
Abstract
Metabolic syndrome (MetS) is a combination of risk factors that include insulin resistance, obesity, dyslipidemia, and hypertension. The consumption of high-fructose diets contributes to the development of MetS. β-sitosterol a naturally occurring phytosterol possesses antiobesogenic and antidiabetic effects. This study evaluated the potential protective effect of β-sitosterol against the development of metabolic dysfunction in growing female rats fed a high-fructose diet, mimicking children fed obesogenic diets. Thirty-five 21-day-old female Sprague Dawley rat pups were randomly allocated to and administered the following treatments: group 1—standard rat chow (SRC) + plain drinking water (PW) + plain gelatine cube (PC); group 2—SRC + 20% w/w fructose solution (FS) as drinking fluid + PC; group 3—SRC + FS + 100 mg/kg fenofibrate in gelatine cubes; group 4—SRC + FS + 20 mg/kg β-sitosterol gelatine cube (Bst); and group 5—SRC + PW + Bst. Following 12 weeks of feeding, the rats were fasted overnight, weighed, and then euthanized. Plasma cholesterol, insulin, glucose, triglyceride, and adiponectin concentrations were determined. Visceral fat was dissected out and weighed. The high-fructose diet increased (P < .05) visceral adiposity and plasma triglyceride concentration but decreased (P < .05) plasma adiponectin concentration. β-sitosterol prevented the high-fructose diet-induced visceral obesity, hypertriglyceridemia, and hypoadiponectinemia. β-sitosterol alone increased plasma adiponectin concentration and reduced plasma insulin concentration and homeostatic model assessment index. In conclusion, β-sitosterol could be potentially used to prevent high-fructose diet-induced metabolic dysfunction.
Introduction
The World Health Organization reported an increase in the prevalence of obesity among infants and young children (aged 0–5 years) from 32 million globally in 1990 to 41 million in 2016. 1 If this trend continues globally, it is projected that the number of overweight or obese infants and young children will increase to 70 million by 2025. 1 The prevalence of obesity is reported to be higher in female children compared to male children. 2 Obesity is a major risk factor for the development of metabolic syndrome (MetS). 3 –5 MetS is defined as the concurrent manifestation of risk factors that increase the risk of developing type II diabetes mellitus (T-II-DM) and cardiovascular disease. 6 These risk factors include insulin resistance, visceral adiposity, atherogenic dyslipidemia, and elevated blood pressure. 7 Plasma triglyceride concentration and visceral obesity have been identified as accurate indicators for screening and predicting the risk for MetS and T-II-DM. 8 –10 Sedentary lifestyles and the consumption of obesogenic diets have been identified as the major causes of obesity and MetS. 11
Fructose is used as a sweetening agent in various processed foods and beverages 12 and due to poor dietary habits, there is an increase in the consumption of fructose-laden foods. The metabolism of fructose bypasses the phosphofructokinase rate-limiting step in hepatocytes and thus results in increased hepatic de novo lipogenesis (DNL). Increased lipogenesis contributes to the development of obesity and MetS. 13 –15
Some features of MetS can be treated or managed through the use of synthetic pharmaceutical agents such as fenofibrate, an antihyperlipidemic drug. 16 However, synthetic pharmacological agents are costly and elicit deleterious side effects. 17 This has resulted in a worldwide increase in the use of natural plant-derived ethnomedicines. 16 Plant-derived ethnomedicines have been and continue to be used in disease management and their role in primary health care is expanding. 16 Ethnomedicinal plants contain phytochemicals such as phytosterols that are known to have multiple biological benefits. 16 –18 β-sitosterol is one of the most abundant and widely distributed phytosterols. 19 It possesses antiobesogenic, anti-inflammatory, and antidiabetic properties. 20,21 β-sitosterol has been shown to increase the release of the anti-inflammatory cytokine (interleukin-10) in macrophage cells. 22,23 Orally administered β-sitosterol [at 10, 15, and 20 mg/(kg·d)] for 21 days decreased glycated hemoglobin, serum glucose, and nitric oxide in streptozotocin-diabetic adult rats. 20 These antidiabetic effects of β-sitosterol were attributed to its antioxidant activity and its ability to stimulate the regeneration of pancreatic β-cells. 20 In a clinical study, the consumption of phytosterols was shown to be effective in treating hyper-low-density lipoprotein (LDL)-cholesterolemia in hypercholesterolemic patients. 24 β-sitosterol exerted its antihyperlipidemic effects through the upregulation of cellular AMP-protein kinase (AMPK) activity, which suppresses DNL, increases fatty acid oxidation, and promotes adipose tissue mitochondrial integrity. 25
Despite the evidence that obesity, MetS, and T-II-DM are increasing in children, the beneficial effects of β-sitosterol on metabolic health have been investigated mainly in adult animal models and adult human studies. Moreover, a large proportion of these studies have investigated the curative properties of β-sitosterol and not its prophylactic potential. Globally, it is well recognized that prevention of diseases reduces the burden of health care to a greater extent than the cure of diseases. Furthermore, most studies investigating diet-induced metabolic dysfunction have a tendency to use male animal models. 26 However, the prevalence of obesity is reported to be higher in female children compared to male children. 2 Thus, there is a need for studies that will interrogate the pathophysiology of diet-induced metabolic dysfunction and the potential plant-derived interventions initially using female and male animal experimental models before conducting human trials. This study, therefore, investigated the potential of orally administered β-sitosterol to prevent high-fructose diet-induced metabolic dysfunction in growing female rats mimicking children fed obesogenic diets.
Materials and Methods
Ethical approval
Ethical clearance for the study was granted by the Animal Ethics Screening Committee (AESC) of the University of Witwatersrand (AESC clearance number 2017/08/55/B). This study was conducted in accordance with the internationally accepted principles and guidelines on the care and use of laboratory animals (National Research Council, 2010).
Chemicals and reagents
All the chemicals and reagents used in the study were of analytical grade. β-sitosterol, fenofibrate, and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich (St. Louis, MO, USA). The fructose was obtained from Nature's Choice (Randvaal, South Africa).
Animals, housing and general care
Thirty-five 21-day-old female Sprague Dawley rat pups were used in the experiment. Each rat was individually housed in a perspex cage with stainless steel mesh lids in the Central Animal Services, University of the Witwatersrand. The cages were lined with wood shavings for bedding. A 12-h light and dark cycle was followed with lights on from 7 am to 7 pm. Room temperature was maintained at 26 ± 2°C. The rats had ad libitum access to a standard rat chow (SRC) [Nutritionhub (PTY) Ltd., Stellenbosch, South Africa] and drinking fluid [water or 20% fructose solution (FS) as described below] throughout the experiment. The rat chow had the following nutrition content: calcium 14 g/kg, phosphorus 8 g/kg, vitamin A (min) 16,000 IU/kg, vitamin D 2000 IU/kg and vitamin E 100 mg/kg, crude protein 240 g/kg, crude oils and fats 53 g/kg, linoleic acid 14 g/kg, crude fiber 45 g/kg, and crude ash 75 g/kg. The rat pups were acclimatized to handling for 2 days [postnatal days (PNDs) 21 and 22] before the commencement of the experiment on PND 23.
Experimental design
On PND 23 the 35 pups were randomly distributed and fed the following treatment regimens for 12 weeks: group 1, the negative control (n = 7)—SRC + plain drinking water (PW) + plain gelatine cube (PC), group 2 in which metabolic dysfunction was induced (n = 7)—SRC + 20% w/w FS to drink + PC, group 3 in which fenofibrate was used a positive control for the prevention of metabolic dysfunction (n = 7)—SRC + FS + 100 mg/kg body mass per day of fenofibrate in a gelatine cube (FF), group 4 in which the potential prophylactic effects of β-sitosterol against high-fructose diet-induced metabolic dysfunction were explored (n = 7)—SRC + FS + 20 mg/kg body mass per day of β-sitosterol in a gelatine cube (Bst), and group 5 in which the impact of β-sitosterol alone on the metabolic health of the rats was explored (n = 7)—SRC + PW + Bst.
The fenofibrate and β-sitosterol doses used were similar to those previously described by Hong et al. 27 and Baskar et al. 28 for nonalcoholic fatty liver disease and colon cancer respectively. Due to the poor aqueous solubility of β-sitosterol, DMSO (0.5%) was used as a vehicle to dissolve β-sitosterol and the fenofibrate. 28,29 The PCs in the control groups also contained (0.5%) DMSO. The gelatine cubes were prepared as previously described by Kamerman et al. 30 with modification. The gelatine cubes were prepared using 8 g brown sugar (Huletts, Tongaat, South Africa Ltd.) instead of 16 g. Throughout the experiment, the rats were weighed twice weekly to ensure the maintenance of a constant dose of the interventions.
Terminal procedures and sample collection
Following 12 weeks of the administration of the treatments, the rats were fasted overnight but allowed access to PW. The rats were weighed and fasting blood glucose and triglyceride concentration was determined from a drop of blood obtained from the tail vein using a pinprick. 31 Blood glucose and triglyceride concentration were measured using a calibrated glucose meter (Contour Plus Meter, Bayer, Isando-Johannesburg, South Africa) and a calibrated Accutrend triglyceride meter (Roche, Mannheim, Germany), respectively. Immediately thereafter the rats were euthanized with an intraperitoneal injection of sodium pentobarbitone at 200 mg/kg body mass. Following euthanasia, blood samples were collected, via cardiac puncture using 18G needles and 10 mL syringes, into heparinized blood tubes (Becton Dickinson Vacutainer Systems Europe, Meylan Cedex, France). The blood samples were centrifuged 20°C for 15 min at 5000 g in a Sorvall RT® 6000B centrifuge (Pegasus Scientific, Inc., Rockville, MD, USA). The harvested plasma was stored in microtubes at −20°C pending determination of plasma cholesterol, insulin, and adiponectin concentration.
The abdomen of each rat was opened via a midline incision and the visceral fat pad was dissected out and weighed. The visceral fat mass related to body mass (% BM) was computed according to the formula: visceral fat mass (g)/terminal body mass (g) × 100.
Determination of adiponectin and insulin concentration
Fasting adiponectin (pg/mL) and insulin (ng/mL) concentration were measured using an adiponectin and insulin enzyme-linked immunosorbent assay (ELISA) kit for rats (Elabscience Biotechnology Co, Wuhan, Hubei, China). A standard curve was constructed using calibrator concentrations. The concentrations of insulin and adiponectin in the samples were then determined from the constructed standard curve. Insulin resistance was estimated by computing the homeostatic model assessment index (HOMA-IR) using the fasting blood glucose and insulin concentration, by using the equation
HOMA-IR = insulin (ng/mL) blood glucose (mmol/L)/22.5. 32
Determination of cholesterol
The plasma cholesterol concentrations were determined using a colorimetric-based clinical chemistry analyzer (IDEXX VetTest® Clinical Chemistry Analyzer; IDEXX Laboratories, Inc., USA) as per the manufacturer's instructions.
Statistical analysis
GraphPad Prism 6.0 (Graph-pad Software, Inc., San Diego, CA, USA) software was used to analyze data. Data are expressed as mean ± standard deviation. Data on weekly body mass were analyzed using a repeated measures analysis of variance (ANOVA). Multiple-group data were analyzed using one-way ANOVA, followed by Tukey's post hoc test. Significance was accepted when P < .05.
Results
Body mass
Figure 1 shows the initial and terminal body masses of female rats across treatment regimens. The initial body masses of the rats were similar (P > .05) (Fig. 1). Following the 12-week intervention across treatment regimens the rats had a significant increase (P < .0001) in body mass (Fig. 1). However, the terminal body masses were similar (P > .05) across treatments (Fig. 1).
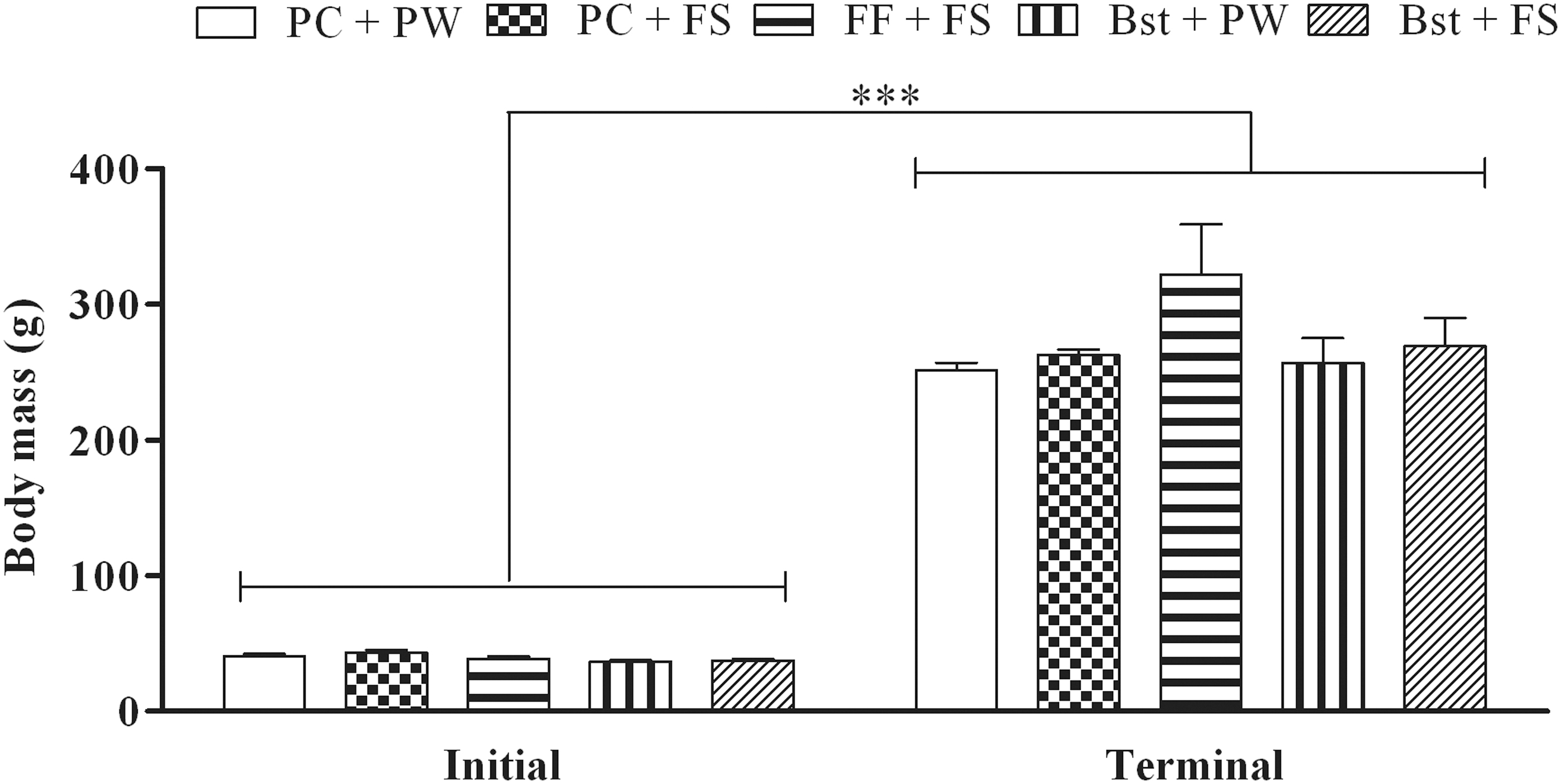
Initial and terminal body masses of the female rats. Data presented as mean ± SD; n = 7 per treatment. ***P < .0001; ANOVA, Tukey's post hoc test. ANOVA, analysis of variance; Bst + FS, gelatine cube containing β-sitosterol at a dose of 20 mg/kg body mass per day +20% (w/w) fructose solution to drink; Bst + PW, gelatine cube containing β-sitosterol at a dose of 20 mg/kg body mass per day + plain tap water to drink; FF + FS, gelatine cube containing fenofibrate at a dose of 100 mg/kg body mass per day +20% (w/w) fructose solution to drink; PC + FS, plain gelatine cubes +20% (w/w) fructose solution to drink; PC + PW, plain gelatine cubes + plain tap water to drink; SD, standard deviation.
Visceral obesity
Figure 2 shows the absolute (A) and relative (B) visceral fat pad masses of female rats following respective treatments. Rats that were fed the high-fructose diet (PC + FS) had significantly (P < .001) increased visceral fat mass (absolute and relative to body mass) compared to their control (PC + PW) counterparts (Fig. 2). β-sitosterol (Bst + FS) prevented (P < .001) the high-fructose diet-induced increase in absolute and relative (to body mass) visceral fat mass (Fig. 2). Rats fed fenofibrate (FF + FS) had significantly lower (P < .05) absolute visceral fat mass compared to those on fructose alone (PC + FS) (Fig. 2).

Effect of β-sitosterol on absolute
Triglyceride and cholesterol concentration
Figure 3 shows the plasma triglyceride (A) and cholesterol (B) concentration in the female rats following their respective treatments. Rats that were fed high-fructose diet alone (PC + FS) had significantly increased (P < .0001) plasma triglyceride concentration when compared to their control (PC + PW) counterparts (Fig. 3). β-sitosterol (Bst + FS) prevented (P < .0001) the high-fructose diet-induced hypertriglyceridemia (Fig. 3). Fenofibrate (FF + FS) failed to prevent (P > .05) the high-fructose diet-induced hypertriglyceridemia (Fig. 3). Although the high-fructose diet had not significantly increased (P > .05) the plasma total cholesterol concentration (Fig. 3), the high-fructose diet-fed rats had 26% higher (P = .08) plasma total cholesterol compared to that of rats fed the control diet. Rats fed the high-fructose diet in combination with fenofibrate (FF + FS) or β-sitosterol (Bst + FS) had significantly higher (P < .05) plasma cholesterol concentration compared to their control (PC + PW) counterparts (Fig. 3). Rats that fed β-sitosterol alone (Bst + PW) had significantly lower (P < .001) plasma cholesterol concentration compared to their counterparts who fed the high-fructose diet (PC + FS; Bst + FS; FF + FS) (Fig. 3).

Effect of β-sitosterol on plasma triglyceride
Glucose, insulin, adiponectin, and HOMA-IR
Figure 4 shows the glucose (A), insulin (B), and adiponectin (C) concentration and HOMA-IR (D) of the female rats following their respective treatments. The high-fructose diet had no effect on glucose and insulin concentration and HOMA-IR (Fig. 4). Rats fed the high-fructose diet (PC + FS) had an undetectable adiponectin concentration (Fig. 4), thus were assigned the lowest detectable ELISA adiponectin concentration of 46.88 pg/mL. Rats fed the high-fructose diet in combination with β-sitosterol (Bst + FS) or fenofibrate (FF + FS) had significantly higher (P < .0001) plasma adiponectin concentration compared to their control (PC + PW) and high-fructose diet (PC + FS) counterparts (Fig. 4). β-sitosterol alone (Bst + PW) significantly increased (P < .001 compared to PC + PW and PC + FS) plasma adiponectin concentration and lowered (P < .001 compared to all other treatments) plasma insulin concentration and HOMA-IR (Fig. 4).
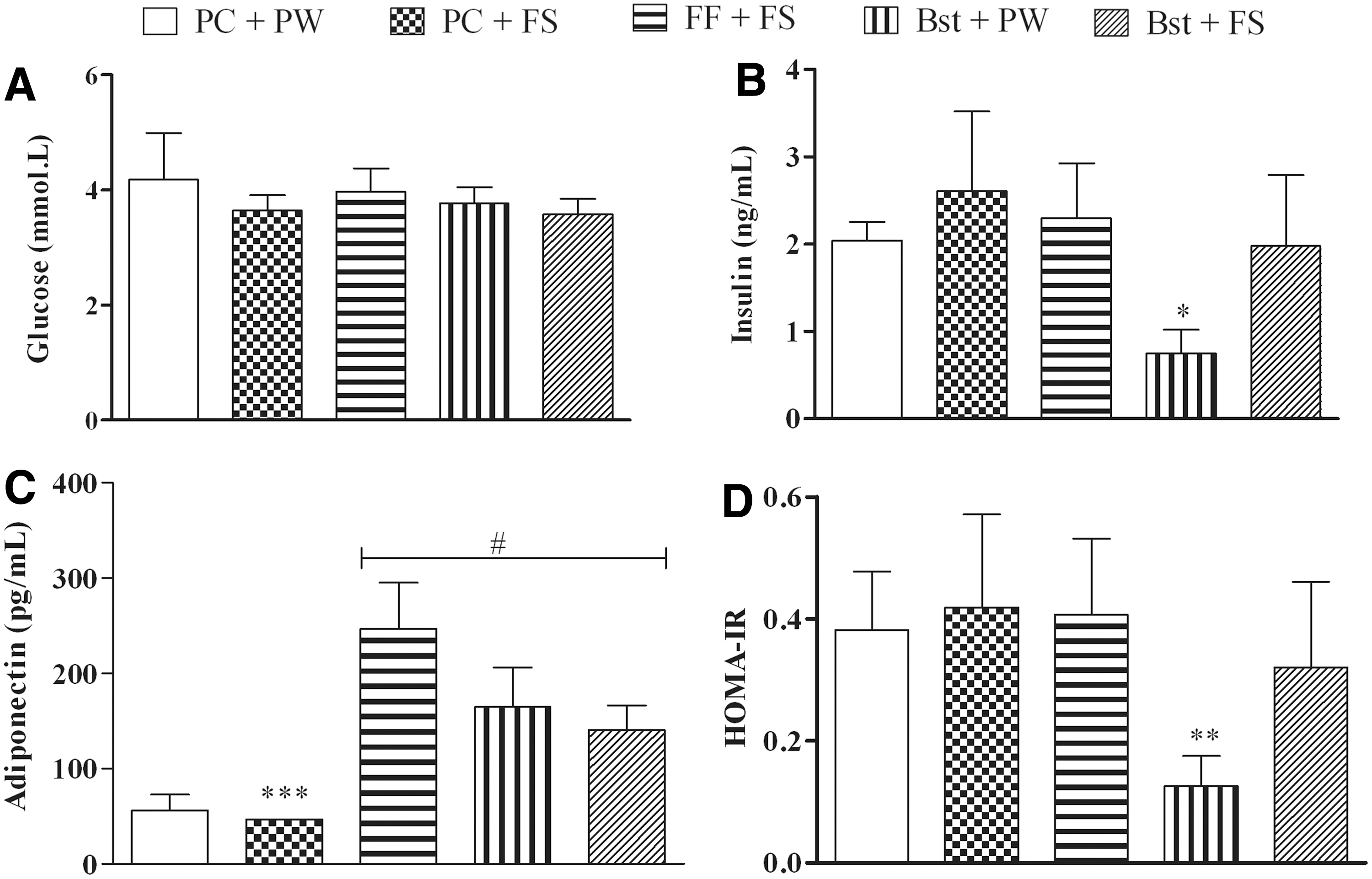
Effect of β-sitosterol on glucose
Discussion
The study aimed to investigate the potential of orally administrated β-sitosterol to protect growing female rats against the development of fructose-induced metabolic dysfunction. Results from the current study showed that the high-fructose diet had no effect on body mass gain of female rats (Fig. 1). These findings are in agreement with Ramos et al. who reported that the consumption of 20% fructose diet for 8 weeks had no effect of body mass gain but caused increased visceral adiposity. 33 Ramos et al. contended that the observed increase in adiposity without a concomitant increase in body mass to be a consequence of the high adipogenic and low energetic nature of fructose. 33 We therefore speculate that the observed dichotomy between the body mass and visceral adiposity in the current study is a result of the low energy efficiency of fructose. β-sitosterol and fenofibrate had no effect on body mass gain in the current study (Fig. 1). Therefore, these findings suggest that β-sitosterol and fenofibrate had no effect on body mass gain in the current study (Fig. 1). Body mass gain is an important indirect indicator of an animal's health status and therefore, these findings suggest that β-sitosterol and fenofibrate did not negatively impact the rats' health as indicated by body mass. In the present study, the consumption of a high-fructose diet for 12 weeks by female rats resulted in metabolic dysfunction characterized by visceral obesity, hypoadiponectinemia, and hypertriglyceridemia. β-sitosterol protected the female rats against the development of high-fructose diet-induced metabolic dysfunction. Fenofibrate only prevented the high-fructose diet-induced visceral obesity and hypoadiponectinemia.
In the liver, the metabolism of each molecule of fructose produces three carbon intermediates, which can enter the glycolytic pathway. 34 These intermediates may be interconverted or further metabolised to acetyl-CoA. 35 Acetyl-CoA can be converted to malonyl-CoA, a metabolite that is utilized in de novo triglyceride synthesis. 36 The acetyl-CoA can also form HMG-CoA and undergo a series of reactions to form cholesterol. 37 The newly synthesized triglycerides are packed into very low-LDL and transported to the peripheral adipose stores and thus causing dyslipidemia and visceral obesity. 38,39 In the current study, the high-fructose diet caused hypertriglyceridemia and visceral adiposity (Figs. 2 and 3). These findings suggest that the high-fructose diet caused de novo triglyceride synthesis 40 that resulted in hypertriglyceridemia and visceral adiposity. 41 Several studies have reported that MetS can be accurately diagnosed using only plasma triglyceride concentration and visceral obesity. 9,10 Thus, hypertriglyceridemia and visceral obesity are key predictors of MetS. In the current study, consumption of the high-fructose diet increased (P > .05) mean plasma cholesterol by 27% when compared to control (Fig. 3). It has been reported that a 10% change in total cholesterol concentration is clinically relevant. 42 A 10% increase in cholesterol significantly increases the risk of coronary heart disease-related mortality. 42 Thus, findings of the current study suggest that the high-fructose diet might have hypercholesterolemic effects.
In the present study, β-sitosterol prevented the high-fructose diet-induced hypertriglyceridemia and visceral obesity (Figs. 2 and 3). These antiobesogenic effects of β-sitosterol might be attributed to its ability to decrease hepatic DNL through AMPK activation. 25 In the current study, cholesterol increased when β-sitosterol was administered to high-fructose diet-fed rats (Fig. 3). However, when β-sitosterol was administrated alone it resulted in a 12% decrease in mean plasma cholesterol concentration compared to control. In vitro, β-sitosterol was reported to decrease cholesterol synthesis by decreasing HMG-CoA reductase activity. 43 Thus, it could be speculated that in this study the administration of β-sitosterol exacerbated the hypercholesterolemic effects of the high-fructose diet. A possible explanation could be that β-sitosterol and fructose, together, have synergistic effects that consequently resulted in the hypercholesterolemia. At the moment the mechanisms involved in these findings are unknown and thus further investigation are needed.
Fenofibrate, an antihyperlipidemic drug, was used as a positive control in the current study. It exerts antiobesogenic effects by stimulating peroxisome proliferator-activated receptor α (PPAR-α)-mediated lipolysis and fatty acid β-oxidation in adipose tissues. 44 PPAR-α activates genes that regulate lipoprotein lipase, carnitine palmitoyltransferase, and acyl-Co-A oxidase activity in the liver and in adipocytes. 45,46 Lipoprotein lipase hydrolyses triglycerides into fatty acid and glycerol. 47,48 Carnitine palmitoyltransferase facilitates free fatty acids uptake into the mitochondria. Inside the mitochondria, acyl-Co-A oxidase promotes fatty acid β-oxidation 48 In the current study, fenofibrate prevented the high-fructose diet-induced visceral obesity (Fig. 2). This finding is in agreement with Jeong and Yoon who reported that fenofibrate decreased visceral fat mass in high fat diet-fed obese mice. 44 It could be speculated that the decrease in visceral fat mass in the current study following fenofibrate administration was due to upregulated triglyceride hydrolysis and fatty acid β-oxidation. In contrast to previous studies, fenofibrate failed to prevent the high-fructose diet-induced hypertriglyceridemia and hypercholesterolemia in the current study. Fructose causes dyslipidemia by increasing DNL in the liver. 40 Thus, it could be speculated that fenofibrate was unable to prevent the high-fructose diet-induced DNL in the liver.
Several studies have reported that visceral adiposity is associated with decreased adiponectin concentration. 49,50 Visceral adiposity increases the production of tumor necrosis factor-α, 51 which in turn inhibits adiponectin gene expression. 52 Adiponectin, an anti-inflammatory adipokine, increases insulin sensitivity by stimulating the activity of PPAR-α and AMPK. 53 In the present study, rats fed a high-fructose diet showed undetectable adiponectin concentration (Fig. 4). This finding suggests that the high-fructose diet exerted hypoadiponectinemia effects (Fig. 4). The high-fructose diet might have decreased adiponectin by causing visceral adiposity. 41 HOMA-IR is used to assess insulin resistance. 54 Although the high-fructose diet decreased adiponectin, it had no effect on insulin concentration and HOMA-IR (Fig. 4).
In the current study, the oral administration of β-sitosterol alone or with the high-fructose diet resulted in increased plasma adiponectin concentration (Fig. 4). β-sitosterol may have exerted its hyperadiponectinemia effects through the reduction of visceral adiposity. Collins et al. reported that the hyperadiponectinemia effect of plant sterols is attributed to their anti-inflammatory activities. 55 The administration of β-sitosterol alone significantly decreased plasma insulin concentration and increased insulin sensitivity (Fig. 4). These results are in tandem with those by Radika et al. who reported reduced insulin concentration and improved insulin sensitivity following 4 weeks administration of β-sitosterol to adult Wistar rats fed a high-fat diet. 56 β-sitosterol reduced plasma insulin concentration and HOMA-IR by exerting anti-inflammatory effects and by decreasing the expression of inducible nitric oxide synthase (iNOS) that interferes with insulin signaling. 57 Therefore, it could be speculated that β-sitosterol decreased plasma insulin concentration and increased insulin sensitivity through anti-inflammatory mechanisms and suppressing iNOS-induced insulin signaling interference. The oral administration of fenofibrate significantly increased plasma adiponectin concentration but had no effect on insulin concentration and HOMA-IR (Fig. 4); thus, it can be speculated that fenofibrate increased adiponectin concentration by decreasing visceral adiposity.
Conclusion
We have shown that feeding a high-fructose diet to growing female rats for 12 weeks caused metabolic dysfunction characterized by visceral adiposity, hypertriglyceridemia, and hypoadiponectinemia. β-sitosterol protected the growing female rats against the development of high-fructose diet-induced metabolic dysfunction. In conclusion, β-sitosterol could potentially be exploited as a prophylactic agent against diet-induced metabolic dysfunction in children.
Footnotes
Acknowledgments
We thankfully acknowledge the Faculty of Health Sciences Research Committee of the University of the Witwatersrand, Johannesburg and the National Research Foundation of South Africa for funding the research. We also wish to acknowledge the staff of the University of the Witwatersrand Central Animal Service unit for assistance with the care and welfare of the rats. The authors also wish to acknowledge Ms. Monica Gomez for her technical input.
Author Disclosure Statement
The authors have no conflicts of interest to declare.
Funding Information
The authors acknowledge the Faculty of Health Sciences Research Committee of the University of the Witwatersrand, Johannesburg and, the National Research foundation of South Africa (Grant number: PR_IFR190125410875).