, OF2: cladode, OF3: fruit peel, OF4: fruit seed, and OF5: fruit flesh) as determined by the MTT assay. Results are expressed as the mean of percentage viability ± SEM from at least five independent experiments. Kruskal–Wallis nonparametric test followed by Dunn's post hoc test: No statistical difference between treated and control cells (P > 0.05). MTT, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide.
HeLa cell viability after 24-h treatment with the O. ficus indica extracts (OF1: flower, OF2: cladode, OF3: fruit peel, OF4: fruit seed, and OF5: fruit flesh) as determined by the MTT assay. Results are expressed as the mean of percentage viability ± SEM from at least three independent experiments. Kruskal–Wallis nonparametric test followed by Dunn's post hoc test: No statistical difference between treated and control cells (P > 0.05).
In vitro assessment of genotoxicity
Genotoxicity of OF extracts was evaluated in HepG2 cells by using both the Comet and γH2AX ICW Assay. In the first experimental conditions, none of the OF extracts showed a significant increase in the “% DNA in the comet tail” compared to the negative control after treatment with the previously MTT-defined maximum noncytotoxic concentration of 2.5 mg/mL for 24 h (P > 0.05 for all the extracts tested) (Fig. 9).
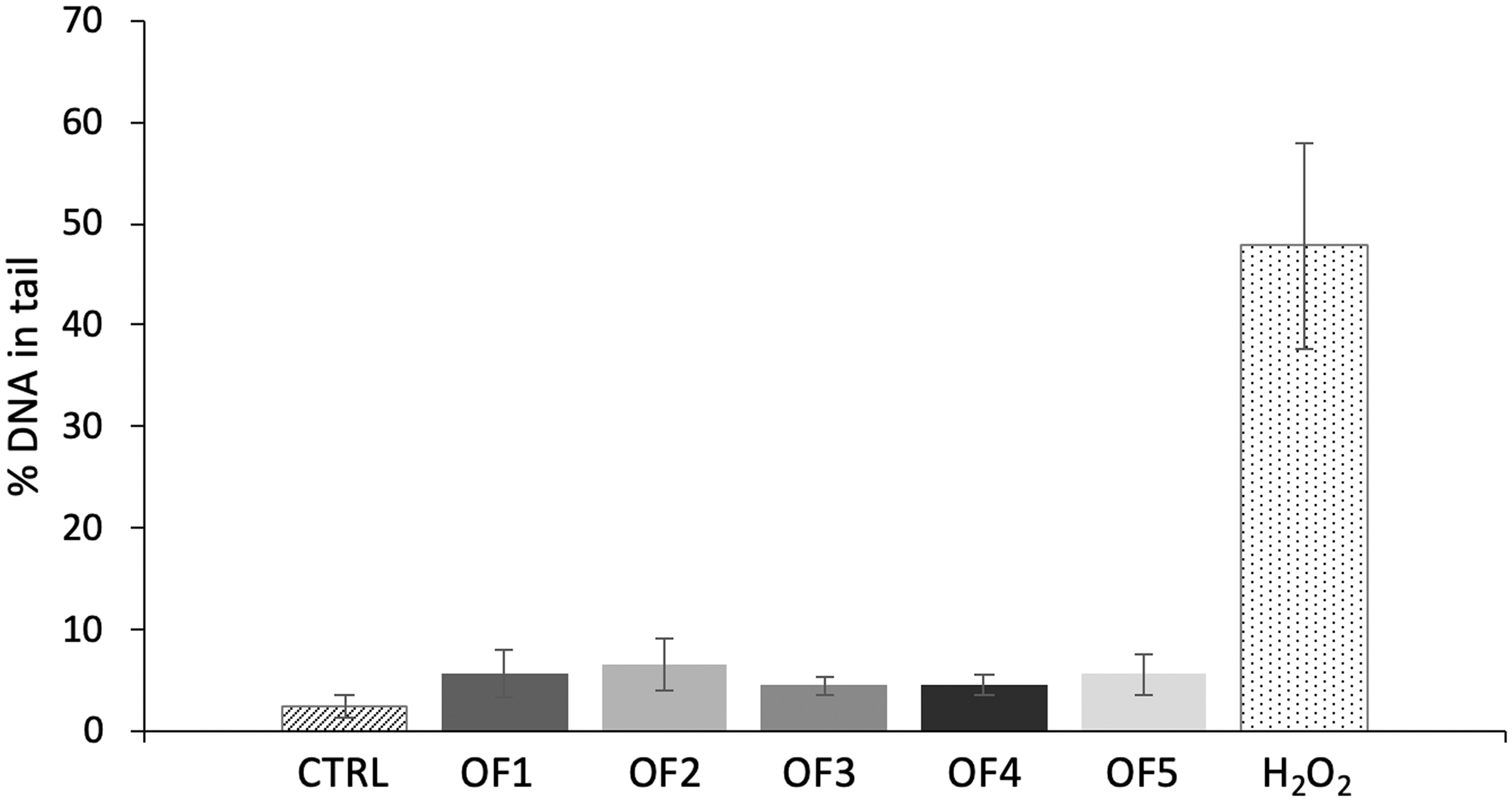
DNA damage in HepG2 cells after 24-h treatment with the O. ficus indica extracts (OF1: flower, OF2: cladode, OF3: fruit peel, OF4: fruit seed, and OF5: fruit flesh) at a concentration of 2.5 mg/mL as determined with the Comet assay. Results are expressed as mean ± SEM “% DNA in the comet tail” from at least three independent experiments. Kruskal–Wallis nonparametric test followed by Dunn's post hoc test: No statistical difference between treated and control cells (P > 0.05).
When γH2AX ICW assay was performed similar results were obtained. Cladode (OF2), fruit peel (OF3), fruit seed (OF4), and fruit flesh (OF5) extracts did not provoke a statistically significant induction of γH2AX compared to the negative control after a 24-h treatment, with concentrations ranging between 0.312 and 2.5 mg/mL, indicating absence of genotoxic potential (P > 0.05) (Fig. 10b–e). However, the flower (OF1) extract caused a statistically significant 2.85-fold induction of γH2AX at the highest concentration of 2.5 mg/mL (P < 0.01), suggesting a DNA damaging activity in this assay (Fig. 10a).

γ-H2AX induction in HepG2 cells and viability after 24-h treatment with the O. ficus indica extracts (

In vitro assessment of the protective effect on H2O2-induced genotoxicity
The protective effect of the OF extracts on H2O2-induced DNA damage in HepG2 cells was assessed by the γH2AX In-Cell Western assay. As shown in Figure 10a, treatment of cells with the flower (OF1) extract at the concentrations of 0.312 and 0.625 mg/mL decreased significantly the induction of γH2AX compared to the positive control (P < 0.01 and P < 0.05, respectively), suggesting a strong protective effect of the extract at low concentrations. At the higher tested concentrations, the increasing cytotoxicity of the extract probably prevented such a protective effect to occur.
Treatment of cells with the cladode (OF2) extract caused a significant decrease in the induction of γH2AX at the highest concentrations (P < 0.01) (Fig. 10b), indicating a protective role at 1.25 and 2.5 mg/mL. As shown in Figure 10d and e, fruit seed (OF4) and fruit flesh (OF5) extracts showed a more pronounced protective effect against H2O2-induced DNA damage, lowering the induction of γH2AX almost at all the concentrations tested between 0.312 and 2.5 mg/mL (P < 0.05 for OF4 extract at the effective concentrations, P either <0.001 or <0.05 for OF5 extract at the effective concentrations). Fruit peel (OF3) extract did not show such a protective effect at the range of concentrations tested (P > 0.05 for all concentrations) (Fig. 10c).
Discussion
In vitro toxicity testing data are essential for screening purposes and an integral part of safety evaluation of chemicals currently. The need of such information for herbal preparations that are of interest for medicinal and dietary purposes is also apparent. In this context, OF extracts were examined for their cytotoxic and genotoxic potential by employing a series of well-established in vitro toxicity tests that examine endpoints of cell death and genotoxicity. Results concerning the toxicity and the antioxidant activity of the extracts are discussed in relevance to their chemical profile.
For the assessment of the potential genotoxicity, HepG2 human hepatoblastoma cells were used. This cell line has been described by the European Centre for the Validation of Alternative Methods (ECVAM) as a suitable in vitro cell model for the detection of genotoxic compounds using different techniques. 36 –43 Furthermore, the HepG2 cells are p53 competent, 44,45 which is known to be a major factor when attempting to reduce the number of “false” positive results in mammalian cell-based genotoxicity assays. 46,47 γH2AX is a well-accepted genotoxic endpoint and its use with HepG2 cells is a suitable test to predict the in vivo genotoxicity of chemicals with different genotoxic MoA. 36,43 Moreover, a good correlation between in vitro micronucleus formation and γH2AX has been demonstrated recently. 48
Cytotoxicity of the OF extracts was first assessed by the MTT assay and subsequently by the γH2AX-ICW assay. The initial testing showed that none of the OF extracts significantly decreased HepG2 and HeLa cell viability after 24 h of treatment at concentrations up to 2.5 mg/mL. Similar results were generated by the γH2AX-ICW in HepG2 cells apart from a significant cytotoxicity observed for the flower (OF1) extract at the highest tested concentration. This difference is related to higher sensibility of γH2AX-ICW assay to determine cytotoxicity. 49
Taking into account the phytochemical profile, the cytotoxicity of the flower extract at the higher concentration could be explained by the pro-oxidant effect of flavonoids. Flavonoid pro-oxidant properties seem to be concentration dependent and directly proportional to the total number of hydroxyl groups. 50 The flower extract resulted particularly rich in quercetin glycosides and especially of its 3′-methoxylated derivative, isorhamnetin-3-O-rutinoside. This compound has been found by several authors as the main flavonoid in flower, cladode, and peel extracts. 51 –53 Quercetin has been repeatedly reported to be cytotoxic in HepG2 cells at concentrations of 50 μM. 53 –56 Kaempferol has been also reported to be cytotoxic. However, cytotoxicity in HepG2 cells was demonstrated at higher concentrations (100 μM). 57 In general, the harmful effect of flavonoids is less severe when they are presented in the glycosylated form. 54,55
Genotoxicity of the OF extracts was examined by using the Comet and the γH2AX ICW Assay. Genotoxicity data followed a very similar trend to cytotoxicity. With the exception of a positive result generated by the flower extract in the γH2AX ICW assay, none of the extracts caused a significant increase in the % DNA comet tail, as well as in the induction of γH2AX. Recently, it was shown that a γH2AX foci assay can be more reliable and sensitive than the Comet assay to detect DNA damage, 58 which can justify the different results obtained by different assays in our study. However, in certain cases, the use of HepG2 cells in the γH2AX-ICW assay fails to detect genotoxicity for compounds that require specific metabolic bioactivation (phase I and II) not sufficiently covered by HepG2 cells. 36,59 –62
The rich content of the flower extract in glycosylated flavonoids compared to the absence or to low levels detected in the remaining extracts could be associated with the genotoxicity observed. Quercetin has been shown to be mutagenic in a variety of widely used in vitro genotoxicity tests, either in bacterial or mammalian/human systems. 36,63 –67 In contrast, isorhamnetin has not been reported as genotoxic, even when tested at three times higher concentration than of quercetin to exhibit a genotoxic effect. 68 However, published data related to the genotoxicity of isorhamnetin are very limited. Kaempferol has also shown mutagenic activity in in vitro genotoxicity tests before, 69 –71 which seems to be enhanced by the transformation of kaempferol to the genotoxic quercetin. 71,72 Although there have been reports that glucosides of quercetin and kaempferol are less mutagenic in contrast to their aglycones in different test systems, 65,69 their genotoxic effect cannot be excluded, given their abundance in the flower extract of our study.
To our knowledge, only the genotoxicity of an OF extract originated from cladodes has been investigated so far. 73 They found no genotoxic potential in the liver of Balb/c mice after the intraperitoneal injection of 100 mg/kg b.w of extract.
In our study, pretreatment of cells with extracts derived from fruit seeds (OF4) and fruit flesh (OF5) showed a pronounced protective effect on the H2O2-induced DNA damage at all concentrations tested. On the contrary, the flower (OF1) and cladode (OF2) extract were effective only at the low (0.312–0.625 mg/mL) and the high (1.25–2.5 mg/mL) concentration, respectively. It is worth mentioning that the flower extract showed the most potent antioxidant activity at low concentrations. It seems that the rich content of flavonoid derivatives in the flower extract may have caused the cytotoxicity observed at the highest concentration. In support of this argument, the extract was effective against H2O2-induced genotoxicity at noncytotoxic concentrations.
As showed by the phytochemical profiling, flavonoid glycosides have not been detected in the seed (OF4) and the fruit flesh (OF5) extracts. These findings were in good accordance with bibliographic data as well. 51,74 The fruit flesh (OF5) extract showed a high content of indicaxanthin, a pigment present in cacti species, while the fruit seed extract (OF4) resulted rich in ferulic acid derivatives. Tesoriere et al. 74 demonstrated that indicaxanthin can defend β-thalassemic RBC from oxidation more effectively than either Trolox or vitamin C, and to repress the expression of intercellular cell adhesion molecule-1 (ICAM-1) in endothelial cells in response to an increase in intracellular oxidants. 75
Fruit seeds have been reported to contain significant amounts of phenolic compounds. 2 In line with our study, feruloyl sucrose isomers are reported as the most abundant feruloyl derivatives. Reports on the function of feruloyl sucrose isomers are rare; however, a research on their antioxidant activity carried out by Nhiem et al. 76 showed similar antioxidant effectiveness to α-tocopherol. The various health benefits might be attributed both to the antioxidant mechanism and to the ability of this molecule to interact with different cellular mechanisms associated with oxidative damage. 77
In summary, for the first time, a comparative assessment has been conducted on five aqueous extracts received from different parts of OF as regard their antioxidant chemical content, cytotoxicity, and genotoxicity, as well as their protective effects against H2O2-induced genotoxicity. Substantial evidences for the lack of cytotoxic and genotoxic effects along specific aqueous extracts of OF strongly support the need for further investigation and exploitation of this plant in the food supplements or functional foods sector.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.