
Abstract
Aging is associated with detrimental cellular and cognitive changes, making it an important public health concern; yet, many of these changes may be influenced by nutritional interventions. The natural sesquiterpene β-caryophyllene (BCP) has anti-inflammatory and antioxidant effects that are mediated by cannabinoid type-2 receptor activation, and these actions promote neuroprotection in different animal models that involve a cognitive damage. Consequently, whether chronic administration of BCP might prevent the age-related cellular and cognitive damage in a model of aging induced by chronic
Introduction
As stated by the World Health Organization, 1 aging is a natural process that cannot be stopped but that needs to be controlled to achieve healthy aging. Cognitive performance is one of the first abilities seriously affected by aging, 2 such as fluid abilities like cognitive flexibility. 3 Although there are several changes that contribute to age-related cognitive deficits, one of the first alterations involves the release of reactive oxygen species and reactive nitrogen species, subsequent oxidative and nitrosative stress, DNA mutations, and cell dysfunction and cell death. When the accumulation of these reactive species surpasses the capacity of the endogenous antioxidant defense system, a deleterious condition arises. 4,5
Several strategies aim to reduce age-related cognitive deficits, including the optimization of nutritional interventions with dietary supplementation, which not only improve cognition but also other biomarkers of aging. 6 β-Caryophyllene (BCP) is a natural sesquiterpene and a phytocannabinoid that has antioxidant 7,8 and an anti-inflammatory 8 –10 activity, achieved by activating the cannabinoid type-2 receptor (CB2R) in vitro and in vivo. Moreover, its administration prevents neuronal death in animal models of focal ischemia, 9 Parkinson's disease, 8,10 vascular dementia, 11 and Alzheimer's disease (AD). 12 In addition, BCP also improves spatial memory in rodents with AD 12 and vascular dementia. 11 These benefits have mainly been attributed to the activation of CB2Rs by BCP and the ensuing effects on the AMP-activated protein kinase/cAMP-response element binding/brain-derived neurotrophic factor, 13 peroxisome proliferator-activated receptor γ coactivator-1α/peroxisome proliferator-activated receptor-γ, 14 and nuclear factor erythroid 2-related factor 2/heme oxygenase-1 7,15,16 pathways. Consequently, this study aims to investigate whether chronic BCP administration could also prevent the cellular and cognitive damage associated with normal aging in an induced-aging model.
Materials and Methods
Chemicals
Animals and experimental design
Four-week-old male BALB/c mice were obtained from Western Biomedical Research Center (CIBO; Guadalajara, Mexico). Animals were transported to the laboratory and maintained under standard conditions on a 12 h light/dark cycle (8:00–20:00 h) with access ad libitum to food and water. They were handled in accordance with Mexican Federal Regulations for the Care and Use of Laboratory Animals (NOM-062-ZOO-1999), which are consistent with the EU guidelines in Directive 2010/63/EU. The mice were divided into four groups and administered the corresponding solutions daily: (1) saline solution (control group, CTRL), (2) BCP 10 mg/kg, administered orally, (3) GAL 300 mg/kg, administered i.p., and (4) GAL+BCP, where BCP treatment was introduced on the 4th week of GAL administration (Supplementary Data). Behavioral tests were performed in the 8th week of GAL administration, starting at 19:00 h, and the mice were killed 24 h later by perfusion and their brain were analyzed histologically to study the long-term effects of the treatments.
Open field test for locomotor activity
The open field test (OFT) was performed in a 40 × 40 cm cage for 5 min with a previous 5-min habituation period in the cage. The total distance and percentage of horizontal activity time were registered for each group (n = 10 mice per group).
Morris water maze test for long-term memory and cognitive flexibility
Long-term memory and cognitive flexibility were evaluated using the Morris water maze (MWM) test (n = 10 mice per group). The maze consisted of a black circular pool (100 cm diameter and 35 cm height) containing water to a depth of 30 cm and at 23 ± 2°C, and it was surrounded by four cardinally located distal cues. Mice were first habituated to the pool for 60 sec and they were then trained to find the platform (6.5 cm diameter and 29 cm height) located in a quadrant (Quadrant 1) over 5 days (four trials per day). Subsequently, the probe test of long-term memory evaluation was carried out 72 h later. 17 The reverse phase started the following day to evaluate cognitive flexibility. It consisted of eight learning trials performed in one day, in which the platform was placed in a new quadrant, Quadrant 2, 18 and again the probe test for cognitive flexibility evaluation was carried out 72 h later.
The latency to find the platform and the proportion of time spent in the target quadrants during the probe tests was assessed. The difference in the time spent in Quadrants 2 and 1 was calculated as the cognitive flexibility index, according to the following equation:
where t2/1 is the time spent in Quadrant 2 or Quadrant 1 during the probe test for cognitive flexibility and tT is the total time for the probe test.
Immunohistochemistry
Brains (n = 4 per group) were obtained from the perfused mice, embedded in paraffin, and 8 μm slices were obtained on a Leica RM2255 microtome, collecting every fourth slice for further analysis. The ventral and dorsal prefrontal cortex (vPFC and dPFC) were obtained from Bregma 1.98 mm and Bregma 1.42 mm, respectively, whereas the hippocampus was analyzed from Bregma −1.46 mm (Fig. 1). 19

Coronal view of the brain areas analyzed in this study.
Immunohistochemistry for glial fibrillary acidic protein (GFAP; Dako/Agilent, Santa Clara, CA, USA) or 8-oxo-2′-deoxyguanosine (8-oxo-dG; Abcam®, Cambridge, United Kingdom) was performed on both the PFC and hippocampal tissue. Samples were deparaffinized with xylol and passed through a descending gradient of ethanol. The sections for 8-oxo-dG staining were also digested for 10 min with 0.05% trypsin at 37°C (TrypLE Express Enzyme; Gibco®, Waltham, MA, USA) and then washed with 0.1 M phosphate-buffered saline (PBS). All the sections were then incubated in a 3.0% hydrogen peroxidase solution (J.T. Baker®/Avantor, Radnor, PA, USA), washed with PBS and blocked for 1 h with a solution containing 10% goat serum and 0.3% Triton X-100. The tissues were then incubated overnight with rabbit anti-GFAP or mouse anti-8-oxo-dG antibody diluted 1:200 in blocking solution (0.1 M PBS, 0.1% Triton X-100, and 10% goat serum). The following day the sections were washed with PBS and then incubated for 1.5 h with anti-rabbit or anti-mouse horseradish peroxidase-conjugated antibodies diluted 1:200 in PBS with 5.0% goat serum. Subsequently, the tissues were washed and then incubated with an avidin–biotin complex (ABC; VECTASTAIN®; Vector Labs, Burlingame, CA, USA) for 30 min in a dark room, and then washed and incubated with 3,3′-diaminobenzidine tetrahydrochloride. GFAP-stained sections were counterstained with hematoxylin and all the samples were finally washed again, dehydrated in graded ethanol, placed in xylene, and coverslipped using mounting medium.
Analysis of GFAP immunostaining
A Sholl analysis was performed at a 400 × amplification and using concentric circles from 10 to 100 μm in diameter. The total number of astrocytes and the total number of intersections were analyzed in six sections per brain area, in four mice per group. All the results were normalized to the CTRL for analysis.
Analysis of 8-oxo-dG immunostaining
Photographs were obtained from a total of six sections per brain area from four mice per group using the Motic® images plus 2.0 software (Kowloon, Hong Kong). Each photograph was color segmented to quantify the positive area using the free software ImageJ 20 and the results were normalized to the CTRL for analysis.
Statistical analysis
GraphPad Prism v.6 software (La Jolla, CA, USA) was used for statistical analysis. An analysis of variance and Tukey's post hoc test were used to analyze the behavioral tests, whereas a nonparametric analysis with the Kruskal–Wallis test, followed by a post hoc Mann–Whitney U-test was performed on the immunostaining data. All the results are shown as mean ± standard error of the mean and any differences were considered statistically significant at P < .05.
Results
Open field test
The OFT demonstrated that GAL and BCP administration did not influence the horizontal activity of the mice in terms of total distance (P = .1351) and their active time (P = .2590; Table 1).
Neither d-Galactose Nor β-Caryophyllene Impaired Locomotor Activity in the Open Field Test
Mean ± SEM; ANOVA test, n = 10.
ANOVA, analysis of variance; BCP, 10 mg/kg β-caryophyllene; CTRL, control group; GAL, 300 mg/kg
MWM test
Long-term memory and cognitive flexibility were evaluated in the normal and reverse phases of the MWM test, respectively. In the first phase, mice learned the platform's position in Quadrant 1 to assess their learning ability and long-term memory. Alternatively, a new position (Quadrant 2) had to be learned in the reverse phase to evaluate their cognitive flexibility (Fig. 2d). Neither GAL nor BCP affected the animal's learning ability (Fig. 2a) or long-term memory (Fig. 2b), as reflected by the latency and the time in the target quadrant during the normal phase of the MWM test, respectively.
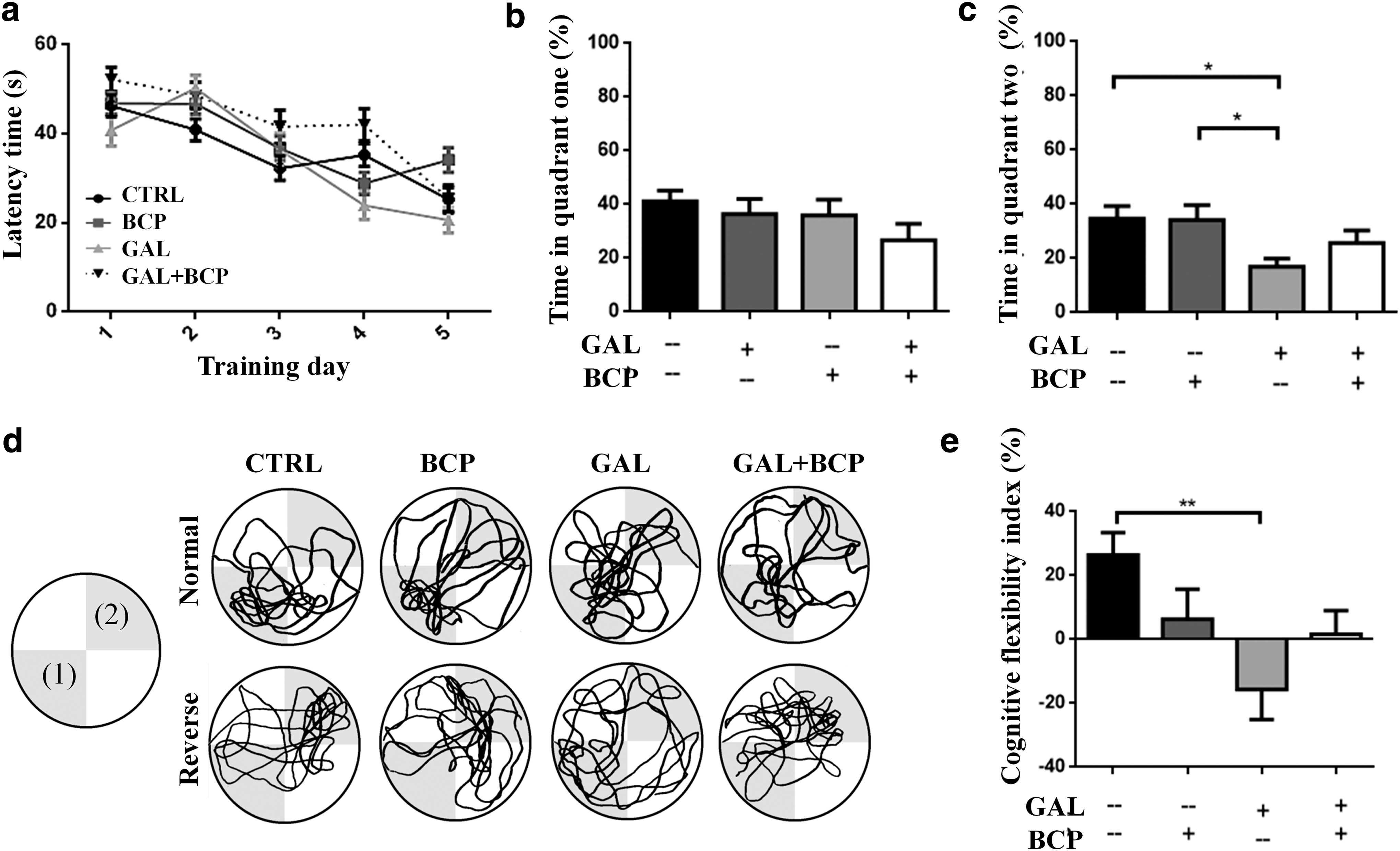
Impaired cognitive flexibility is promoted by GAL but not GAL+BCP.
Nonetheless, GAL treatment did alter cognitive flexibility, reflected as less time spent in the new target quadrant during the reverse phase of the MWM test relative to both the control mice (34.66 ± 4.60 vs. 16.84 ± 3.01, P = .0308) and those that received BCP (34.24 ± 5.43 vs. 16.84 ± 3.01, P = .0315; Fig. 2c). Moreover, there were no significant differences between the mice that received GAL alone and those that also received BCP (GAL+BCP) in terms of cognitive flexibility, although the mice that received GAL+BCP did not present the same alterations in this test as those that received GAL alone (Fig. 2c). The cognitive flexibility index was calculated to assess quadrant preferences in the reverse phase of the MWM test by calculating the difference between the search time in the first and second target quadrants (Fig. 2d; Eq. 1). The mice that received GAL alone displayed a deficit in learning the new platform position (cognitive flexibility), as they mainly searched for the platform at the first position learned and not in the second quadrant (Fig. 2d, e). This deficit was significant compared with the control mice that mainly looked for the platform in the second position rather than in the first one learned (−15.70 ± 9.39 vs. 26.42 ± 7.03, P = .0055; Fig. 2d, e). Furthermore, the same alteration was not evident when GAL was administered along with BCP in the GAL+BCP group (Fig. 2d, e).
GFAP immunostaining
There were significant differences in the total number of astrocytes in the Ammon's horn-3 (CA3; P = .0171), Ammon's horn-1 (CA1; P = .0101), and the dPFC (P = .0141) of the experimental groups when analyzed with a Kruskal–Wallis test. GAL treatment elevated the total number of astrocytes in both hippocampal areas relative to the controls: CA3 (176.20 ± 16.27 vs. 98.74 ± 6.17, P = .0286) and CA1 (149.10 ± 7.72 vs. 98.56 ± 12.48, P = .0286). However, BCP reverted the effect of GAL when they were administered together in the CA3 (100.50 ± 18.33 vs. 176.20 ± 16.27, P = .0286) but not the CA1 region (Table 2; Fig. 3). GAL administration also increased the total number of intersections in the hippocampal CA3 (137.50 ± 6.89 vs. 100.20 ± 8.42, P = .0286) and CA1 (162.30 ± 14.12 vs. 100.30 ± 8.64, P = .0286) regions and although BCP coadministration seemed to dampen the effect of GAL in the CA3 region, this effect was not significant (98.60 ± 7.98 vs. 137.50 ± 6.89, P = .0571; Table 3).
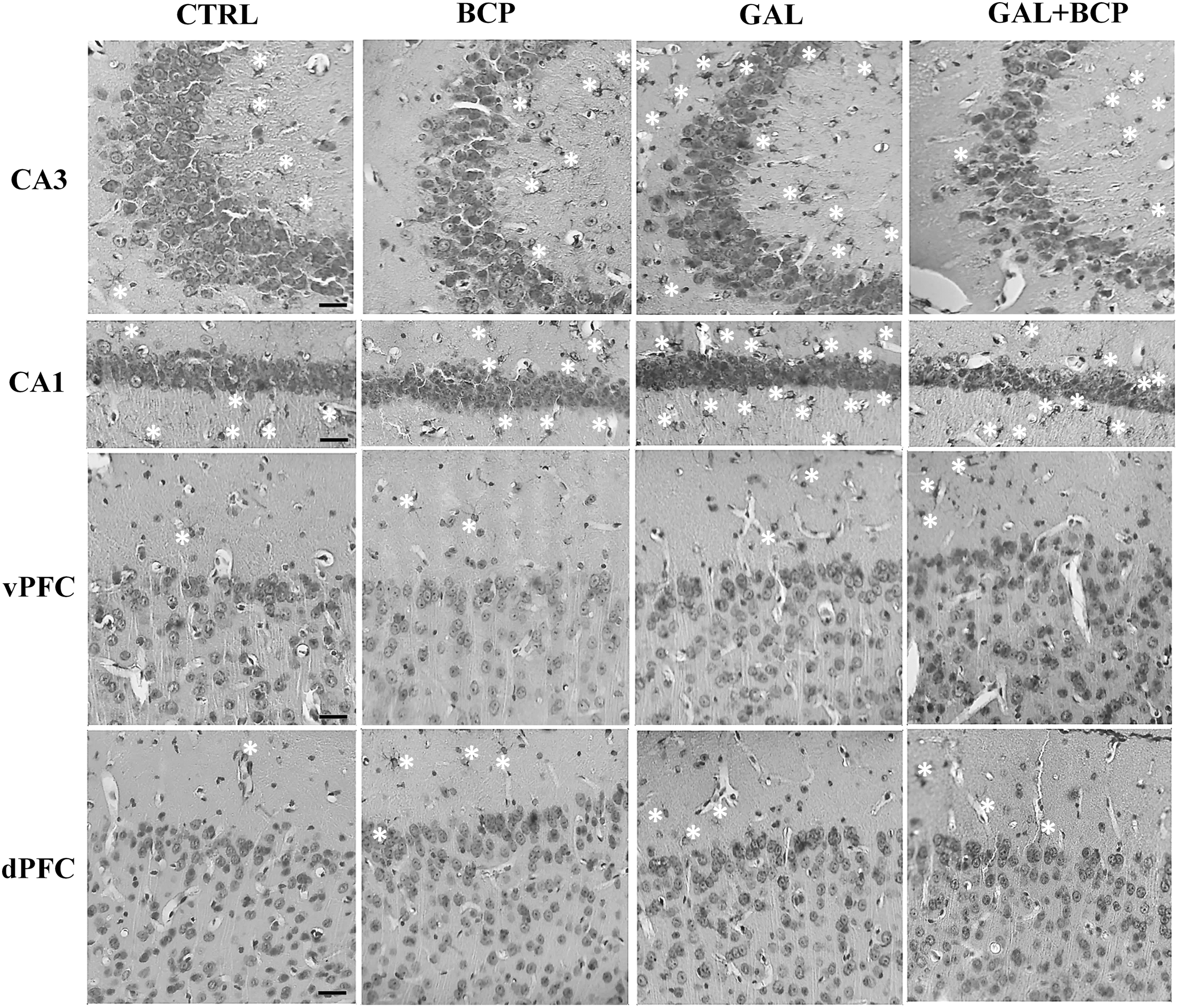
GFAP immunostaining counterstained with hematoxylin. GAL treatment increased the number of astrocytes (white asterisk) in the CA3 and CA1 relative to the controls, a phenomenon reversed by BCP administration in the CA3 region (Table 2). BCP treatment alone significantly elevated both the total number of astrocytes and the total number of astrocyte intersections in the dPFC relative to the controls (Tables 2 and 3). Bar: 50 μm. CA1, Ammon's horn-1; CA3, Ammon's horn-3; GFAP, glial fibrillary acidic protein.
The Increase in the Number of Astrocytes in the Hippocampus After d-Galactose Administration Was Reversed by β-Caryophyllene Treatment
Mean ± SEM, n = 4; Kruskal–Wallis test with Mann–Whitney U post hoc test.
P value is equal to 0.0286 between groups with different letters (a,b).
P < 0.05.
CA1, Ammon's horn-1; CA3, Ammon's horn-3; dPFC, dorsal prefrontal cortex; vPFC, ventral prefrontal cortex.
An Increase in the Total Number of Astrocyte Intersections in a Sholl Analysis Provoked by d-Galactose in the Hippocampus and by β-Caryophyllene in the Cortex
Mean ± SEM, n = 4; Kruskal–Wallis test with Mann–Whitney U post hoc test.
P value is equal to 0.0286 between groups with different letters (a,b).
P < 0.05.
In the PFC, a significant difference in both the total number of astrocytes (P = .0141) and the total number of intersections (P = .0262) was only observed in the dPFC with the Kruskal–Wallis test. In this region, BCP treatment alone promoted a significant elevation in the total number of astrocytes (460.50 ± 72.89 vs. 99.94 ± 51.48, P = .0286) and also in the total number of intersections (622.50 ± 166.30 vs. 97.25 ± 37.03, P = .0286) relative to the CTRL (Tables 2 and 3; Fig. 3).
8-oxo-dG Immunostaining
DNA damage was evaluated by measuring its oxidation through 8-oxo-dG immunostaining. In contrast to GFAP immunostaining, GAL treatment alone promoted more DNA oxidation in the dPFC but not in the hippocampus relative to the control mice (1627.00 ± 774.20 vs. 100.00 ± 87.13, P = .0286). Moreover, the coadministration of BCP successfully reversed the GAL-induced damage in the GAL+BCP mice (282.70 ± 99.68 vs. 1627.00 ± 774.20, P = .0286; Table 4; Fig. 4). These data suggest that the BCP treatment helps to dampen the oxidative damage induced by the chronic GAL administration that induces aging.
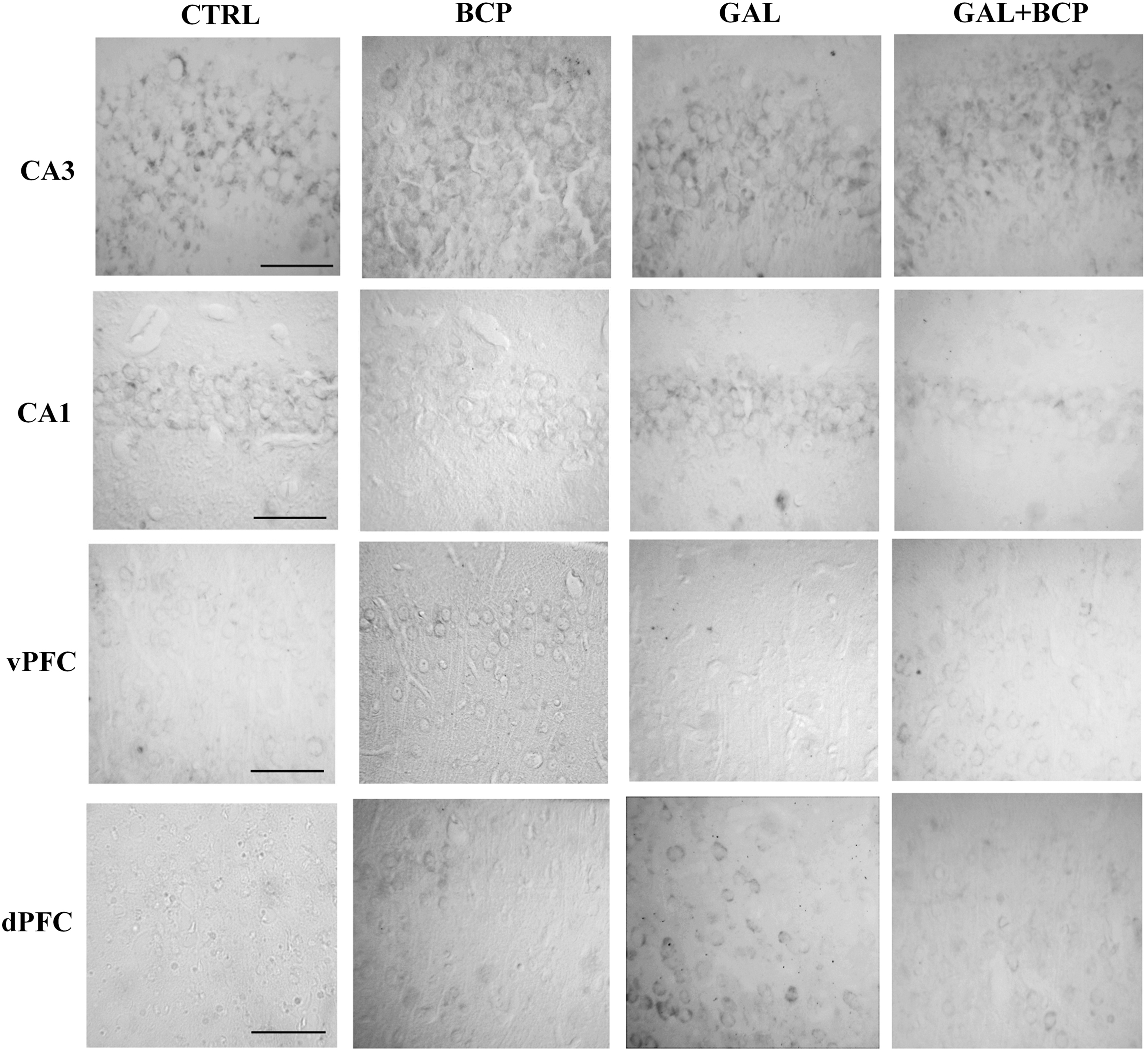
8-oxo-dG immunostaining. GAL treatment elevated the dPFC DNA oxidation relative to the controls, a phenomenon reversed by BCP coadministration (GAL+BCP group; Table 4). Bar: 100 μm. 8-oxo-dG, 8-oxo-2′-deoxyguanosine.
β-Caryophyllene Treatment Reversed d-Galactose-Induced DNA Oxidation in Dorsal Prefrontal Cortex as Evaluated with 8-Oxo-2′-Deoxyguanosine Immunostaining
Mean ± SEM, n = 4; Kruskal–Wallis test with Mann–Whitney U post hoc test.
P value is equal to 0.0286 between groups with different letters (a,b).
P < 0.05.
Discussion
The GAL model of induced aging has been used as an alternative to naturally occurring and genetically modified aged animals, as it offers advantages in terms of time and costs. The administration of 100–150 mg/kg GAL to BALB/c mice for 6–8 weeks provokes a prooxidant and proinflammatory environment in different organs, including the brain. 21 –25 Moreover, the administration of a higher dose of GAL (300 mg/kg) impairs cognitive flexibility (Supplementary Fig. S1) but not locomotor activity, long-term memory, or neuronal death (Supplementary Fig. S2 and Supplementary Table S1). The decline in such capacities diminishes quality of life in the elderly, and cognitive flexibility is considered to be an attribute affected early in aging. 2,3 Activity in the PFC and hippocampus is tightly related to cognitive flexibility and thus, any impairment to one or both of these regions may be reflected by a deterioration in this capacity. 26,27 Hence, the GAL mice mimicked early aging as they not only presented more extensive brain tissue damage, evident as an increase in astrocytic reactivity and DNA oxidation (GFAP and 8-oxo-dG staining, respectively) in the hippocampus and PFC, particularly in the CA3 and dPFC but also, impaired cognitive flexibility.
Chronic administration of BCP improved most of the detrimental changes induced by GAL here. BCP is a cannabinoid with anti-inflammatory and antioxidant activities that are mainly mediated by specific activation of CB2Rs, widely considered as a damage-inducible receptor 10 –12,14,28,29 that is expressed strongly by microglia, followed by astrocytes and finally neurons in the central nervous system (CNS). 30 CB2R activation in the CNS reduces glial activation and neuronal damage. 8,10,12 The reduction of astrocyte activation and prevention of DNA oxidation through BCP administration in the mice that received GAL could be because of the activation of CB2R-dependent pathways, promoting neuroprotection. Indeed, this natural-occurring cannabinoid improves memory and cognition when administered chronically 12,14 because of the activation of neurotrophic factor signaling, 13 in addition to its effect on cell survival 13 driven by reducing neuroinflammation 14 and by its antioxidant activity. 7,11,15 In the GAL+BCP mice, less GAL-induced damage was detected in both the hippocampus and PFC; yet, the protection exerted by chronic administration of BCP (10 mg/kg) was insufficient to produce improvements in cognitive flexibility. Thus, it will be of significant interest to determine why the protection afforded by BCP does not significantly improve cognitive flexibility.
Finally, administration of BCP alone had no effect on long-term memory or cognitive flexibility. This contrasts with the effects of cannabinoid type-1 receptor (CB1R) agonists like HU-210 and Δ 9 -tetrahydrocanabinol that impair these phenomena, 31,32 demonstrating its advantage over the therapeutic use of CB1R agonists. However, it is notable that BCP alone did augment the number of astrocytes and their total number of intersections. Hence, more studies will be necessary to determine if this phenomenon, already reported with other antioxidant molecules like vitamin C, 33 is mainly because of the absence of damage-inducible CB2R activity when BCP was administered alone to healthy mice. Alternatively, BCP administration might improve the metabolic functions of astrocytes and not their reactive responses. 34
In summary, BCP reverts the detrimental changes in astrocytes and DNA oxidation induced by chronic GAL administration, in addition to its other known biological effects. However, its effect on cognitive flexibility remains to be investigated.
Footnotes
Acknowledgments
The authors thank Prof./Dr. Jürg Gertsch from the Institute of Biochemistry and Molecular Medicine, University of Bern, Switzerland for kindly providing the β-caryophyllene used in these experiments.
Author Disclosure Statement
No competing financial interests exist in relation to this study.
Funding Information
This project was supported by the Consejo Nacional de Ciencia y Tecnología (CONACYT) by Student Grant No. 401378.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.