
Abstract
The causative relationship between oxidative stress and aging remains controversial, but it is a fact that many of the pathologies of age-related diseases are associated with oxidative stress. Phytochemicals may reduce damage from oxidative stress; the intake of these through diet could represent a strategy to lessen their pathological consequences. The popular and widely consumed licorice (Glycyrrhiza glabra) is a rich source of potential antioxidants. The aim of this study was to investigate whether licorice increases the oxidative stress resistance and lifespan of the animal model Caenorhabditis elegans. Licorice roots ethanolic extract showed in vitro antioxidant activity, with an IC50 of 51.17 μg/mL using 2,2-diphenyl-1-picrylhydrazyl (DPPH) as free radical. C. elegans pretreated with licorice showed an increase of survival rate when exposed to the oxidant juglone, being this increase up to ∼33.56%. This pretreated population also showed an increase in lifespan of 14.28% at a concentration of 250 μg/mL. In conclusion, we suggest that licorice has a high antioxidant capability both in vitro and in vivo and that this activity may explain the observed extension of lifespan.
Introduction
Oxidative stress has been correlated with numerous pathological states, including cardiovascular diseases, hypertension, diabetes mellitus, cancer and carcinogenesis, neurodegenerative problems, and inflammation. 1,2 These conditions are age-related diseases, and are likely to be exacerbated by the decreased ability to survive stress associated with aging. 3 Antioxidants are usually considered as a possible tool to treat or diminish the consequences of these pathologies. 4 Phytochemicals are especially interesting as they not only lessen the effects of free radicals but also modulate life-prolonging mechanistic pathways 5 and, in contrast, they are frequently found in foods commonly consumed by the population. A clear example is licorice (Glycyrrhiza glabra), a rich source of antioxidant phytochemicals, 6,7 consumption of which comes from the fact that its sweet flavor makes it very popular in the production of candies. 8 This plant has not only been part of diet but also as part of the Chinese traditional medicine 9 and, in fact, licorice has been associated with many pharmacological properties, such as antiallergic, 10 ulcer healing, 11 and anti-inflammatory activities. 12 This bioactivity is thought to be due to the presence of triterpenoid saponins, with glycyrrhizin being the most relevant followed by glycyrrhizic acid. 13
Many of the proprieties of licorice have been studied, but its effects on aging have never been reported. Caenorhabditis elegans represents a useful tool to study aging, not only because of its short and well-studied life cycle but also because of many manipulations that increase stress resistance also increase C. elegans longevity. 14 The C. elegans genome encodes enzymes of the endogenous antioxidant defense systems such as catalase (CAT), glutathione peroxidase (GPx), peroxiredoxins (Prxs), and superoxide dismutase (SOD). 15
The aim of this study was to test whether licorice ethanolic extract improves the ability of C. elegans to face lethal oxidative stress and to study the influence of this plant on the antioxidant defensive system of the worm, by measuring CAT activity. Lifespan assay was also carried out to check if the antioxidant capability of licorice could be correlated to a possible increase on the worm lifespan.
Materials and Methods
Plant material and extraction
Dry licorice roots were provided by Plantarom®. They were crushed to homogeneous particle size. A soxhlet equipment was used to make the extraction. Fifty grams of the crushed licorice was extracted with 500 mL of ethanol for 4 h. Solvent was removed from the extract with a rotary flash evaporator and the extract was then preserved at −20°C to avoid degradation until its use.
High-performance liquid chromatography analysis of glycyrrhizin
The high-performance liquid chromatography (HPLC) analysis was performed with a Waters Alliance e2695 equipped with a 2998 Photodiode Array Detector and a 2414 Refractive Index Detector and chromatographic separations were performed on a C18 2,7 μm (150 × 4,6 mm) column. Gradient elution was used with a mobile phase containing water A (H2O), glacial acetic acid B (CH3COOH), and acetonitrile C (CH3CN) in the following manner: 0–10 min (60% A; 30% B; 10% C), 10–18 min (10% A; 30% B; 60% C), and 18–25 min (60% A; 30% B; 10% C) at a flow rate of 0.8 mL/min. Separations were carried out at 40°C with an injection loop of 20 μg/mL. The analysis was performed at 254 nm. Glycyrrhizin content of the licorice extract was identified by comparing its retention times and UV spectra with those of the standard under identical analysis conditions. Solutions containing different concentrations of glycyrrhizin standards were made to calibrate concentration and peak areas, allowing calculation of the glycyrrhizin content of the licorice extract.
Strains and culture conditions
SS104 and N2 C. elegans strains used in this study were obtained from Caenorhabditis Genetics Center (CGC) USA. SS104 glp-4(bn2) I strain is a temperature-sensitive defect in germ-line proliferation during larval development. This defect can be reversed by shifting worms from restrictive (25°C) to permissive temperature (16°C). SS104 was maintained at 16°C and N2 was maintained at 20°C on Nematode Growth Media (NGM) agar plates with Escherichia coli OP50 as the food source. The medium was prepared using 0.003 g/mL NaCl, 0.005 g/mL peptone, 0.02 g/mL high purity agar, 0.1% cholesterol, 0.1% MgSO4 (1 M), 0.1% CaCl2 (1 M), and 2.5% potassium phosphate buffer (1 M). All chemicals were from Sigma-Aldrich.
Antioxidant activity in vitro: 2,2-diphenyl-1-picrylhydrazyl method
To measure the antioxidant activity, a method based on the reduction of 2,2-diphenyl-1-picrylhydrazyl (DPPH) was carried out. 16 Extracts and DPPH were solved in ethanol. DPPH ethanolic solution was used at 0.04 mg/mL. Absorbance was measured at 517 nm after 30 min of reaction at room temperature in an iEMS Reader Lab systems. Ascorbic acid was used as a positive control. Each concentration was tested in triplicates. Background interferences from solvents were deducted from the activity values of the corresponding extracts before calculating % radical scavenging capacity (% RSC) as follows:
where Abscontrol is the absorbance of the control and Abssample is absorbance of the sample.
The in vitro antioxidant activity of the extract is expressed as IC50, which is defined as the concentration of extract (μg/mL) required to scavenge 50% of DPPH radicals.
Stress-resistance assays
Surco-Laos et al. 17 method was followed with some modifications. Eggs were prepared by bleaching adults, and incubated in M9 buffer until the eggs hatched. L1 larvae were incubated on NGM plates containing E. coli OP50 and different concentrations of licorice, from 0 to 500 μg/mL. Afterward, adult worms were subjected to lethal oxidative stress by means of 5-hydroxy-1,4-naphthalenedione, juglone (150 mM). To assess resistance to oxidative stress, survival was measured after 24 h incubation at 20°C by a touch-provoked movement. Worms that reacted to the mechanical stimulus were scored as alive, whereas nonresponding worms were considered as dead. % Survival rate (% SR) was calculated as a percentage. Assays were performed three times using about 100 individuals per study group.
Measurement of CAT activity
After sublethal oxidative stress induced by juglone (150 mM for 2 h) worms were lysed to extract their proteins. This lysis was made using a cold buffer (150 mM NaCl, 50 mM Tris-HCl pH 8, and 1% TWEEN 20 at 4°C) and breaking worm bodies with ultrasound equipment (JP Selecta 3000683). The concentration of total protein in homogenized worms was measured using BCA kit Thermo Scientific® beforehand and adjusted to the specifications of the CAT activity quantification kit (Cayman Chemical®). Forty nanograms of protein was used per each condition, after the proportional amount of CAT was assessed by taking into account the dilution factor.
Lifespan analysis
Virk et al. 18 method was followed with some modifications. Longevity of temperature-sensitive sterile C. elegans glp-4 (bn2) on four different concentrations of licorice extract was measured in this analysis: 25, 50, 100, and 250 μg/mL and compared with controls.
Gravid adults were used to lay eggs onto fresh NGM OP50 plates at 15°C. Eggs were raised at 15°C till L3/L4 stage, due to temperature sensitivity of mutant phenotypes. Once this stage was reached, animals were transferred to 25°C. After 24 h at that temperature, 25 worms were put onto each of 5 replicate plates for each condition. Animals were transferred to fresh plates after 7 and 14 days and scored for survival every 2–3 days. The scoring method was the same used as for the juglone oxidative stressed assays. Results are expressed as % SR and mean lifespan.
Statistical analysis
Graph Pad Prism® 5.0 was used for statistical analysis. IC50 values were estimated by a nonlinear regression. The statistical differences between the control and treated worms were determined with the aid of the parametric t-test, in the case of the resistance to oxidative stress and toxicity experiments and nonparametric Mann–Whitney test, for the measurement of CAT activity (P value <.05).
Lifespan data were analyzed by JMP statistical software (SAS Institute, Inc., Cary, NC, USA). Relevant statistical significance was determined using the log-rank and Wilcoxon tests of fitting to the Kaplan–Meier survival model. 18
Results
Extract composition
Glycyrrhizin determination on the extract was carried out, as it is the major component of licorice, and one of the main components responsible for its bioactivity. 19 Figure 1 shows that the retention time (min) for the glycyrrhizin present in licorice extract used for this essay and glycyrrhizin standard measured at 254 nm is 9.20 min for both. It can be seen that the presence of glycyrrhizin licorice dry extract was 1.18% ± 0.18% (w/w).
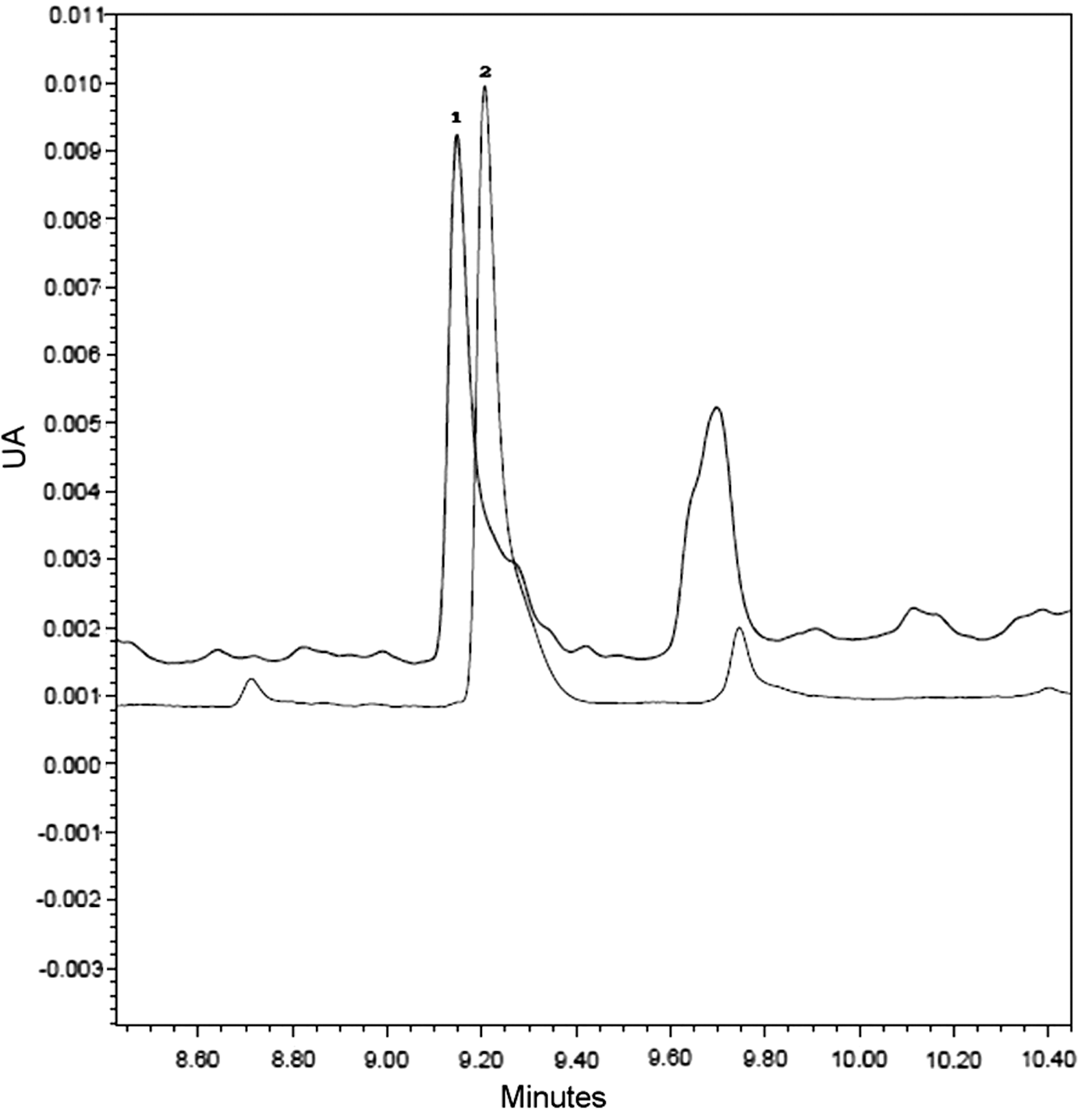
UA versus time (min). HPLC chemical profile of licorice extract. Detection at 254 nm. (1) Glycyrrhizin present in licorice extract. (2) Glycyrrhizin standard at 2.5 ppm. HPLC, high-performance liquid chromatography; UA, units of absorbance.
Antioxidant activity in vitro: DPPH method
DPPH method was carried out as radical scavenging activity to scan the hydrogen-donating capacity of licorice extract. The calculated IC50 value of licorice was 51.17 μg/mL, just one order of magnitude higher than our positive control with ascorbic acid IC50 = 3.20 μg/mL.
Stress-resistance assays
Given the rich content of antioxidant phytochemicals, licorice could be expected to increase the resistance of C. elegans to oxidative stress. The assessment of the antioxidant capability of licorice was carried out by exposing C. elegans to different concentrations of the plant extract with a posterior exposition to an oxidative stress caused by juglone. Figure 2 shows the % SR of pretreated worms at different concentrations of licorice after a 24 h exposition to juglone (150 μM). % SR of the pretreated populations at 50, 100, 250, and 500 μg/mL were 17.78% ± 2.27%, 24.42% ± 2.97%, 24.01% ± 3.21%, and 33.56% ± 2.97%, respectively, being the survival of the control 3.18% ± 0.91%. All of the pretreated population showed significant differences compared with the control (P < .0001).
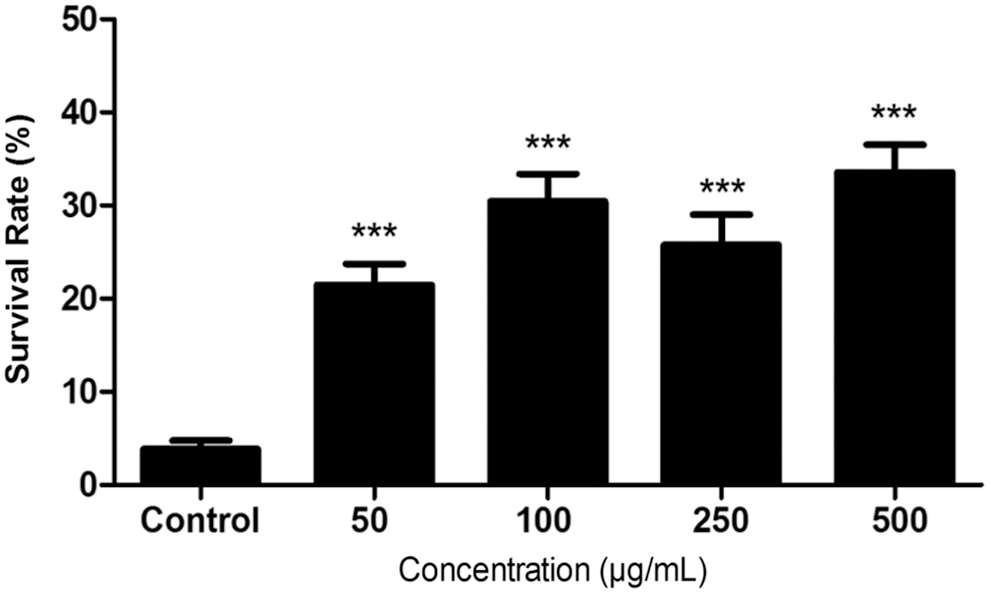
% SR versus licorice concentration (μg/mL). Juglone-induced oxidative stress after pretreatment with 50, 100, 250, and 500 μg/mL of licorice compared with untreated control. Results were obtained after four different experiments, with 350 worms per condition. Data are presented as percentage of survivals (mean ± SEM) ***P < .001. % SR, % survival rate; SEM, standard error of the mean.
Measurement of CAT activity
The modulation of the antioxidant defensive system could have an impact on the protection against oxidative stress. The effect of licorice on one of the enzymes forming part of this defensive system in C. elegans, CAT, was measured. Figure 3a shows the CAT activity in worms treated with 100 μg/mL of licorice or untreated (control). In contrast, Figure 3b shows the CAT activity of the worms treated only with juglone (150 mM, control) and worms previously treated with licorice, 100 μg/mL, before exposed to a sublethal oxidative stress by juglone (150 mM). Population treated with licorice (Fig. 3a) showed an increase of CAT activity of 1600.0% ± 255.9%. The pretreatment with licorice and subsequent exposition to the pro-oxidant juglone (Fig. 3b) produced an increase of CAT activity of 270.5% ± 45.19%.
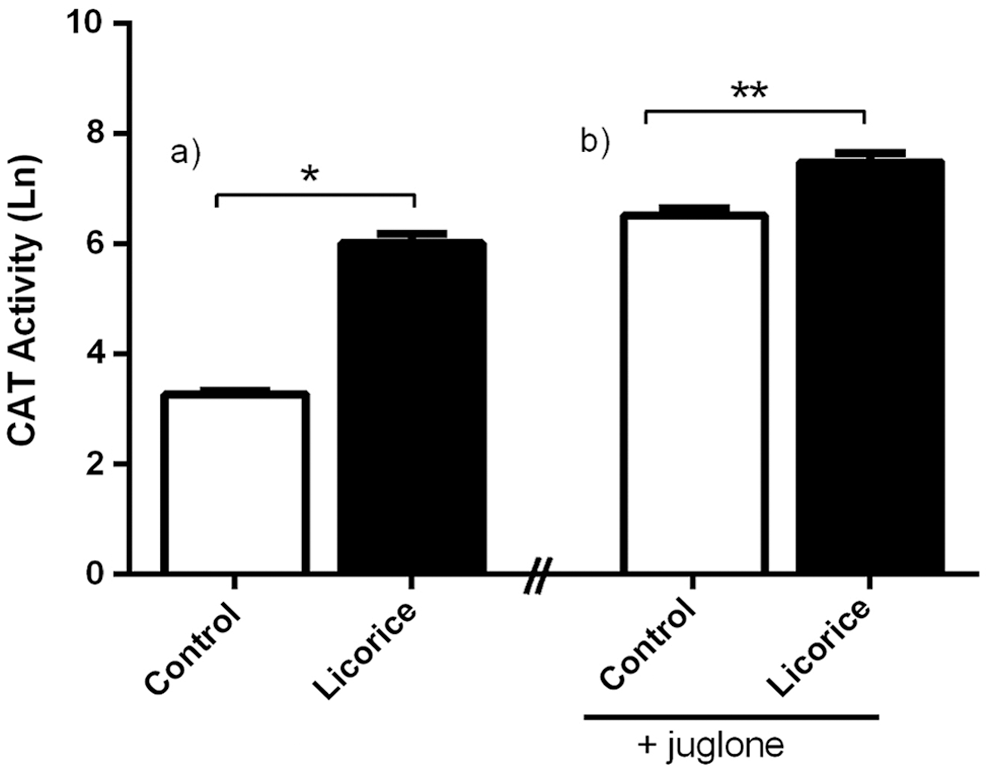
(
Lifespan analysis
Despite the unclear correlation between oxidation and aging, the effect on lifespan of the proven antioxidant extract was tested, to check its possible effects increasing C. elegans lifespan. Figure 4 shows the % SR against time (days) of glp-4 animals (25°C) treated with a concentration of licorice (25, 50, 100, and 250 μg/mL) compared with control. Mean lifespan was 12.84 ± 0.20, 12.92 ± 0.20, 12.58 ± 0.19, and 13.59 ± 0.19 days for 25, 50, 100, and 250 μg/mL licorice concentrations, respectively, being the control survival 11.87 ± 0.17 days. This effect is stronger at 250 μg/mL, with 14.28% extension of mean lifespan (P < .0001). There were significant differences between 250 μg/mL and the other concentrations (P < .05), but there were not among 25, 50, and 100 μg/mL.
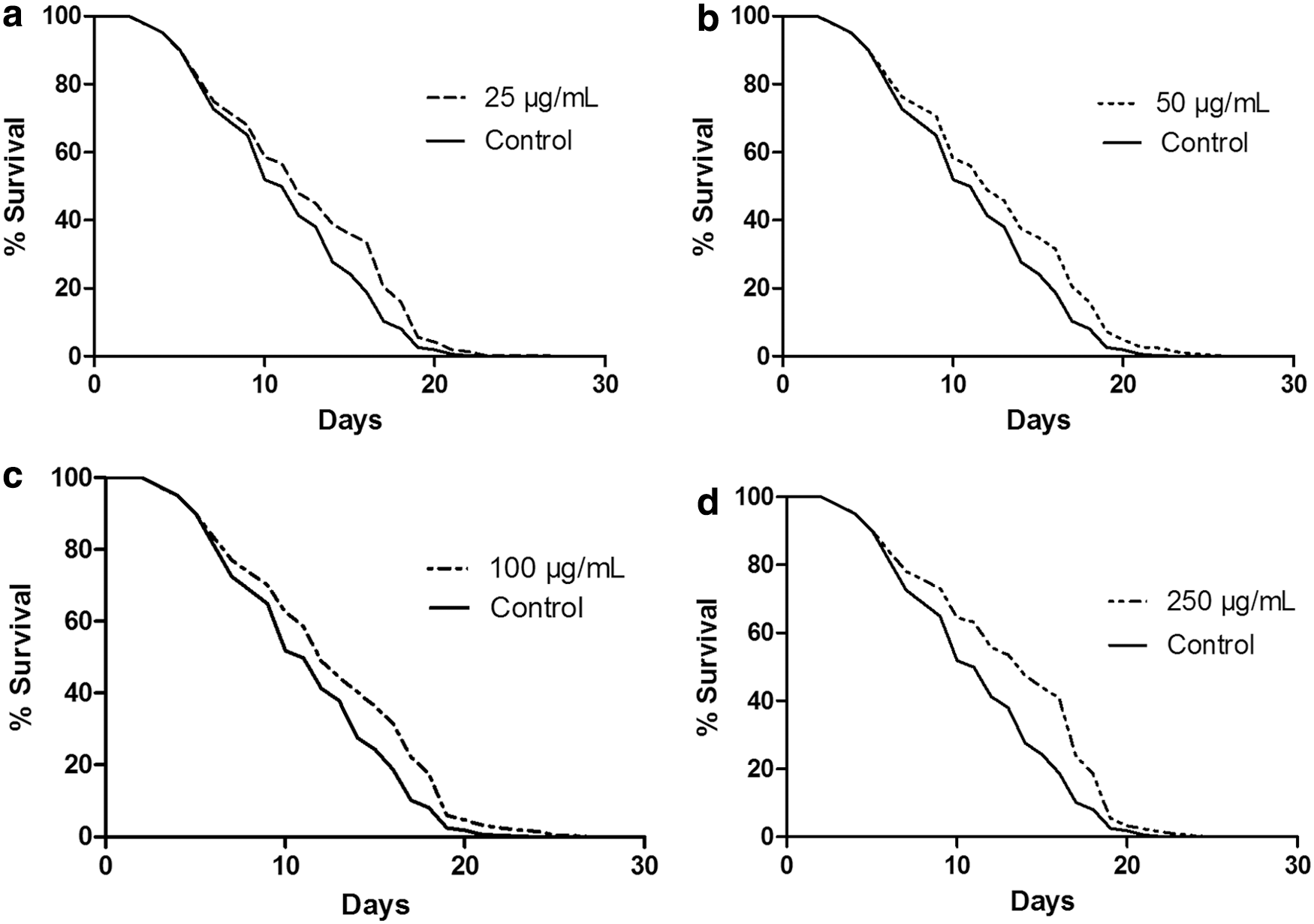
% SR against time (days) of SS104 glp-4 animals (25°C) treated with a concentration of licorice of
Discussion
The chemical composition of the licorice extract mainly includes triterpenoid saponins (glycyrrhizin and glycyrrhizic acid), flavonoids, chalcones, and other compounds present in lower quantities. 20 Glycyrrhizin, apart from being present in higher quantity, also presents significant bioactivity. 19 Licorice capabilities, discussed later, are probably given by the cited content of glycyrrhizin and polyphenols, and their synergic action with other untested components.
As cited before, the IC50 value of licorice obtained with the DPPH method was one order of magnitude higher than the ascorbic acid, but, taking into account that licorice extract is a mixture of chemical compounds and vitamin C is a pure substance, licorice ethanolic extract could be considered as a source of antioxidant compounds. This capability is biologically important as the ability to scavenge free radicals of the DPPH type may imply the utility of licorice in preventing the damage of cellular membrane, the oxidation and cross-link of proteins and harmful interactions with DNA, both nuclear and mitochondrial. 21
Owing to the complexity of licorice composition, its antioxidant activity could be given by several simultaneous mechanisms of action. Thus, the animal model C. elegans was used to test the antioxidant in vivo capability, and to test the influence of licorice extract on the endogenous antioxidant defensive system of the worm. This oxidant stress was generated by juglone. Its toxicity is caused by several mechanisms of action, including the ability to undergo oxidation–reduction cycles with concomitant formation of free radicals. 22 The pretreatment of C. elegans with licorice has shown an increase of the survival at all tested concentrations (Fig. 2). According to our results, the protection is not given in a dose-dependent manner. The increase in SRs indicates that licorice is an efficient antioxidant in vivo.
As far as we know, there are no previous studies of the licorice antioxidant activity on C. elegans, but similar results were obtained with green tea, a well-known antioxidant, where the average of survival for a population pretreated with 100 μg/mL of green tea extract was 32.43% ± 1.71% after 24 h exposure to juglone (80 μM). 23 In this experiment with green tea, the juglone concentration was lower, leading to a higher survival of both the control and the treated population. In any case, the increase of survival was in the same order of magnitude of the one obtained with licorice at the same dose.
Flavonoids are one of the most important phytochemical contributors to the antioxidant capability of plants. Epigallocatechin gallate, from green tea, or quercetin, a major flavonoid in human diet, has shown its efficacy in increasing C. elegans resistance to stress. 17,24 Also, terpenes such as specioside (6-O-coumaroylcatalpol) isolated from the plant Stereospermum suaveolens have shown this protection against oxidative stress. 25 According to the literature, the rich content of flavonoids and terpenes of licorice 20 could be responsible for its capability to increase the survival of C. elegans in response to oxidative stress.
This improvement of the survival against the pro-oxidant juglone might not be given only by the RSC of licorice. Formation of free radicals is part of many physiological processes 1 that, in an ideal situation, are balanced by an elaborated endogenous antioxidant defensive system, composed of enzymes such as SOD, CAT, or GPx, among others. 26
As it can be seen in Figure 3, CAT activity was enhanced on licorice-treated worms in comparison with untreated control group. There was an increase of CAT activity on the untreated population when exposed to oxidative stress. This could be expected, as CAT forms part of the endogenous enzymatic system of protection of C. elegans. 27 In contrast, licorice treatment increases CAT activity in C. elegans both in response to oxidative stress induced by external stressor, as well as in absence of it. This fact suggests that licorice consumption could improve CAT activity as part of the natural molecular mechanisms of cellular detoxification against free radicals.
Some authors establish a correlation between the enhanced stress resistance and longevity. 28,29 A lifespan essay was carried out with licorice, as its probed antioxidant capability suggested its capability to increase C. elegans lifespan. According to our results, licorice treatment increases the life of C. elegans in a dose-dependent manner, as even all concentrations extended lifespan, the highest concentration showed the strongest effect.
Despite the fact that there is no background literature on the effects of licorice on C. elegans lifespan, there are previous studies about the activity of other phytochemicals. Some examples are polyphenols from blueberry, 30 epigallocatechin gallate from green tea 24 and terpenes, such as specioside, from patala (S. suaveolens). 31 Both, terpene and epigallocatechin gallate, also showed an increase in C. elegans tolerance to oxidative stress induced by juglone, which, in concordance with the results obtained for licorice, agrees with previous studies that show that longevity is closely linked to oxidative stress resistance. 24,32 The way licorice prolongs lifespan by diminishing oxidant damage may be given by its radical scavenging capability itself and the enhancement of the activity of C. elegans endogenous enzymatic system.
In conclusion, according to the exposed results, we conclude that licorice has a strong antioxidant capability both in vitro and in vivo. This antioxidant capability may be given by the radical scavenging capability of licorice and the enhancement of CAT activity provoked by licorice extract, both in basal conditions or when exposed to an oxidative stress. It has been also shown that licorice extends C. elegans lifespan. This lifespan extension may be correlated with the previously cited antioxidant capability.
Even though the in vivo model used in this essay is still far from the human being, it shows how diet has a huge influence not only on preventing pathologies but also on aging.
Footnotes
Acknowledgments
We thank Dr. Mainar and Carlota Giménez for their collaboration on HPLC analysis. We also thank Claire Maynard for the training and carryout of lifespan assays.
Funding Information
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author Disclosure Statement
No competing financial interests exist.