
Abstract
Metabolic syndrome is recognized as a proinflammatory condition leading to hepatic steatosis and nonalcoholic steatohepatitis (NASH). We tested the effects of a succulent species Hoodia parviflora N.E. Br., of the genus Hoodia sweetex Dence, on animal models of NASH and insulin resistance (ob/ob mouse and the sand rat Psammomys obesus). IL6 secretion was evaluated by ELISA and hepatic signal transducer and activator of transcription 3 by Western blot. We followed liver enzymes, weight, glucose, hepatic histology, hepatic triglycerides (TGs), and total fat and serum insulin. Oral administration of extracts derived from H. parviflora alleviated the insulin resistance manifested by improved glucose tolerance tests. Treatment alleviated the liver injury noted by a decrease in liver enzyme levels, improved intrahepatic TG content, total hepatic fat, and improved hepatic histology. Similarly, treatment with H. parviflora reduced hepatic inflammation in mice with Concanavalin A-induced hepatitis. These effects were independent of food consumption and weight. H. parviflora was associated with alleviated insulin resistance, hepatic steatosis, and liver injury. The data support its use as a liver protector.
Introduction
Nonalcoholic fatty liver disease (NAFLD) is a common form of chronic liver disease in the Western world and is estimated to affect >30% of the U.S. population. 1 NAFLD is considered a hepatic manifestation of the metabolic syndrome, and its prevalence is expected to increase as the obesity and diabetes epidemics evolve. 2 The first manifestation of injury is the accumulation of fat within hepatocytes that in some patients is followed by the development of necroinflammatory activity and nonalcoholic steatohepatitis (NASH), leading to cirrhosis. Liver cell ballooning and the development of inflammation determine whether a patient progresses to irreversible liver damage and fibrosis. 3 –6
Clinical and epidemiological evidence suggest that NASH, being the hepatic manifestation of metabolic syndrome, is associated with insulin resistance. 5 Insulin resistance, oxidative stress, mitochondrial dysfunction, immune dysregulation, and adipokines seem important to the pathogenesis of NAFLD. 3 Cytokines, and particularly chemokines, are important in driving the inflammatory infiltrate in NASH. 7 The role of visceral adipose tissue and adipokines in the exacerbation of both insulin resistance and NASH has been described. 8,9 As the immune system plays a role in the pathogenesis of NASH and insulin resistance, immunomodulatory agents are being developed as potential therapies for NASH. 10 –18
Hoodia is a genus of succulent plants, originally from the arid regions of South Africa and Namibia. 19 Hoodia plants are multistemmed succulents with thick, erect, cylindrical, glabrous, fleshy, and fairly hard stems that are gray–green to gray–brown. They bear large cup-shaped corollas that can reach up to 100 mm in diameter. 19 The genus Hoodia is classified as one of the Stapeliads, a group of stem succulents belonging to the family Apocynaceae. The Hoodia genus includes at least 13 reported species: H. alstonii, H. currorii, H. dregei, H. flava, H. gordonii, H. juttae, H. mossamedensis, H. officinalis, H. parviflora (HP), H. pedicellata, H. pilifera, H. ruschii, and H. triebner. 20 Historically, the San people of the Kalahari Desert would eat fresh Hoodia plants during their hunting trips to suppress hunger. Within the past decade, Hoodia has grown from an almost forgotten spiny desert plant to an important commercial product, a food and dietary supplement, and an herbal medicine used as an appetite suppressant. 21,22
Oral administration of Hoodia plants is common in many developed countries for its reported suppressive effect on appetite. 23 Studies have identified extracts from various Hoodia species, including H. gordonii and H. pilifera that possess appetite-suppressing properties. 19 The appetite-suppressing properties have been attributed to a family of Hoodia steroid glycosides identified in dried plant material, purified and enriched extracts, and in various prototype food products fortified with H. gordonii extracts. 24,25 Two pregnane glycosides were isolated by fractionating dried stems from H. gordonii or H. pilifera. Oral administration of these compounds to rats resulted in decreased food consumption over an eight-day period and a decrease in body mass compared with vehicle. 19 In a comparison of the chemical fingerprint of four Hoodia species, including H. gordonii, H. currorii, H. ruschii, and H. parviflora, a chemical diversion among the different Hoodia species was identified. For instance, a novel compound was found that was more prominent in HP than in other species of Hoodia. 26 It is assumed that the different chemical profiles of the various Hoodia species may result in distinct effects on the consumer.
The anti-inflammatory effects of Hoodia extracts were not studied previously. In this study, we focused on the effect of HP on the Concanavalin A (Con A) model of immune-mediated hepatitis, and in two models of NASH: the leptin-deficient ob/ob mouse 15,27,28 and the sand rat (Psammomys obesus). 13 The aim of this study was to determine the mechanism of HP-mediated anti-inflammatory effects and to assess its role in insulin resistance and NASH.
Materials and Methods
Preparation of HP extracts
Two HP extracts were tested in this study: P2 and pure sap. To prepare P2, HP daughter plants cultivated from seed from certified H. parviflora mother plants were harvested. Mother plants were maintained in isolated greenhouses (temperature range of 25–30°C and relative humidity range of 30–35%) at Kibbutz Yotvata (Southern Arava Valley, Israel), an area characterized by its desert climate. Immediately after harvest, the fresh aerial parts were washed and frozen at −18°C. Within 24 h, the frozen plants were cut into 5-cm sections and resuspended in 5–12% (v/v) mineral water. The suspended HP plant tissue was then sonicated for 10 min in a 43-kHz ultrasonic bath (MRC, model D-80H) for further extraction. Aliquots of the suspended HP plant tissues (200 mL each) were homogenized for 2 min using a homogenizer (IKA WERKE, model T 25 B). The suspended HP plant tissues were then separated from the aqueous medium by filtration using a 20-μm filter and centrifugation at 10,000 rpm for 5 min to obtain clear liquid HP extract. All steps were carried out at temperatures <30°C. To prepare pure sap, HP daughter plants, as already described, were cut crosswise with a sharp knife, and a thick liquid was collected directly from the plant stem using an injector. The sap was then centrifuged at 10,000 rpm for 5 min to obtain clear resin. All steps were carried out at temperatures below ∼30°C.
Animals
Several animal models were investigated in this study. For the Con A model, we used 11–12-week-old male C57BL/6 (B6) mice that were purchased from Harlan Laboratories (Jerusalem, Israel). For the NASH model, we used 6–7-week-old male leptin-deficient ob/ob mice purchased from Harlan Laboratories. All mice were maintained in the Animal Core of the Hadassah-Hebrew University Medical School. Mice were given standard laboratory chow and water ad libitum and kept in a 12-h light/dark cycle. Mice were treated orally once 2 h before Con A injection.
For the nutrition-induced type 2 diabetes mellitus (T2DM) model, we used the sand rat, P. obesus. Five-month-old male P. obesus rats were purchased from Harlan laboratories (Jerusalem, Israel) for these studies. Rats were housed in solid-bottomed polypropylene cages equipped with water bottles and Aspen woodchip bedding and were fed a high-energy (HE; 2.93 kcal/g) artificial diet. All animal experiments were carried out in accordance with the guidelines of the Hebrew University-Hadassah Institutional Committee for Care and Use of Laboratory Animals and with the committee's approval. Animals were treated orally five days per week.
Administration of Con A
Con A was purchased from MP Biomedicals (Ohio, USA). Con A was dissolved in 200 μL of 50 mM Tris (pH 7), 150 mM NaCl, and 4 mM CaCl2 and injected (20 mg/kg) to mice intravenously 20 h before sacrifice.
Hepatocellular damage
The effect of HP extracts on liver injury was determined by assaying serum aspartyl transaminase (AST) and alanine aminotransferase (ALT) activities, which were measured by the Reflovet Plus system (Roche Diagnostics, Mannheim, Germany).
Cytokine determination
Serum IFN-γ and IL6 levels were determined by “sandwich” ELISA using commercial kits (Quantikine, R&D Systems, Minneapolis, MN, USA), according to the manufacturer's instructions.
Immunoblot analysis
For analyzing whole tissue lysates, snap-frozen liver sections from P. obesus were homogenized in 0.75 mL of ice-cold phosphate-buffered saline and then centrifuged at 4°C for 5 min at 2000 rpm. Supernatants were discarded, and pellets were lysed in 50 mM β-glycerophosphate, 1.5 mM EGTA (pH 7.3), 1 mM EDTA (pH 7.3), 1 mM DTT, 0.1 mM sodium vanadate, 1% NP-40, and cocktails of proteases and phosphatases (Sigma, Rehovot, Israel) during a 10-min incubation on ice. Lysates were centrifuged at 4°C for 10 min at 14,000 rpm, and the supernatants were transferred to clean tubes. Volumes of lysates containing equivalent amounts of protein (15 μg per lane), as determined by the Bradford assay (Bio-Rad, Rehovot, Israel), were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membranes. Proteins were detected with signal transducer and activator of transcription 3 (STAT3) and phospho-STAT3-Ser727 antibodies (Cell Signaling Technologies, MA, USA); antibodies directed against β-actin (Abcam, Cambridge, UK) were used as a loading control. All blots were developed with horseradish peroxidase-conjugated secondary antibodies (Jackson Laboratories Bar Harbor, ME, USA) and Western blot-Luminol Reagent (Santa Cruz Biotechnology, CA, USA).
Hepatic fat content measured by magnetic resonance imaging
P. obesus rats underwent magnetic resonance imaging (MRI) on day 21. Hepatic fat content was measured with a double-echo chemical shift gradient-echo sequence technique. This technique provides in-phase and out-of-phase images in a single acquisition for fat assessment and quantification. T1-weighted out-of-phase MRI is sensitive for the detection of relatively small proportions of tissue fat. Magnetic resonance images were acquired with a 1.5-T system (Sigma LX; General Electric, Milwaukee, WI, USA). Double-echo MRI was performed with a repetition time of 125 msec, double echo times of 4 and 6.5 msec, and a flip angle of 80°. Imaging parameters included a section thickness of 3 mm, a 13-cm field of view, and a 256 × 160 matrix. Axial and coronal images were obtained. Signal intensity (SI) changes between in-phase and out-of-phase images were computed. The SI index was calculated as follows: SI index = (SIip − SIop)/SIip, where SIip is in-phase SI and SIop is out-of-phase SI. Low SI index values indicate a smaller amount of tissue fat.
Body fat distribution measured by MRI
To determine the effect of treatment on body fat distribution, cervical and intraperitoneal fat contents were evaluated as measures of peripheral and central fat.
Insulin determination
Serum insulin levels were determined using a commercially available ELISA kit (Mercodia AB; Uppsala, Sweden), according to the manufacturer's instructions. Serum was collected from euthanized ob/ob mice or from P. obesus rats on the day of sacrifice.
Glucose tolerance test
Mice or P. obesus rats in all groups underwent an oral glucose tolerance test (OGTT) after overnight fasting on day 30. Glucose was administered orally (1.25 g/kg). Serum glucose measurements were performed on tail-vein blood every 15 min for 3 h and measured using a standard glucometer.
Assessment of lipid accumulation in the liver
The accumulation of hepatic triglycerides (TGs) within the liver was quantified using a modification of the Folch method. TGs were extracted from aliquots of snap-frozen livers and assayed spectrophotometrically using the GPO-Trinder kit (Sigma, Rehovot, Israel). Heaptic TGs levels were normalized to the protein contents in the homogenate. For Oil Red O staining, livers were taken immediately after sacrifice and cut into small square sections and mounted on stubs with tissue-Tek (Reichert Jung, Buffalo, NY, USA) and rapidly frozen in liquid nitrogen. Cut sections (6-μm thick) were mounted on gelatin-coated slides for Oil Red O staining.
Measurement of serum lipids
Serum TG and total cholesterol levels were determined using a Reflovet Plus clinical chemistry analyzer (Roche Diagnostics).
Statistical analysis
The comparison of two independent groups was performed using the Student's t-test and nonparametric Mann–Whitney test. The association between two variables was assessed by calculating the Pearson and the Spearman correlation coefficients. All tests applied were two-tailed, and a P-value of .05 or less was considered statistically significant.
Results
Oral administration of HP alleviates liver injury in Con A immune-mediated hepatitis
The Con A immune-mediated hepatitis model was used to determine the effect of HP extract on liver inflammation. We tested two volumes of P2 HP extract (11 and 33 μL) and compared it with pure sap derived from the same plant. As shown in Figure 1A, both P2-treated and pure sap-treated mice experienced a decrease in serum levels of ALT and AST in response to Con A injection relative to control mice, which were injected with vehicle alone.
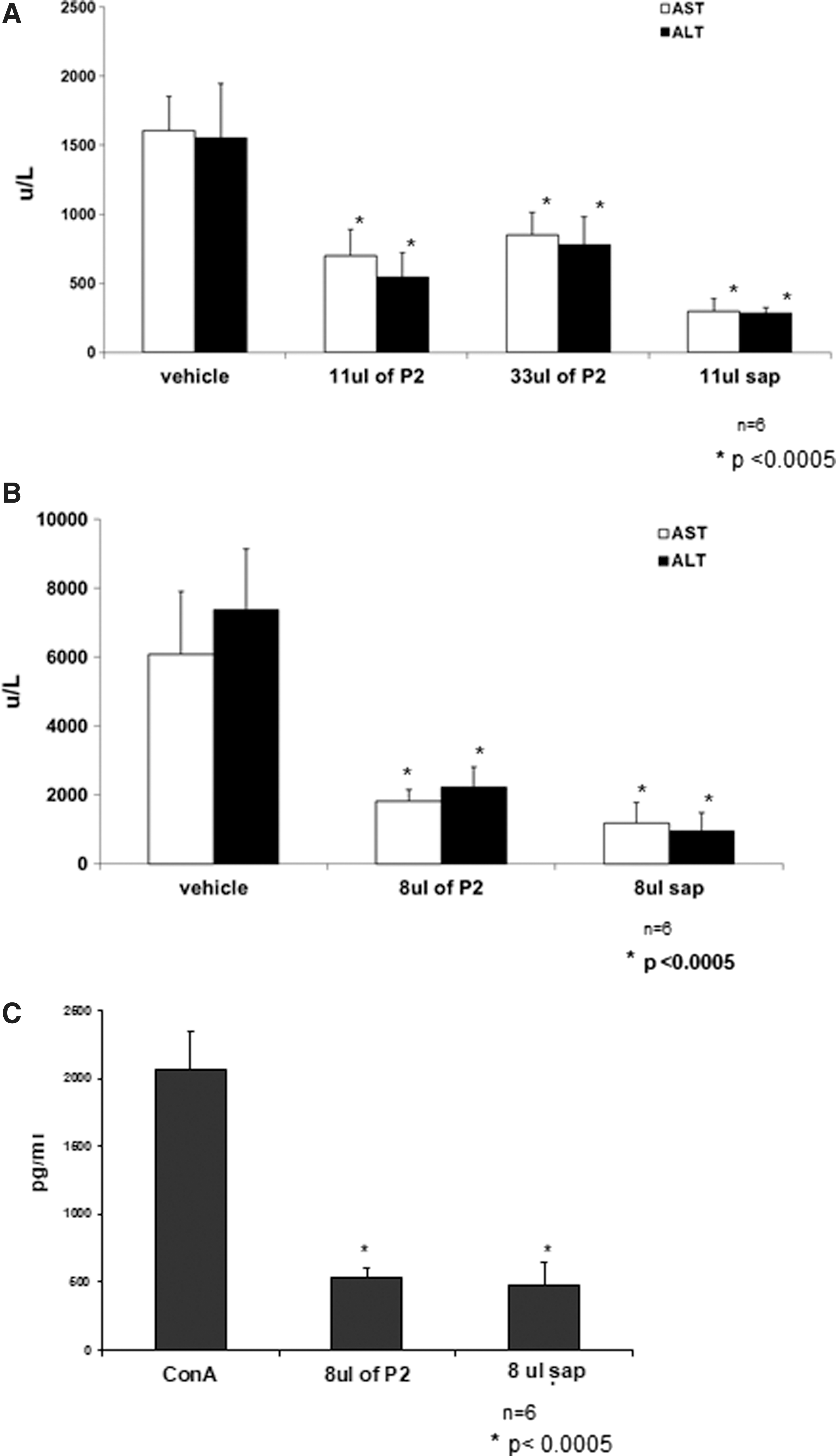
The effect of HP extracts on liver enzymes in Con A-induced hepatitis.
Mice treated with lower doses of P2 extract (8 μL) exhibited a similar beneficial decrease in liver enzymes (Fig. 1B). Serum ALT levels were more than threefold lower in P2-treated mice, whereas ALT levels in sap-treated mice were decreased by more than sevenfold.
Con A-induced hepatitis is associated with the release of high amounts of IFN-γ. Figure 1C shows that the anti-inflammatory effect of P2 extract and its sap-derived counterpart were associated with significant decreases in serum IFN-γ levels. Serum IFN-γ levels in P2-treated mice were fourfold lower than in vehicle-treated control mice. Taken together, these data imply that P2 extracts possess anti-inflammatory properties.
Effect of oral administration of HP on activation of P-STAT3 and serum IL6 secretion in P. obesus rats
The sand rat is a model of nutrition-induced T2DM that develops significant hyperinsulinemia, hyperglycemia, and hypertriglyceridemia when placed on a HE diet. The IL6-STAT3 pathway was suggested to play a role in the pathogenesis of NASH. 29 To determine the potential mechanism by which HP extracts affect the metabolic syndrome, we examined this pathway in the livers of P. obesus rats. Figure 2A shows that oral administration of P2 extract showed a trend for increased P-STAT3 activation (P = NS). By contrast, P-STAT3 activation was not observed in animals given HP sap or GT2 Hoodia extracts (HP grown in Gordonii tunnels). Serum IL6 levels were elevated in HP-treated animals as shown in Figure 2B.
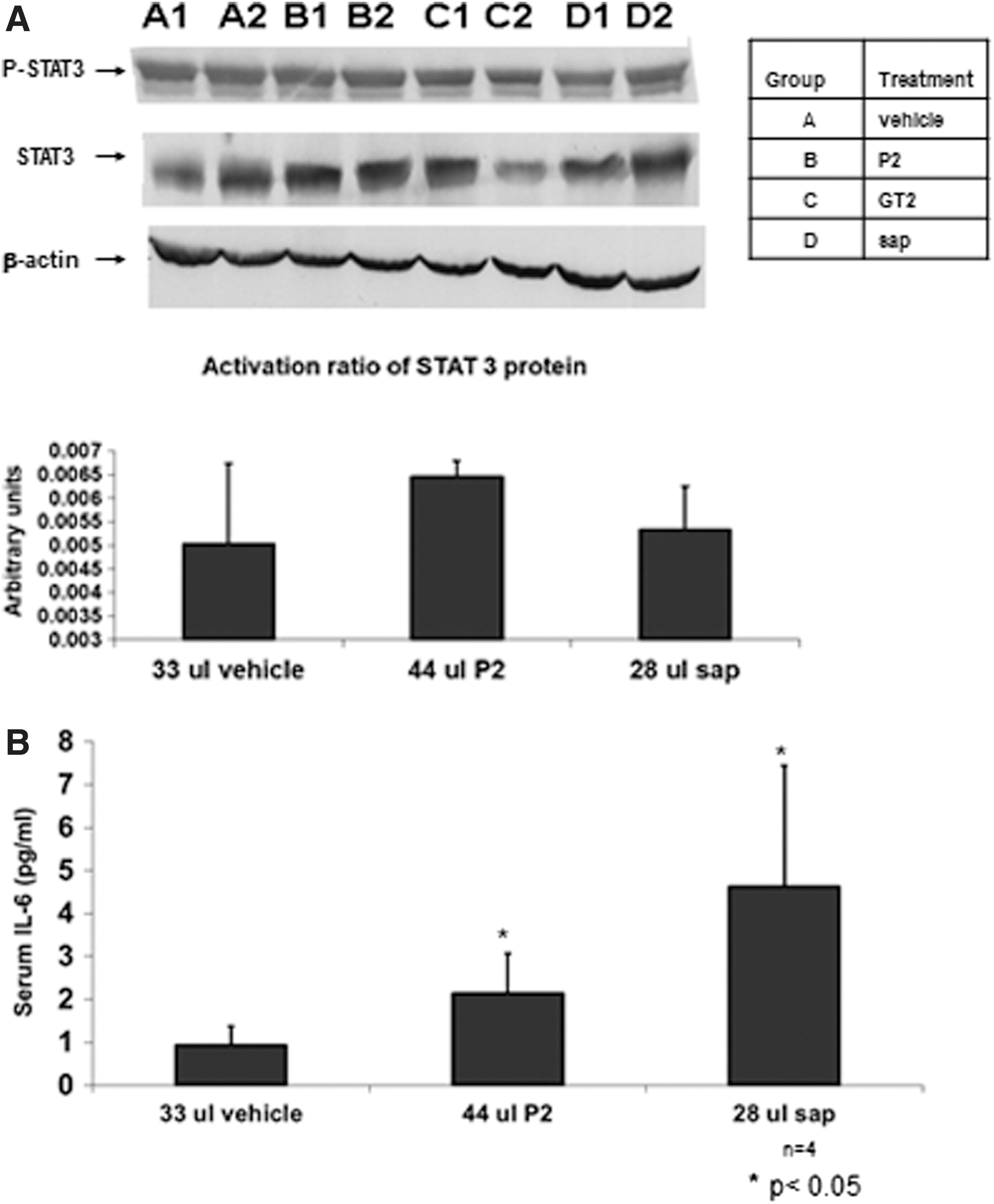
Effect of P2 and pure sap extracts on the IL6-STAT3 signaling pathway of P. obesus.
Oral administration of HP alleviates metabolic syndrome in P. obesus rats
Figure 3A shows a decrease in serum ALT and AST levels on days 7, 14, and 28 of treatment (P = NS). On day 28, ALT and AST serum levels were increased in control mice, which contrasted with results from P2- and pure sap-treated mice.
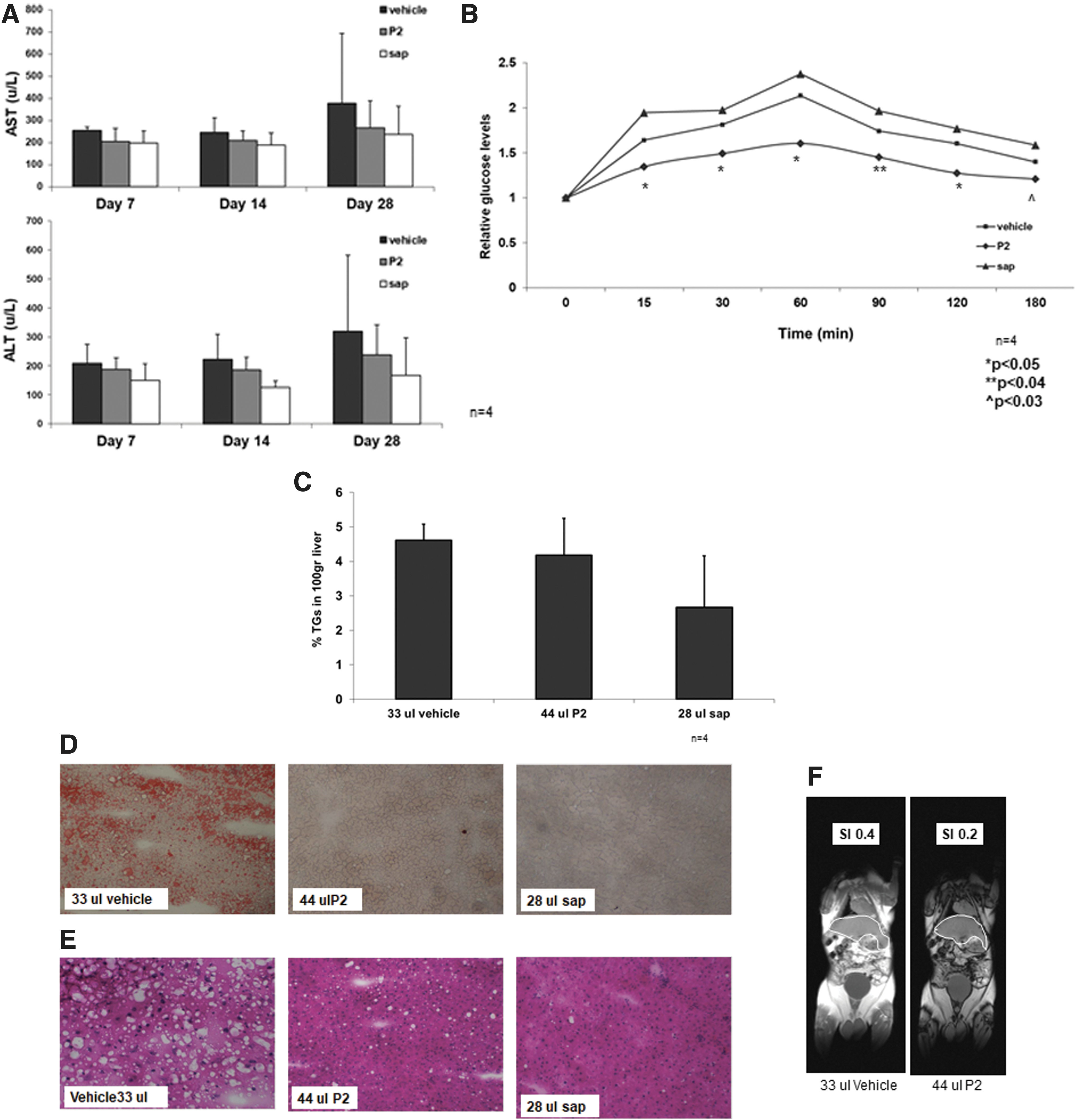
The effects of P2 and pure sap extracts in P. obesus rats on liver enzymes.
Figure 3B shows the beneficial effect of Hoodia extracts on insulin resistance. The OGTT was performed on P. obesus rats that received one of the extracts of Hoodia orally. For each time point, an improvement in glucose levels was observed in rats treated with any of the two extracts. The most profound effect was noted for rats treated with P2: at 15 min, the levels in treated animals were 1.34 and 1.72 versus 1.83 mg% for P2- and sap-treated animals versus controls (P < .05); at 30 min, P2- and sap-treated animals harbored blood glucose levels of 1.49 and 1.94, respectively, versus 1.93 mg% in untreated controls (P < .05); at 60 min, blood glucose levels were 1.6 and 2.09, versus 2.13 mg% (P < .05); at 90 min, levels were 1.45 and 1.75 versus 1.86 mg% (P < .04); at 120 min, levels were 1.27 and 1.56 versus 1.60 mg% (P < .05); and at 180 min, blood glucose levels were 1.211 and 1.09 versus 1.40 mg% (P < .03).
Figure 3C shows the beneficial effect of pure sap extract on liver steatosis. A trend (P < .06) of decrease in liver TG content was noted for sap-treated animals. We further examined hepatic fat accumulation by immunohistochemistry. Figure 3D shows Oil Red O staining of livers from treated P. obesus rats, demonstrating a clear reduction in hepatic fat content. This effect was also noted in H&E liver staining (Fig. 3E), in which a lower fat content was observed in P2- and sap-treated animals compared with control-treated animals. To further assess the quantitative decrease in liver steatosis in treated animals, a whole-body MR was performed for a quantitative assessment of the amount of fat per liver on two animals per group. Figure 3F shows representative MR sections, indicating a decrease in the amount of fat accumulation in the livers of treated animals.
Oral administration of HP alleviates metabolic syndrome in ob/ob mice
To further evaluate the effect of HP extracts in a NASH model, we explored the effect of these extracts on the leptin-deficient ob/ob mouse model. Figure 4A shows that neither P2 nor pure sap had a significant effect on weight gain. Figure 4B shows the effect of HP extracts on serum insulin levels. A small trend for an increase in serum insulin levels was noted in P2- and sap-treated animals after fasting (P = NS). Further evaluation of the effect of HP extracts on insulin resistance was carried out by OGTTs. Figure 4C shows a trend for an improved OGTT in P2-treated animals for each time point examined (P = NS). However, glucose tolerance was not improved in pure sap-treated ob/ob mice.
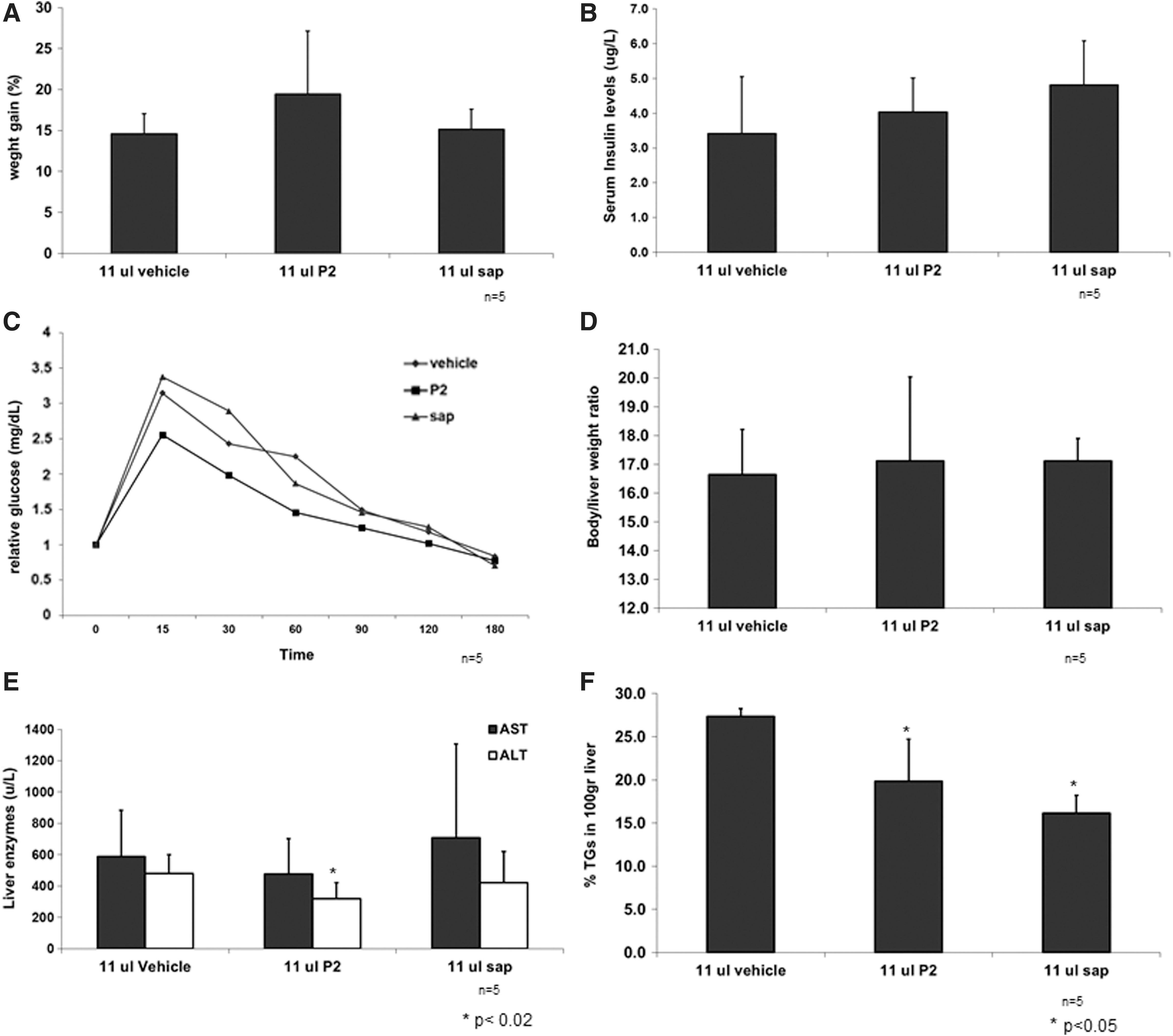
The effect of P2 and pure sap extracts in leptin-deficient mice.
Figure 4D shows that HP extracts did not lead to changes in the body-to-liver weight ratio. Figure 4E shows the effect on liver injury in leptin-deficient mice. On day 30 post-treatment, a significant decrease in ALT levels was observed in P2-treated mice compared with control mice. Figure 4F shows a beneficial effect of P2 and sap extracts on the liver TG content. A significant decrease in fat content was also noted in P2- and sap-treated ob/ob mice.
Discussion
In this study, our data show that oral administration of HP extracts, P2 and pure sap, exerts an anti-inflammatory effect that improves insulin resistance, alleviating hepatic steatosis and associated liver injury.
Metabolic syndrome is recognized as a proinflammatory condition. 30,31 Chronic inflammation also contributes to the pathology of T2DM. 32 –35 In the process of inflammation, macrophages and endothelial cells contribute to increased serum levels of cytokines, including IL1β, IL6, and TNF-α in these patients. 36 Dysregulated inflammatory processes, manifested by visceral inflammation in the liver and adipose tissue, result in insulin resistance. 37
In the Con A immune-mediated hepatitis models, a significant decrease in ALT and AST levels was observed in HP-treated mice. The HP extracts exhibited a dose-dependent effect, with improved symptoms observed in mice treated with low doses. The anti-inflammatory effect was reflected by a significant decrease in serum IFN-γ levels.
P. obesus is a member of the gerbil subfamily and serves as a model of nutritionally induced T2DM, characterized by primary insulin resistance. 38 –41 P. obesus is adapted to life on a low-energy (LE) diet, characterized by a thrifty metabolism. When transferred to a HE diet, they develop obesity, hyperinsulinemia, hyperglycemia, and hypertriglyceridemia. 13,39 Increased hepatic TG production and apolipoprotein B induction have been demonstrated in hyperinsulinemic P. obesus rats, 42 which leads to hepatic steatosis. 43 The ob/ob leptin-deficient mouse develops insulin resistance that may be mediated by immune defects in TNF production or NKT cell function. 28,27 Insulin resistance was alleviated in HP-treated animals, as reflected by improved OGTT in P. obesus rats. In P2-treated ob/ob mice, a trend for improved OGTT was noted at each of the time points tested. The improved insulin resistance was associated with an effect on liver steatosis in P. obesus rats and ob/ob mice, including a significant decrease in liver TGs and improved scores in liver biopsies.
To further assess the quantitative decrease in liver steatosis in treated animals, a whole-body MRI was performed. This enabled a quantitative assessment of the amount of intrahepatic fat, which identified a decrease in the amount of fat accumulation in the liver of HP-treated P. obesus. In addition, a decrease in serum ALT and AST levels was noted in both models after treatment.
In this study, a trend for activation of P-STAT3 and increase in serum IL6 levels after treatment with HP extracts induced a potent anti-inflammatory effect, as noted by alleviation of liver injury in Con A immune-mediated hepatitis, and by alleviation of insulin resistance and liver damage in NASH in the P. obesus rats and leptin-deficient ob/ob mice models.
STAT3 regulates hepatic inflammation in a cell type-dependent manner during alcoholic liver injury. 44 STAT3 in hepatocytes promotes inflammation, whereas STAT3 in macrophages/Kupffer cells suppresses inflammation. 44 Ethanol-fed STAT3 KO mice showed more hepatic inflammation, worse injury, and increased hepatic expression of proinflammatory cytokines compared with wild-type mice. 44 Fructose-fed rats were hyperleptinemic and exhibited increased tyrosine phosphorylation of the STAT3 transcription factor compared with glucose-fed rats. 45 In contrast, mice that were fed a methionine/choline-deficient diet, which develop macrovesicular steatosis with minor necrosis and inflammation, manifested reduced phosphorylated STAT3 levels. 46 These conflicting results suggest that STAT3 plays distinct roles in the pathogenesis of NASH in different models.
The effect of HP extracts in our model of nutritionally induced T2DM was associated with a trend for an increase in serum IL6 levels. A threefold increase in serum IL6 levels was noted in HP-treated animals. IL6 induces hepatoprotection of steatotic liver isografts by preventing sinusoidal endothelial cell necrosis and consequent amelioration of hepatic microcirculation, thereby protecting against hepatocyte death. 29 IL6 pretreatment of steatotic Zucker rat liver isografts reduces mortality and liver injury after transplantation reperfusion. 29
In summary, HP-derived extract is associated with alleviation of insulin resistance, improved hepatic steatosis, and improved liver injury. The data support the potential liver protector effects of these extracts in NASH and other immune-associated liver disorders.
Author Disclosure Statement
Y. Ilan is the medical director of Plantylight, and a consultant for Teva, Enzo Biochem, Protalix, Therapix, Nasvax, and Natural Shield and Immuron.
Funding Information
This study was supported, in part, by Desert Labs Ltd, Plantylight, and The Roman-Epstein Liver Research Foundation.