
Abstract
Supplements containing pharmacological concentrations of biotin are commercially available over the counter. Classical toxicity studies have considered biotin administration as harmless; however, recent investigations have shown that biotin supplementation modifies tissue morphology without changes in toxicity markers, raising concerns about the consequences of morphological changes on tissues' functions and the safety of pharmacological concentrations of the vitamin. Testes are very sensitive to toxicants, and testicular histology is a reliable method to study its function. In this work, we investigated the effects of dietary biotin supplementation on testis morphology and spermatogenesis function using an experimental model, in which we have not observed unfavorable effects on other tissue functions or toxicity markers. Male BALB/cAnNHsd mice were fed a control or a biotin-supplemented diet (1.76 or 97.7 mg biotin/kg diet) for 8 weeks. Compared to the control group, the biotin-supplemented mice presented remarkable testis morphology changes, including increased spermatogonia layers; the cellular mechanism involved is related to increased proliferation. Sperm count and serum testosterone levels were not affected, but spermatozoa motility and morphology were significantly impaired in the biotin-supplemented mice. These results caution against the use of supplements with high concentrations of biotin and indicate that biotin's pharmacological effects on morphology need to be considered in toxicological studies.
Introduction
Biotin is a B-complex vitamin whose physiological role is to act as a covalently bound coenzyme of carboxylases. At present, it is well accepted that pharmacological concentrations of biotin modify biological processes. 1,2 Several reports have documented that pharmacological concentrations of biotin modify the expression of genes involved in glucose and lipid homeostasis, have hypolipemic effects, and decrease hyperglycemia. 1,2 The effects of biotin on these functions indicate its potential to be used in strategies to managing diabetes and dyslipidemia. Indeed, biotin products with pharmacological concentrations of the vitamin are commercially available for different proposes—for example, nail and hair health, pregnancy and breastfeeding, and to reduce blood glucose in people with diabetes. Despite its use in elevated amounts, the highest average level of daily intake that is likely to pose no risk of adverse health to most individuals (tolerable upper intake level of a nutrient [UL]) have not been established. 3,4
Recent studies showing that biotin supplementation modifies tissue architecture without changes in toxicity markers have raised concern about the consequences of morphological changes on tissues' functions and the safety of pharmacological concentrations of biotin. 5 We found that mice fed a biotin-supplemented diet during 8 weeks showed an increased proportion of nucleomegaly and binucleated hepatocytes, altered portal triads with increased dilated sinusoids (27%), increased vascularity (37%), and increased number of bile conducts (19%); these modifications were not associated with increases in classical liver damage indicators and oxidative stress markers, such as malondialdehyde, glutathione, urea, creatinine, and serum liver enzymes. 5 These results indicate that the studies to determine the safeness of pharmacological concentrations of biotin need to take into consideration its effects on tissue morphology.
Other studies support the action of biotin pharmacological concentrations on tissue structure. In animal models of hyperglycemia, biotin supplementation ameliorated the pathological changes in the cellular architecture of the pancreas, 6 kidney, 6 and liver 6,7 in the diabetic animals. In the pancreas of normal mice, we found that 8 weeks of a biotin-supplemented diet increased islet size and changed its typical architecture of alpha-cells at the periphery and beta-cells at the core. 8 In female mice, our studies found that a biotin-supplemented diet decreased both Graafian and ovarian primary follicle numbers. 9 Atrophy of the corpus luteum and ovary stroma were found in rats injected with 50 mg of biotin/kg body weight. 10
The effects of biotin supplementation on the male reproductive system have been poorly investigated. In male rats, Sawamura et al. 11,12 investigated the effects of biotin-supplemented diets on testes, and did not detect changes in testis weight in rats fed a diet containing 100 mg/kg diet or 1000 mg of biotin/kg diet over 6 weeks after weaning. 11 In other studies, 12 the same investigators found that rats fed 400, 800, 1000, and 2000 mg biotin/kg diets for 28 days did not change testis weight, but with 5000 and 8000 mg biotin/kg diets the tissue weight was decreased; unfortunately, they did not analyze testis morphology in these studies.
In rats fed a diet containing 10,000 mg biotin/kg over 6 weeks, the authors found that the weight of the testes decreased by about 75%. Their analysis of testes morphology found that the development of seminiferous tubules was inhibited, and few spermatogonia were observed. Also, the number of mature sperm was markedly lower, and sperm with morphologically abnormal heads, mostly round heads, was increased. However, 10,000 mg biotin/kg diet intake was found to be toxic and caused 50% mortality, 11,12 so it is very likely that the testes changes at this extremely high concentration of biotin resulted from the vitamin's toxic effects at systemic level.
The testes are very sensitive to toxicants, drugs, or radiation, 13,14 and testicular histology is a sensitive and reliable method for studying sperm production. 15 Because the effects of pharmacological concentrations of biotin on tissue morphology raise concern about the consequences for tissue function, in this work, we have investigated the effects of dietary biotin supplementation on testis morphology and function using an experimental model, in which no unfavorable effects on tissue functions or toxicity markers were found. 5,8,9 We hypothesize that this strategy will help to expose possible deleterious effects associated with biotin supplementation and might help to establish biotin's UL.
Materials and Methods
Animal model and experimental design
Animal procedures were done according with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (National Academy of Sciences, Washington, DC, USA, 1996) and approved by the Ethics Committee for Experimentation of the Instituto de InvestigacionesBiomédicas, Universidad NacionalAutónoma de México.
At weaning, 3-week-old male Balb/cAnN mice were fed one of the following diets: biotin-control (TD-97126) or biotin-supplemented diet (TD-02458) containing 1.76 mg and 97.7 mg of free biotin/kg diet, respectively (Harlan Teklad, Madison WI, USA), as described previously. 16 Mice were kept in 12 h light/dark cycles at 22°C ± 2°C and provided water and food ad libitum. After 8 weeks of diet administration, the mice were deprived of food for 12 h and anesthetized with Sevoflurane (Abbott Laboratories, Mexico City, Mexico). The blood, testes, and cauda epidydimal sperm were obtained. Finally, mice were killed by cervical dislocation.
Testosterone measurements
After 8 weeks of diet administration, blood samples were obtained from anesthetized mice between 9 and 10 AM and centrifuged at 10,000 g and 4°C for 10 min. Sera were stored at −20°C until used. Testosterone concentrations were determined by ELISA using a commercial kit (DRG Testosterone ELISA, GmbH, Germany) according to the manufacturer's instructions.
Morphological and morphometric analysis
One testis from each mouse was fixed in formalin. Consecutive 5 μm-thick sections were cut and stained with hematoxylin and eosin. Slides were examined for histological changes under a light microscope (Olympus IX-71 inverted microscope; Olympus Corporation, Tokyo, Japan). Morphology was assessed according to Bacha and Bacha, 17 and included histopathology markers commonly used to quantify testicular injury, which include evaluation of seminiferous tubule diameters, and sloughing. 18
We evaluated a total of about 100 tubules randomly selected from three different sections from eight mice. Tubular spaces were considered as unstained spaces inside the tubule. Alteration of cells which surround the seminiferous tubules in the testis was considered as changes in the epithelium basal lamina. Seminiferous tubule circularity was determined in cross-sections of testes using NIH ImageJ software by the formula 4π (area/perimeter 2 ), a value of 1 indicates the circular shape, and as the value decreases, it denotes elongatation. 19 Tubule elongation degree was determined by measuring the major axis, which goes from one side of the tubule, through the center, to the other side, at the widest part of the tubule. The minor axis is the shortest diameter. 20 Testes structures appearing with a frequency minor to 5% were classified as sporadic anomalies.
The seminiferous degree of maturity was determined by the Jonhsen index, 21 assigning a score to each tubule according to the main cell-type present, ranging from 1 (no cells) to 10 (complete spermatogenesis with more than 10 late spermatids). The number of interstitial Leydig cells was determined as reported. 22 Sertoli cells were identified by the presence of two nucleoli. The analyses were performed by two persons who were blinded to the group identity. All images were analyzed using ImageJ 1.50 software (National Institute of Mental Health, Bethesda, MD, USA).
Immunofluorescence studies
Proliferation and apoptosis were analyzed by immunofluorescence as described previously. 23 Cell proliferation was assessed with anti-rabbit Ki67 (Abcam #15580, Cambridge, MA, USA). After 1.5 h at 37°C of incubation, slides were washed and incubated for 2 h at room temperature with AlexaFluor-594-conjugated secondary antibody (Abcam #150080). Nuclei were stained with DAPI (Sigma #D9542, St. Louis, MO, USA). As a negative control, the primary antibody was substituted with phosphate-buffered saline (PBS). Seminiferous tubule cell proliferation was measured by quantifying the number of Ki67-positive nuclei in relationship to the number of total nuclei. A total of 127 tubules were counted from each group using ImageJ 1.50 software.
Apoptosis was analyzed using the TUNEL Assay Kit (In situ Cell Dead Detection Kit, fluorescence; Roche Diagnostics #11684795910, Manheim, Germany). Nuclei were counterstained with DAPI, and apoptosis was analyzed by quantifying the number of TUNEL-positive nuclei in relationship to the number of total nuclei. A total of 155 tubules from both groups were counted using ImageJ 1.50 software.
Western blot analysis
The tissue was homogenized and total protein quantified as described previously. 24 Next, 35 μg of protein were electrophoresed on preparative SDS polyacrylamide gel. The proteins were transferred to a nitrocellulose membrane and probed with antibodies against Ki67, caspase-3 (Cell Signaling Technology #96615; Danvers, MA, USA), and glyceraldehyde-3 phosphate dehydrogenase (GAPDH; Cell Signaling Technology #88845). Anti-IgG-HRP conjugates were used as secondary antibodies (Cell Signaling Technology #70765 and 7074). Protein bands were detected in a Molecular Imager ChemiDoc XRS System (Bio-Rad, Hercules, CA, USA) and quantified by densitometry using ImageJ 1.40 (Research Services Branch, National Institute of Mental Health).
Spermatozoa isolation
Both epididymides were obtained, placed on a prewarmed culture dish, maintained at 37°C. Then, fat and connective tissues were removed. 25 Caudal regions from the epididymis were isolated and minced. Sperm cells were allowed to disperse out of the cauda. Afterward, sperm in suspension were washed in PBS by centrifugation at 600 g for 5 min. Finally, sperm cells were counted using a hematocytometer.
Sperm morphology and motility
Spermatozoa were fixed in formaldehyde and counted as previously described. 26 To evaluate flagellar alterations, 1000 spermatozoa from each mouse (n = 8) strain were classified and counted. Sperm morphology was examined by phase-contrast microscopy (Olympus IX-71 inverted microscope; Olympus Corporation) and classified as follows: (1) with normal flagella, not curved or twisted or (2) with any number of all curved flagella in the middle piece. 26 Sperm motility was analyzed using a light microscope and counting motile and nonmotile spermatozoa and was expressed as motility percent. 27
Statistical analysis
All data are presented as the mean ± SEM; n denotes the number of evaluated subjects. Statistical analysis was performed using GraphPad (La Jolla, CA, USA). The data were analyzed by Student's t-test. P ≤ .05 was considered statistically significant.
Results
Effect of biotin supplementation on body and testis weight and serum testosterone concentration
We determined the effect of 8 weeks of biotin supplementation on body and testis weight and hormonal concentration. As shown in Table 1, no differences were observed regarding the body and testis weight, the testis weight to body weight ratio, or the serum testosterone levels between the groups.
Effects of Biotin on Body and Testis Weight, Testis Weight/Body Weight Ratio, and Serum Testosterone Concentration
Values are means ± SEM n = 8 mice for group.
Effects of biotin supplementation on testis morphology
No differences between the control and the biotin-supplemented group were observed in the external appearance of the testes (Fig. 1A). Light microscopy of the testes revealed a number of differences between the biotin-supplemented group and the control group (Fig. 1B–D). As seen in Figure 1C, the testis of supplemented mice presented an increased percent of nonvisible seminiferous tubule lumen (control: 22.5% ± 0.94%; biotin-supplemented: 29.4% ± 2.90%; P < .05) and in the number of seminiferous tubules with intratubular spaces (control 7.90% ± 2.70%; biotin-supplemented 24.0% ± 3.80%; P < .005). Sloughing of round germs in the lumen was observed. The testis interstitial space showed an increase in Leydig cells (control: 16.0% ± 0.42%; biotin-supplemented: 19.7% ± 0.67%; P < .0001). Furthermore, the supplemented group (Fig. 1D) presented loss of circularity, increased elongated seminiferous tubules (control: 13.8% ± 2.37%; biotin-supplemented: 49.3% ± 8.14%; P < .0001) with cellular disorganization, more than three spermatogonia layers, and alteration in the germinal epithelium (control: 0.38% ± 0.18% biotin supplemented: 3.50% ± 0.32%; P < .0001). We also found a decrease in the degree of maturity determined by the Jonhsen Index (control: 9.03 ± 0.10; biotin-supplemented: 8.42 ± 0.09; P < .0001). No difference between the groups was observed in the number of Sertoli cells (control: 12.4 ± 0.18; biotin-supplemented: 12.5 ± 0.18).
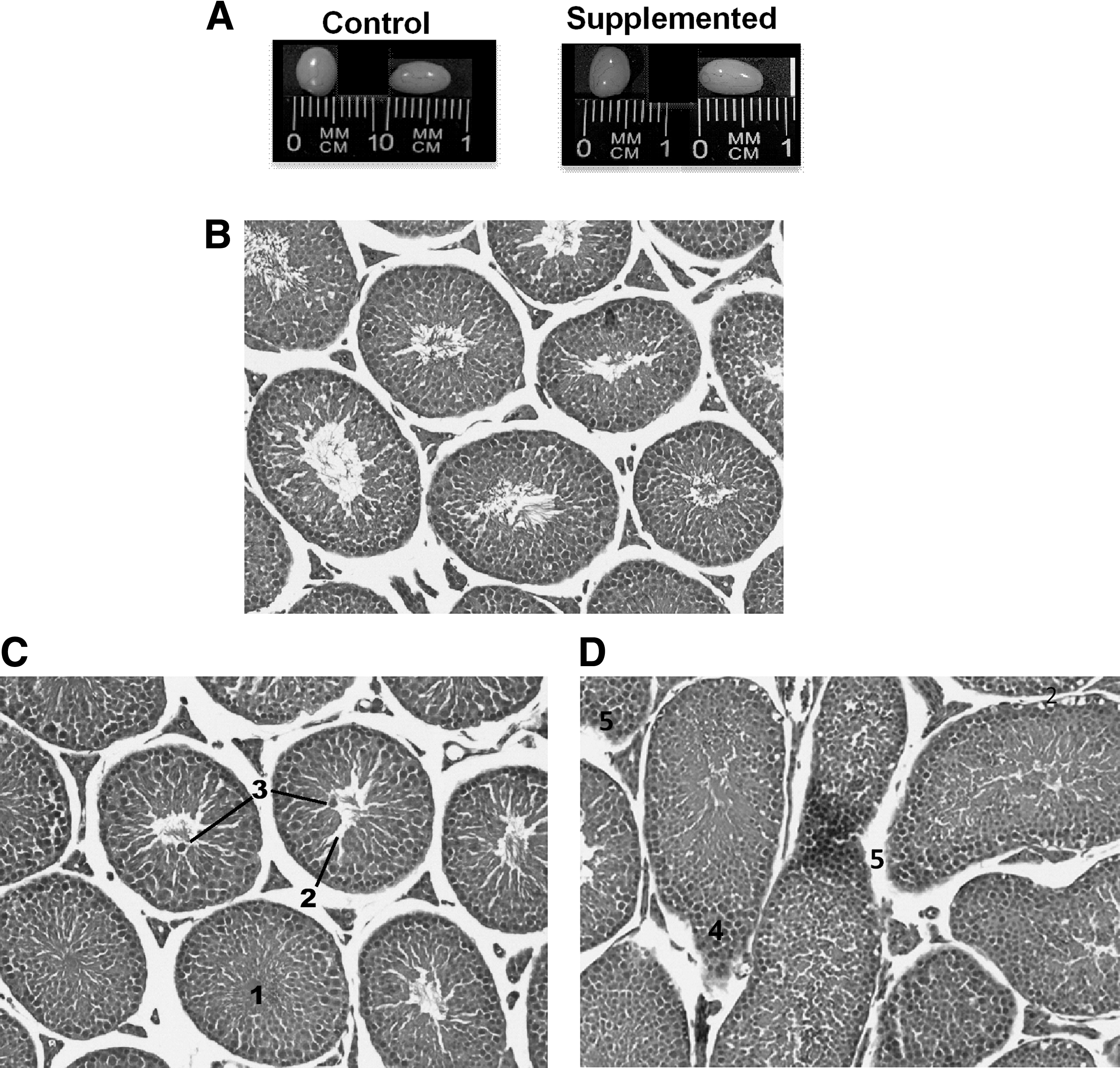
Effect of biotin supplementation on testis size and morphology.
Testes structures appearing with a frequency minor to 5% in the biotin-supplemented mice included sloughing (2.0% ± 0.21%) (Fig. 2A, B), incomplete spermatogenesis (1.97% ± 0.33%) (Fig. 2C), intratubular spaces in the elongated tubule (1.42% ± 0.33%) (Fig. 2C), and Leydig cell hyperplasia (1.5% ± 0.32%) (Fig. 2 D). None of these structures was found in the control group.

Effect of biotin supplementation on testis producing sporadic anomalies.
Effect of biotin supplementation on sectional seminiferous tubule circularity
We quantified the loss of seminiferous tubule circularity. Compared to the control group, the biotin-supplemented mice presented decreased circularity (Fig. 3A) (control: 0.93 ± 0.006; biotin-supplemented: 0.78 ± 0.017; P < .0001). Also, we assessed the degree of tubule elongation by measuring the long and short diameter of the seminiferous tubules (Fig. 3B). The long diameter of the seminiferous tubules was higher in the biotin-supplemented mice than in the control group (control: 231 ± 4.29 μm; biotin-supplemented: 353 ± 16.7 μm; P < .0001). No significant statistical difference between the groups was observed regarding the short diameter (control: 173 ± 1.41 μm; biotin-supplemented: 185 ± 4.29 μm). These data indicate that the shape changes produced by biotin supplementation were due to elongation rather than to a decrease in the tubule diameter.

Effect of biotin supplementation on seminiferous tubule circularity and diameters.
Effect of biotin supplementation on cell proliferation of testes
We further explored the mechanisms involved in the effects of biotin supplementation on testes. The effect of biotin on cell proliferation was determined by measuring the nuclear expression of Ki67. As shown in Figure 4A, immunofluorescence studies showed that the Ki67 label was increased in the spermatogonia (Fig. 4A). Quantification of spermatogonia immunofluorescent positive nuclei showed an increase in the biotin-supplemented mice (control: 20.3% ± 0.95%; biotin-supplemented: 27.6% ± 1.05%; P < .0001) (Fig. 4B). The Ki67 label was also present in the nuclei of Leydig cells (Control: 0.50% ± 0.19%; biotin-supplemented: 0.44% ± 0.18%; P > .05), as well as in the cytoplasm. We also analyzed Ki67 protein abundance by western blot (Fig. 4C). Accordingly with the augment observed in the immunofluorescence studies, biotin supplementation increased Ki67 protein expression (Fig. 4D) (control: 0.19 ± 0.016; biotin-supplemented: 0.94 ± 0.047; P < .001).
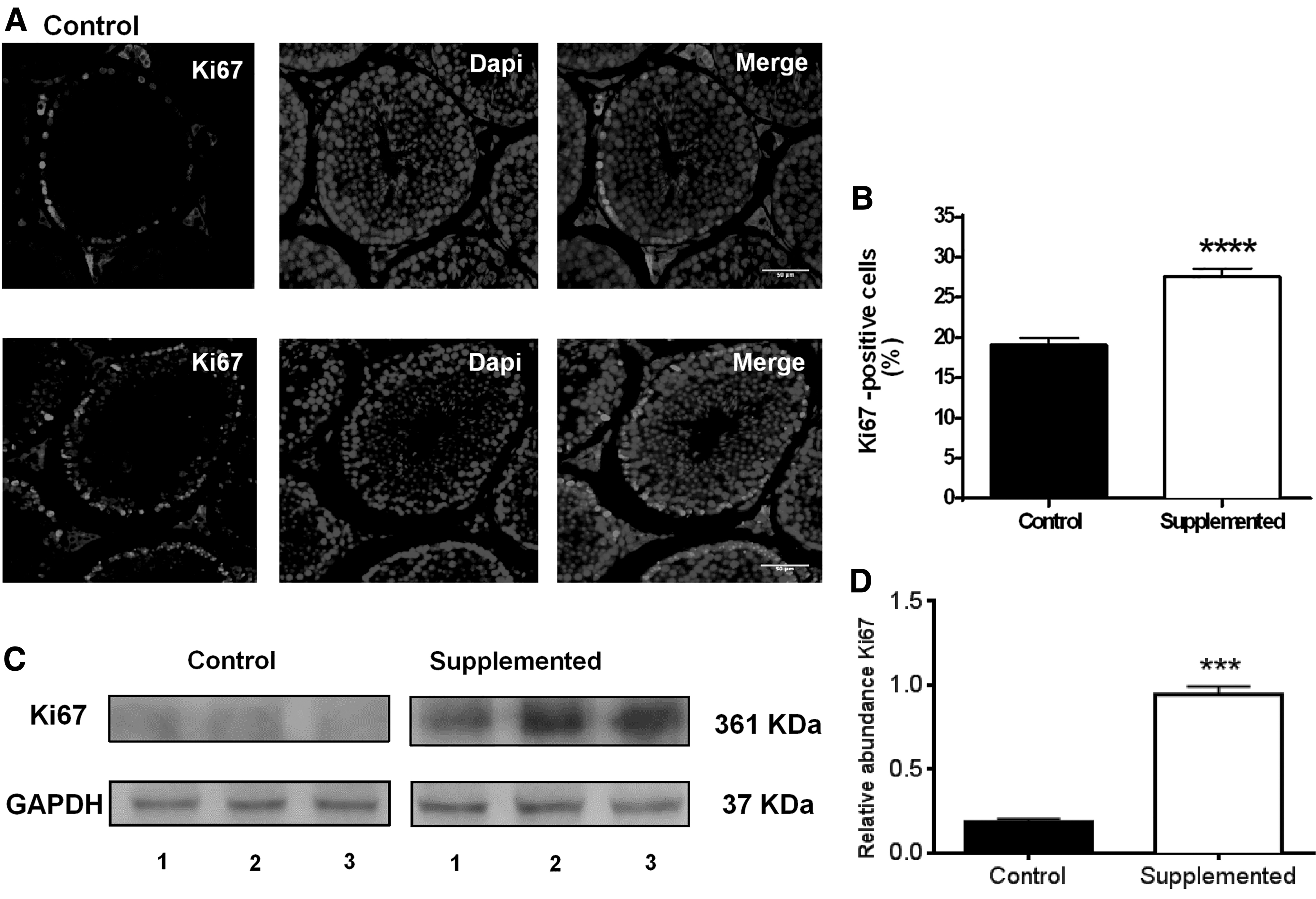
Effect of biotin supplementation on seminiferous tubule proliferation.
Effects of biotin supplementation on testis apoptosis
Immunofluorescence studies revealed that the percentage of TUNEL-positive nuclei was not significantly different between the control and the supplemented group (Fig. 5A, B) (control: 0.40% ± 0.057%; biotin-supplemented 0.35% ± 0.049%). We also determined the expression of the apoptosis protein caspase-3 (Fig. 5C, D). The data showed that the protein abundance of the active form of caspase (caspase-3) was increased in the biotin-supplemented group (control: 0.25 ± 0.030; biotin-supplemented: 0.68 ± 0.12; P < .005). No significant difference was observed in the caspase inactive form (procaspase) protein levels (control: 0.67 ± 0.084; biotin-supplemented: 0.84 ± 0.10).
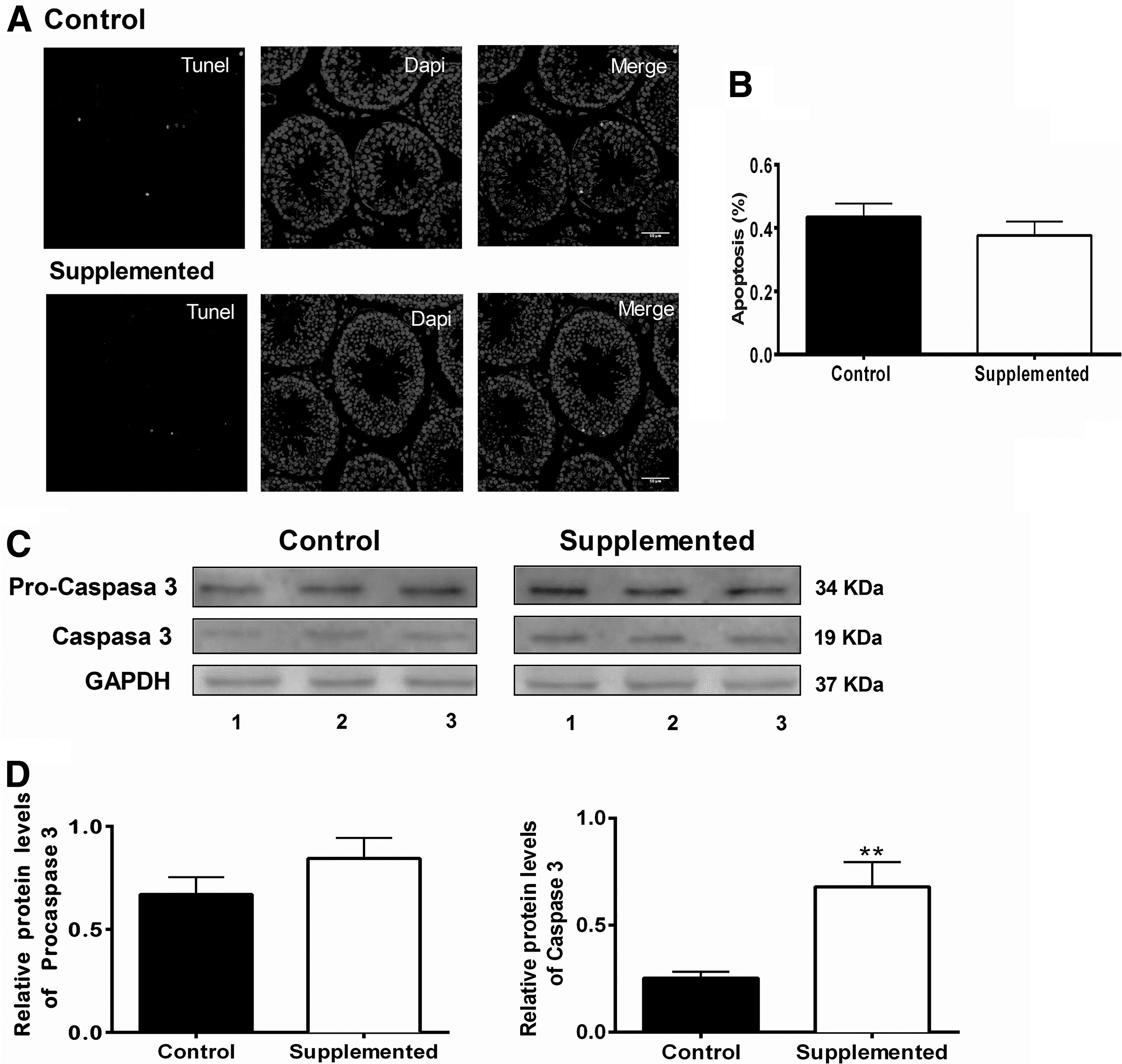
Effect of biotin supplementation on seminiferous tubule apoptosis.
Effects of biotin supplementation on sperm number and quality
The analysis of the effects of biotin supplementation on spermatozoa number found no significant differences in the cauda sperm count between groups (Fig. 6A). The sperm motility of the mice fed the biotin-supplemented diet was significantly decreased compared to the control group (Fig. 6B). Furthermore, sperm from the biotin-supplemented group showed altered morphology (Fig. 6C) with a significant increase in the number of spermatozoa with twisted flagella in the middle piece (control: 6.50 ± 0.68; biotin-supplemented: 30.4 ± 2.04; P < .0001) (Fig. 6D).
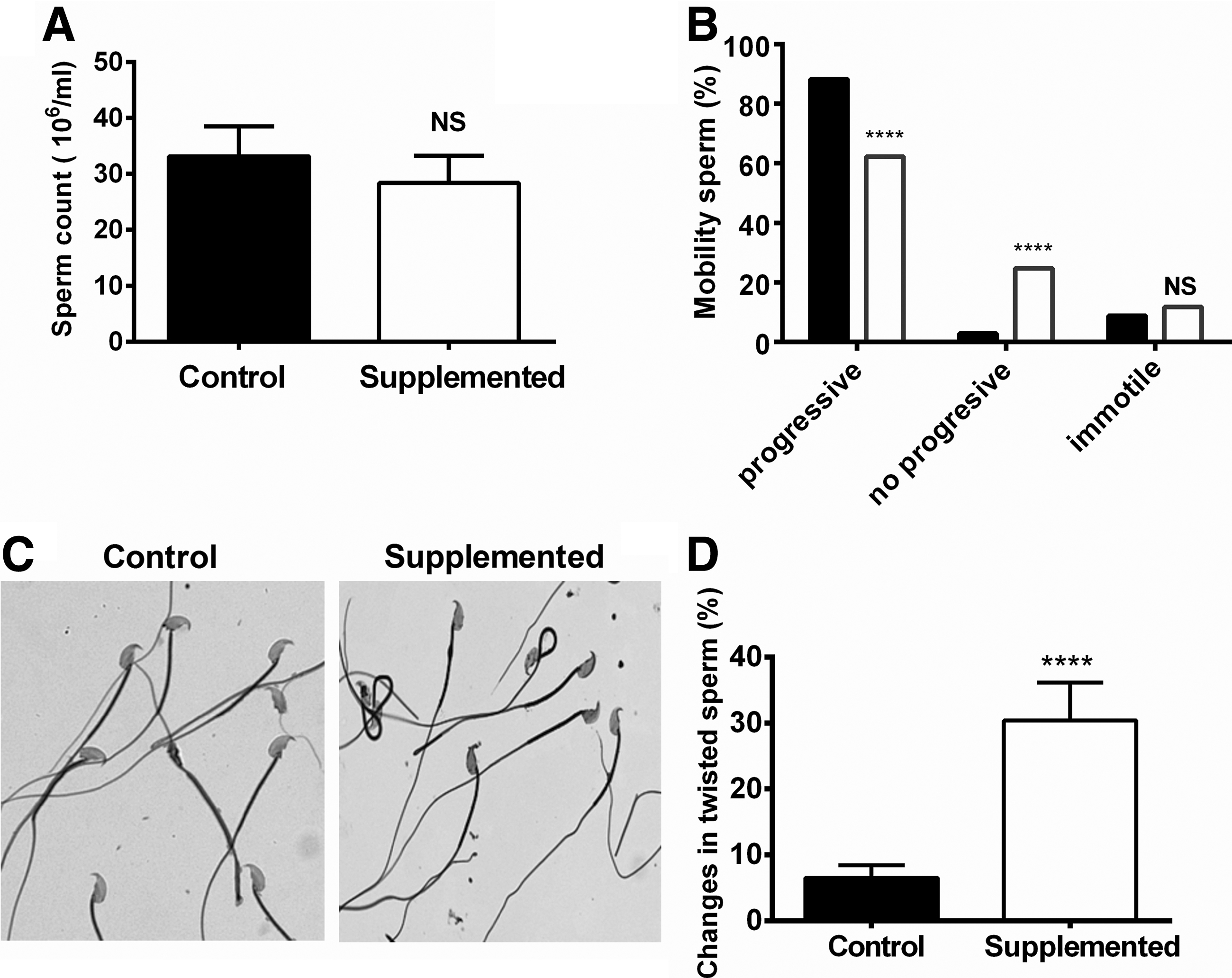
Effect of biotin supplementation on spermatozoa.
Discussion
Biotin administration is considered harmless. 28 However, recent studies showing that biotin supplementation affects tissue morphology without modifying toxicity markers have raised concern about the functional consequences of these changes and the safety of pharmacological concentrations of biotin.
Our present results showed that biotin supplementation produced striking effects on the histology of mouse testis and also affects the sperm quality. Compared to the control group, the testes of supplemented mice presented increases in (1) the number of seminiferous tubules with nonvisible lumen; (2) intratubular spaces; (3) sloughing of round germs in the tubular lumen; and (4) increased Leydig cell accumulation. Also, the supplemented group presented alteration in the germinal epithelium and elongated seminiferous tubules with cellular disorganization as well as increased spermatogonia layers. Investigation of the mechanisms involved in these changes revealed increased spermatogonia proliferation. The increased proliferation without augmented spermatozoa number and reduced Jonhsen index suggest delayed spermatogenesis.
The changes in the morphology of the seminiferous tubules resulted in defective spermatozoa formation and impaired sperm motility. Some features observed in the biotin-supplemented mice, such as Leydig cell hyperplasia or tubule elongation are produced by other toxics, pathologies 29 or by transgenesis of the plus end-tracking protein EB1, 19 but the ensemble of biotin-induced changes appear to be sui generis; however, we cannot rule out the possibility that these changes might lead to other well-defined degenerative process.
Sawamura et al. 11,12 studied the effects of 6 weeks of biotin-supplemented diets on the morphology rat testes. As in our present investigation, they observed no changes in testis weight, food intake, and body weight gain when comparing a diet containing a biotin concentration and the diet used in this work (about 100 mg/kg diet); unfortunately, they did not perform morphological analysis in these animals. In rats fed a biotin-supplemented diet with the 10,000 mg biotin/kg diet, their investigations found that the weight of the testes decreased by about 75%. Histological analysis showed a severe decrease in the total sperm count (less than 1 sperm/mL), increased incidence of sperm with abnormal morphology, mainly round heads, decreased spermatogonia, decreased diameters of seminiferous tubules, and did not showed changes in tubule circularity. No changes were observed in testicular testosterone. In contrast, in our studies in mice fed a biotin-supplemented diet containing 97.7 mg biotin/kg diet over 8 weeks, we found no significant differences in the total sperm count, sperm abnormalities were observed in the flagellum, and the tubule area and the number of spermatogonia layers were increased. Serum testosterone levels were not significantly different when comparing the control and supplemented groups.
It is important to note that the data reported by Sawamura et al. 11 are difficult to interpret since the same report showed that rats fed with a diet containing 10,000 mg biotin/kg diet also presented significant decreases in food intake as well as in body, liver, kidney, and brain weight. Indeed, this magnitude of biotin was shown to be toxic in other studies. 12 In contrast, in the conditions of our work (97.7 mg biotin/kg diet during 8 weeks), a model in which unfavorable effects have not been observed on body weight gain, tissues weight, food consumption, external mice appearance, or toxicity markers, 5,8,9 we found increased incidence of sperm with abnormal morphology and structural changes of the testes, indicating that the effects of biotin supplementation observed on testes in our experimental model was not due to systemic toxicity.
In prior investigations, we found that normal mice fed the biotin-supplemented diet used in this study presented morphological variations in different organs but did not show function alterations. 5,8,9 The cell topology changes produced in the pancreatic islets did not negatively affect their hormone secretion. 8 In the liver, despite the structural changes, hepatic functions such as gluconeogenesis and glycogenesis were not different from those presented in the control mice. 30 Furthermore, hepatic damage enzymes indicators or oxidative stress markers were not modified. 5 In contrast, in the testes, biotin supplementation, in addition to modifying the tissue morphology, affected their function of sperm quality production. Since testes are very sensitive to toxics 13,14,31 and they have high capacity of biotin accumulation, 11 testes are likely to be one of the tissues more susceptible to pharmacological concentrations of biotin, as demonstrated in the present work.
Proliferation and apoptosis are cellular mechanisms that participate in the normal mammalian testes during spermatogenesis. 32 Proliferation is required to augment cell production, and appropriated apoptosis is a regular cellular mechanism in the normal mammalian testes during the development of spermatogonia and is used to remove excess germ cells. Our results demonstrate that biotin supplementation significantly increases Ki67 in total cellular extracts of testes, which results from the presence of the protein in the spermatogonia and Leydig cells, as shown in the immunofluorescent studies. No significant differences were observed in TUNEL staining. These data suggest that the mechanism involved in enhanced cell layer number is mainly due to cell proliferation. In support of the effects of biotin supplementation on cell proliferation, previous investigations in our laboratory have also found that pharmacological concentrations of biotin increases cell proliferation but not apoptosis. 23
Biotin supplementation induced Leydig cell hyperplasia; however, at 8 weeks of biotin supplementation, Ki67 labeling was mainly present in the cytoplasm. Leydig cell proliferation is achieved between postnatal days 21 and 35, and they gradually mature into fully steroidogenic adult cells by postnatal day 56. 33,34 Because we initiated biotin-supplementation at 3 weeks of age, the results suggest that Leydig cell hyperplasia was produced between the first and third week of experimentation (3–5 weeks of age), during the time that these cells proliferate.
The changes produced on Leydig cell hyperplasia were not translated into increases in serum testosterone levels. Studies by Sawuamura 11 found that testicular testosterone was not affected by a diet containing 10,000 mg biotin/kg. Further studies will be required to determine the effect of biotin on male steroidogenesis because, in female mice, we found that the biotin-supplemented diet used in the present study increased serum estradiol levels. 9
It is noteworthy that we found a discrepancy between the increase in the expression of caspase-3 and no difference in TUNEL immunoreactivity. Apoptosis is characterized by nuclear chromatin condensation and DNA fragmentation caused by apoptotic signaling cascades. Apoptosis signaling cascades are mediated by caspases, which trigger cell death by cleaving specific proteins in the cytoplasm and nucleus. Cleaved caspase-3 is an apoptosis effector that translocates to the nucleus, where it cleaves substrates that induce DNA fragmentation. Once activated, caspase-3 might be subject to inhibition by apoptosis protein inhibitors (IAP protein families). 35 Because the TUNEL assay identifies DNA fragmentation, which is the end-stage apoptosis, and caspase-3 is an undergoing apoptosis effector, it is possible that an increased length of a biotin-supplemented diet and/or higher biotin content could translate to increased TUNEL positive cells. However, we cannot rule out the possibility that active caspase-3 might be subject to downstream regulation by apoptosis protein inhibitors. 35 Studies in our laboratory are addressing this issue.
Studies regarding human spermatozoa by Kalthur et al. 36 found that in vitro, biotin supplementation 2.44 mg/mL (10 nM) augmented motility and prolonged the survival of semen samples. Furthermore, in a recent report, 37 the same group reported that this concentration of biotin increases the fertilizing ability of mice spermatozoa and subsequent mice preimplantation embryo development, suggesting that the vitamin may have benefits in assisted reproduction. In contrast, our present studies show that in vivo a biotin-supplemented diet decreased spermatozoa motility. The dissimilar effect of biotin in these studies versus our investigation might be related to the concentration of biotin. The serum concentrations of biotin in mice fed with the present diet for 8 weeks are 144.1 ± 1.59 mg/mL (590 ± 6.5 nM), 8 about 60-fold the amount present in the media of Kalthur studies. However, we cannot rule out that the different study conditions and/or the different sensitivity to biotin between species, as demonstrated in other studies, 38 might account for the discrepancy.
The use of vitamin supplements in larger amounts has escalated considerably in recent years. Water-soluble vitamins are considered to be eliminated and to be safe at higher doses, it is clear that there is a need to assess the risk of vitamins in the context of the wide availability of supplements. Commercially available biotin supplements contain up to 10 mg of the vitamin. In humans, supplementation of 1.2 mg biotin/day for 14 days results in serum concentrations in a range of 9.4–47.7 nM 39 (2.29–11.6 mg/mL), so it is likely that the serum biotin levels attained by supplements with high levels of biotin could reach the vitamin concentration in the same order of magnitude observed in our studies. 8
Classical toxicity tests 40,41 and other studies 2 have suggested that biotin administration is harmless 28 ; however, taking into account the effects of biotin on testes function and morphology obtained in this work, and that biotin supplements with high levels of biotin could attain the vitamin concentrations observed in our studies, 8 human pharmacokinetic and pharmacodynamic studies are imperative given that biotin's UL has not been established. 3,4
In conclusion, the results of this work showed that studies meant to determine the safety of pharmacological concentrations of biotin need to consider its effects on tissue morphology and function.
Footnotes
Acknowledgments
The authors are grateful to PhD Candidate Esmeralda Enríquez Calderón and BS Armando Elizalde Gualito for technical assistance, and Sandra Delfín Azuara for technical and English assistance.
Author Disclosure Statement
No competing financial interest exists.
Funding Information
This work was supported by funds from Consejo Nacional de Ciencia y Tecnología CONACyT 219787 (C.F.M.), Fondos Federales 074/2013 (C.F.M.), and CONACyT 284183 (E.O.H.G.).