
Abstract
Excess of visceral adipose tissue (VAT) characteristic of obesity leads to a proinflammatory state disrupting the insulin signaling pathway, triggering insulin resistance (IR) and inflammation, the main processes contributing to obesity comorbidities. Ursolic acid (UA), a pentacyclic triterpenoid occurring in a variety of plant foods, exhibits anti-inflammatory properties. The aim of this study was to evaluate UA effects on IR, hyperinsulinemia, and inflammation in experimental diet-induced obesity. Forty male Wistar rats were randomly assigned to eight groups (n = 5). One group was used for time 0. Three groups were labeled as OBE (control): receiving high-fat diet (HFD; fat content 45.24% of energy) during 3, 6, or 9 weeks; three groups UA-PREV: exposed to simultaneous HFD and UA during 3, 6, or 9 weeks to evaluate UA preventive effects; one group UA-REV: receiving HFD for 6 weeks, followed by simultaneous HFD and UA for three additional weeks to analyze UA reversal effects. Measurements were performed after 3, 6, or 9 weeks of treatment. Adiposity was calculated by weighing VAT after sacrifice. Serum markers were quantified through colorimetric and enzyme-linked immunosorbent assay methods. VAT adipokines RNAm expression was evaluated by quantitative reverse transcriptase–polymerase chain reaction. Data were analyzed by Kruskal–Wallis and Mann–Whitney U tests. UA significantly decreased adiposity, IR, hyperinsulinemia, triacylglycerides, and cholesterol levels, and also VAT mRNA expression of MCP-1 (monocyte chemoattractant protein-1), IL (interleukin)-1β and IL-6, concomitantly increasing adiponectin levels. UA metabolic effects demonstrated in this study support its potential therapeutic utility to improve IR, hyperinsulinemia, and inflammation observed in obesity and diabetes.
Introduction
Obesity represents a major health problem worldwide characterized by excessive adipose tissue, 1,2 subclinical chronic inflammation, hyperinsulinemia, and insulin resistance (IR). 3,4 IR, a condition impeding an appropriate response to circulating insulin, constitutes a major risk factor for type 2 diabetes mellitus (T2D) and other obesity comorbidities. 5 Visceral adipose tissue (VAT) accumulation in the peritoneal cavity positively disrupts the proinflammatory/anti-inflammatory balance of adipokines gene expression promoting hyperinsulinemia and IR. This proinflammatory profile is evidenced by monocyte chemoattractant protein-1 (MCP-1), interleukin 6 (IL-6), IL-1β, and tumor necrosis factor α (TNF-α) increase, simultaneously with adiponectin and anti-inflammatory adipokines diminution, among other molecules. 6 –10 MCP-1 induces macrophage recruitment increasing the proinflammatory gene expression and impairing insulin signaling 11,12 ; IL-1β leads to insulin signaling blockage and progressive loss of the insulin-producing pancreatic β cell 13 ; IL-6 and TNF-α contribute to insulin signaling pathway disruption, and are negatively associated with adiponectin expression, an insulin-sensitizing adipokine. 14 –16
Hyperinsulinemia constitutes an underlying factor of obesity-related comorbidities and its role in initiating, sustaining, and expanding IR, a T2D hallmark, is relevant. 5,17 Furthermore, IR is associated with metabolic and cardiovascular disorders, such as dyslipidemia, hypertension, obesity, glucose intolerance, and endothelial dysfunction. 18 IR and hyperinsulinemia coincide with an increased cancer risk in a variety of conditions, including abdominal obesity, T2D, hyperglycemia, and subclinical inflammation. 19
Ursolic acid (UA) is a lipophilic pentacyclic triterpenoid carboxilic acid widely distributed as a component of diverse fruits and herbs traditionally used in folkloric medicine, originally discovered in Arctostaphylos uva-ursi leaves more than 150 years ago, as mentioned by Sando. 20 This triterpenoid is a rosemary component and contributes to the waxy coats of apples, pears, prunes, and other fruits. 21 –25 UA highest content has been reported in Rosmarinus officinalis (2.95 g/100 g dried matter). 24 Antiangiogenesis, 26 antitumoral, antioxidant, 27,28 anti-inflammatory activity, 29 –31 and other biological properties have been attributed to UA. 26 –28,32 UA targets multiple proinflammatory transcription factors, kinases, proinflammatory cytokines and enzymes, chemokines, and adhesion molecules promoting its anti-inflammatory action. 21,33 As inflammation is considered one of the pivotal causes of IR and hyperinsulinemia in obesity, 6 –15 the aim of this study was to assess whether UA may improve proinflammatory adipokines expression, IR, and hyperinsulinemia in rats with diet-induced obesity.
Materials and Methods
UA, high-fat diet, and food intake
UA, purchased from Indofine Chemical Company, Inc. (Hillsborough, NJ, USA), 98% purity, was dissolved in dimethyl sulfoxide 0.05%-polyethylene glycol 400 0.1%-distilled water and orally administered (50 mg/kg/day p.o.). 29 High-fat diet (HFD) was weekly prepared in accordance with the American Institute of Nutrition recommendations, 34 fat content 45.24% of energy (lard was added), 35 protein 16.63%, and carbohydrates 38.13%, providing 493.2 kcal/100 g. Ingredients were mixed until obtaining a homogeneous powder, analyzed by bromatological studies (Table 1) and stored at 4–8°C. Food intake was calculated once a week every 3 weeks by subtracting the remaining food from the fresh portion served.
High-Fat Diet Composition
MUFA, monounsaturated fatty acid; PUFA, polyunsaturated fatty acid; SFA, saturated fatty acid.
Experimental animals
Forty 7-week-old male Wistar rats, 182.5 ± 11.3 g, exposed to 12 h:12 h light/dark cycles, 22°C, constant humidity, and ad libitum access to water/food were randomly assigned to eight groups (n = 5). Initial measurements of one group were used for time 0. Based on metabolic changes occurring in experimental obesity models under HFD supplementation, 36 experiment was carried out for 9 weeks divided into three periods. Three groups were labeled as OBE: animals receiving HFD during 3, 6, or 9 weeks, used as controls; three groups labeled as UA-PREV: exposed to simultaneous HFD and UA supplementation during 3, 6, or 9 weeks to evaluate UA preventive effects; and one group labeled UA-REV: receiving HFD during 6 weeks, followed by simultaneous exposure to HFD and UA for three additional weeks to analyze UA reversal effects.
The guidelines of University of Guadalajara Ethics Committee, based on the technical specifications for the production, care, and use of laboratory animals, according to NOM-062-ZOO-1999, 37 and the International Guiding Principles for Biomedical Research Involving Animals established in 1985, were followed. 38
Sample collection
Animals fasted 12 h were anesthetized with 30 mg/kg i.p. Zoletil® Virbac (Carros, Cedex) after 0, 3, 6, or 9 weeks of treatment. Retro-orbital plexus blood (3 mL) was collected and centrifuged to determine metabolic and inflammatory serum markers. Before sacrifice, ∼30 mg of mesenteric VAT were obtained for adipokines mRNA expression quantification. The remaining VAT (mesenteric, retroperitoneal, epididymal, and perirenal fat depots) was collected after sacrifice and reported as adiposity. Serum and VAT samples were stored at −80°C.
Biochemical determinations
Fasting circulating cholesterol, and triacylglycerides were analyzed using commercial kits (Boehringer, Mannheim, Germany) involving a spectrophotometric wet chemistry method. Fasting blood glucose was determined by a pyrroloquinoline quinone-dependent glucose dehydrogenase enzyme method using an amperometric glucose sensor (Accu-Chek Performa Roche glucometer; Mannheim, Germany).
Insulin, IR, adiponectin and adipokines determination
Circulating insulin, adiponectin, MCP-1 and TNF-α were quantified by enzyme-linked immunosorbent assay (ELISA) kits from ALPCO Diagnostics (Salem, NH, USA), IL-1β and IL-6 by R&D ELISA kits (Minneapolis, MN, USA). These measurements were made by duplicate utilizing an iMark® Microplate Absorbance Reader from Bio-Rad (Hercules, CA, USA). To determine IR, the homeostasis model assessment of insulin resistance (HOMA-IR) was calculated as fasting glucose concentration (mmol/L) × fasting insulin concentration (μUI/mL)/22.5. 39
RNA isolation and synthesis of complementary DNA
Total RNA was isolated from mesenteric VAT sample according to the Chomczynski and Sacchi method 40 using a polytron homogenizer and a purification silica-based system (Invitrogen, Carlsbad, CA, USA). RNA quantification was performed in a Nanodrop 2000 Thermo Scientific spectrophotometer (Waltham, MA, USA) and stored at −80°C. Reverse transcription was performed using 2 μg RNA in a 0.05 M Tris-HCl pH 8.3 buffer containing 0.05 μg/μL random hexamers, 1 mM dNTP mix, 0.05 U/mL RNAse inhibitor, and 200 U/mL M-MLV reverse transcriptase. Samples were incubated for 10 min at 70°C, then for 60 min at 37°C. Finally, reverse transcriptase was inactivated by incubation for 10 min at 95°C, storing the complementary DNA (cDNA) at −20°C. 12
Analysis of proinflammatory gene expression
MCP-1, IL-1β, IL-6, and TNF-α gene expressions were analyzed by quantitative reverse transcriptase–polymerase chain reaction in a real-time thermocycler (Corbett Life Science Rotor Gene® 6000 system; Concorde, NSW, Australia), using TaqMan probes (Applied Biosystems, Foster City, CA, USA). Reactions were performed in a total volume of 10 μL containing 5 μL TaqMan® Master Mix (Invitrogen), 0.5 μL specific TaqMan probe (Applied Biosystems), 1 μL (100 ng) cDNA, and 3.5 μL water. Samples were heated at 50°C for 2 min and 94°C for 10 min, followed by 45 cycles at 94°C for 15 sec and 60°C for 40 sec. Gene amplification was normalized against β-actin expression. Samples were analyzed by triplicate and mean values were calculated. Relative quantification by the 2−ΔΔCT method was accomplished considering the control group as an internal calibrator. 41
Statistical analysis
Data were analyzed with the Statistical Package for the Social Sciences version 12. Variables were expressed as the mean ± standard deviation and analyzed by the Kruskal–Wallis test owing to the small sample size. Differences among groups were assessed by post hoc Mann–Whitney U test and P < .05 was considered statistically significant. 42
Results
UA prevents hyperinsulinemia, improves HOMA-IR and adiponectin levels
Fasting circulating glucose, insulin, and adiponectin levels were quantified. Glucose levels ranged between normal values (70.8 ± 4.9 to 84.2 ± 18.3 mg/dL) (Table 2). UA prevented hyperinsulinemia in animals receiving simultaneously HFD and UA, as statistically significant (P < .05) lower insulin levels were observed in UA-PREV animals versus OBE after 6 (26.1 ± 12.9 μUI/mL vs. 82.0 ± 9.6 μUI/mL) and 9 weeks of treatment (22.3 ± 2.8 μUI/mL vs. 60.7 ± 7.6 μUI/mL). Similarly, significant hyperinsulinemia reduction was found in UA-REV group compared with the OBE group (34.8 ± 10.7 μUI/mL vs. 60.7 ± 7.6 μUI/mL) (Fig. 1a). Consequently, HOMA-IR statistically diminished in UA-PREV animals compared with OBE after 6 (5.5 ± 2.7 vs. 16.5 ± 1.9) and 9 weeks of treatment (4.1 ± 0.8 vs. 10.8 ± 1.8), whereas a lowering tendency was seen in UA-REV group (Fig. 1b). Likewise, statistically higher (P < .05) levels of adiponectin were observed in UA-PREV group after 9 weeks of treatment (12.8 ± 0.3 μg/mL vs. 11.1 ± 0.4 μg/mL) and UA-REV group compared with the OBE group (14.0 ± 1.5 μg/mL vs. 11.1 ± 0.4 μg/mL) (Fig. 1c).
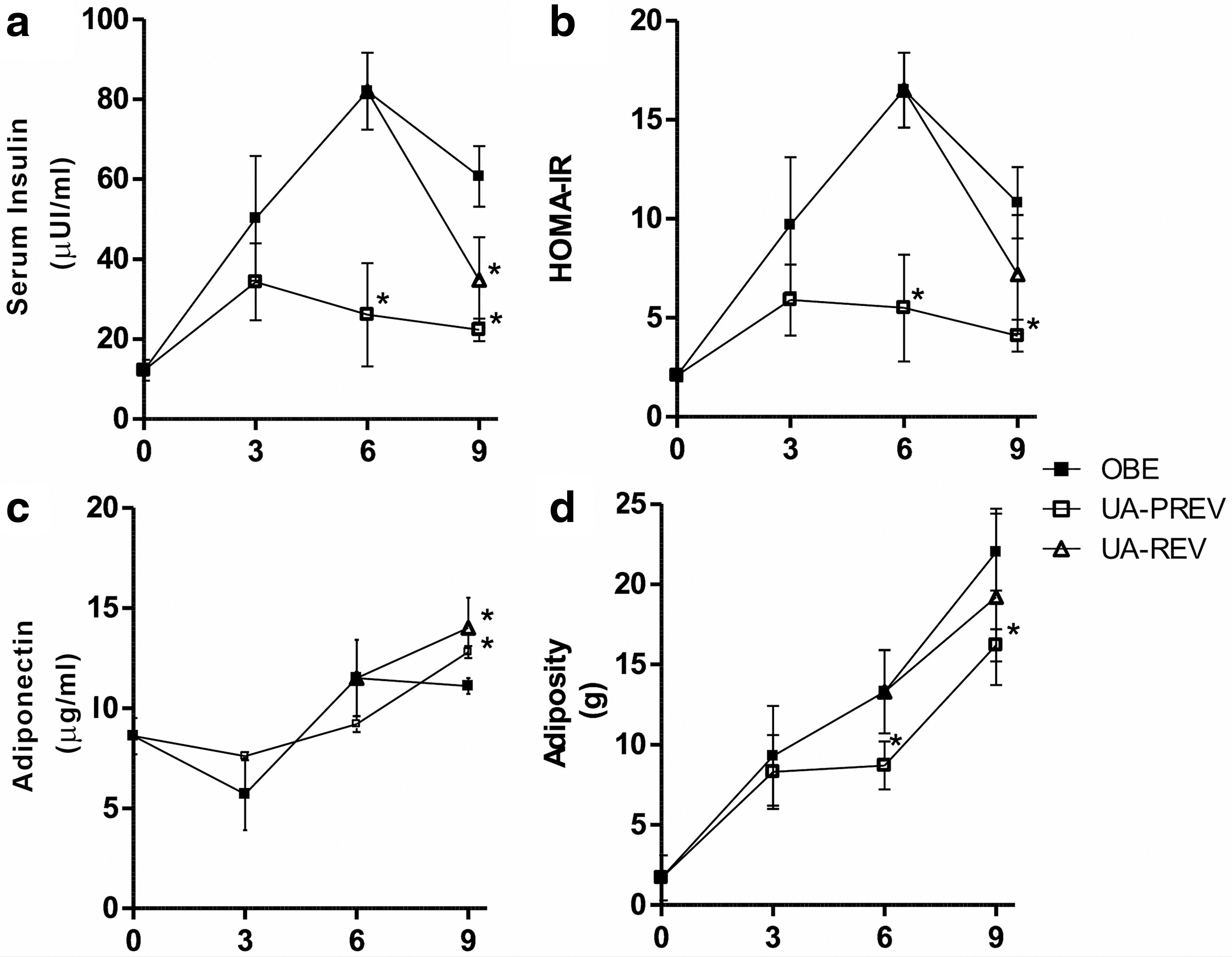
Effect of UA on metabolic markers and adiposity.
Effect of Ursolic Acid on Body Weight, Food Intake, Metabolic and Inflammation Markers in Rats with Diet-Induced Obesity
Values are given as mean ± standard deviation (n = 5 per group).
P < .05 versus OBE.
MCP-1, monocyte chemoattractant protein-1; OBE, high-fat diet group; TC, total cholesterol; TG, triacylglyceride; UA, ursolic acid; UA-PREV, high-fat diet plus UA group; UA-REV, high-fat diet during 6 weeks before simultaneous high-fat diet plus UA for three additional weeks.
UA prevents adipose tissue accumulation and ameliorates lipid profile
Adiposity was measured as the weight of mesenteric, retroperitoneal, epididymal, and perirenal adipose tissue. Adiposity in UA-PREV group after 6 (8.7 ± 1.5 g vs. 13.3 ± 2.6 g, P < .05) and 9 weeks of treatment (16.2 ± 1.0 g vs. 22.0 ± 2.4 g, P < .05), equivalent to 34.6% and 26.4% less, respectively, in comparison with OBE group, was observed (Fig. 1d). Food intake was similar among groups and body weight diminution was not statistically significant. Total cholesterol in UA-PREV group was statistically (P < .05) lower than OBE group after 3 (48.3 ± 4.9 mg/dL vs. 71.0 ± 8.2 mg/dL), 6 (39.6 ± 5.9 mg/dL vs. 62.7 ± 16.8 mg/dL), and 9 weeks of treatment (50.3 ± 4.0 mg/dL vs. 66.0 ± 6.2 mg/dL), and in UA-REV group (47.2 ± 6.1 mg/dL vs. 66.0 ± 6.2 mg/dL). In addition, significant (P < .05) reduction in triacylglyceride levels was observed in UA-PREV group in comparison with OBE group after 3 (57.7 ± 19.6 mg/dL vs. 116.3 ± 17.6 mg/dL), 6 (44.7 ± 7.4 mg/dL vs. 114.3 ± 27.2 mg/dL), and 9 weeks of treatment (80.0 ± 6.2 mg/dL vs. 114.0 ± 17.3 mg/dL), and in UA-REV group (71.0 ± 11.2 mg/dL vs. 114.0 ± 17.3 mg/dL) (Table 2).
UA effect on proinflammatory adipokines expression
Statistically significant reduction (P < .05) in VAT expression of MCP-1, IL-1β, and IL-6, was observed after UA treatment compared with the OBE group. MCP-1 mRNA expression diminished 61% (0.61 times vs. OBE) and 72% (0.72 times vs. OBE) after 6 and 9 weeks of preventive treatment, respectively, and 72% (0.72 times vs. OBE) after the reversal treatment (Fig. 2a). IL-1β mRNA expression reduced 64% (0.64 times vs. OBE) and 72% (0.72 times vs. OBE) after 6 and 9 weeks of preventive treatment, respectively, and 46% (0.46 times vs. OBE) after the reversal treatment (Fig. 2b). IL-6 mRNA expression reduced 64% (0.64 times vs. OBE), 72% (0.72 times vs. OBE), and 71% (0.71 times vs. OBE) after 3, 6, and 9 weeks of preventive treatment, respectively, and 76% (0.76 times vs. OBE) after the reversal treatment (Fig. 2c). TNF-α mRNA expression was not different between groups (data not shown). On the contrary, nonstatistically significant lower circulating MCP-1 levels in UA-PREV group compared with the OBE group after 6 (157.6 ± 27.5 pg/mL vs. 196 ± 25.8 pg/mL) and 9 weeks of treatment (187.5 ± 16.4 pg/mL vs. 216.6 ± 29.8 pg/mL), and nondetectable concentration of IL-1β, IL-6, and TNF-α were observed. UA-REV group presented nonstatistically significant decrease in MCP-1 levels compared with the OBE group (193.2 ± 22.7 pg/mL vs. 216.6 ± 29.8 pg/mL) (Table 2).

Effect of UA on the mRNA expression of proinflammatory adipokines. Expression was quantified by duplicate through qPCR after 3, 6, or 9 weeks of treatment in rats with diet-induced obesity. β-Actin gene was used as a housekeeping gene and results were analyzed by the comparative relative expression CT (2−ΔΔCT) method.
Discussion
The antihyperlipidemic, 28 antiobesity, 32,43,44 and anti-inflammatory activity of UA 31,45 have been well documented. However, this is the first report showing direct evidence of the simultaneous metabolic and anti-inflammatory effects of UA on IR and hyperinsulinemia in an in vivo model of diet-induced obesity evaluating UA preventive and reversal effects after 3, 6, or 9 weeks of treatment, according to metabolic changes reported in animals under HFD supplementation. 36 The limitation of this study was the small size of sample per group owing to the several time points evaluated. IR and hyperinsulinemia, resulting from the insulin signaling cascade disruption because of adipose tissue accumulation and inflammation, have been well established. 16,46 This evidence supports the rationale that nutritional interventions exerting anti-inflammatory effects could offer a suitable alternative to improve insulin action.
Adipose tissue expansion constitutes a key fundamental process in obesity triggering metabolic abnormalities. 18,47 Whereas the ability to predict VAT from circumferences in obese humans has been validated, in animals it can be predicted either by weighing mesenteric, retroperitoneal, epididymal, and perirenal fat depots or even two of them, after dissection. 48,49 In humans, abdominal adiposity, measured by waist circumference, has provided a more reliable mortality predictor than body mass index. It has been reported that VAT reduction of 35% corresponds to ∼10% of total body fat mass loss. 50 In our study, VAT reduction of up to 34.6%, with no significant change in body weight after UA treatment, was observed. This finding could be attributed to UA-induced reduction of adipogenic transcription factors inhibiting lipid accumulation through the LKB1/AMPK (liver kinase B1/AMP-activated protein kinase) pathway and stimulating muscle hypertrophy through Akt increased activation. 44,51,52 However, further studies could help to better understand UA participation in adipocytes maturation or hypertrophic processes.
Amelioration of lipid profile, estimated by significant decrease of triacylglycerides and cholesterol, were evidenced in our study after UA preventive and reversal treatments. These effects could be explained by the reported UA activity on lipid absorption reduction, pancreatic lipase increase, phosphodiesterase inhibition, and diacylglycerol acyltransferase downregulation, the primary enzyme of triglyceride synthesis. 53,54 In addition, the UA-induced reduction of perilipin A expression, a triacylglyceride storage-promoting enzyme, rising of hormone-sensitive lipase translocation and triacylglyceride lipase expression, together with diminution of liver fatty acid synthase activity, increasing of hepatic fatty acid β-oxidation and carnitine palmitoyltransferase activities, are probably also mediating this effect. 55,56 Moreover, the UA inhibitory effect on the intestinal cholesterol acyltransferase activity could explain cholesterol reduction. 57
Hyperinsulinemia constitutes a very important concern in public health because its role in the development of comorbidities associated to obesity has been well established. 5,17 –19 In our study, the reduction of hyperinsulinemia after UA preventive treatment might probably be attributed to the UA insulin-mimetic activity and the enhancement of GLUT-4 translocation. 58 This effect may also be related to the inhibition of the PTP1B (protein tyrosine phosphatase 1B), a downregulating enzyme of the insulin signaling pathway, demonstrated in cell cultures exposed to UA. 59 Consequently, a similar pattern in HOMA-IR was observed when animals simultaneously received HFD and UA for at least 6 weeks. Furthermore, after obesity was established, UA reduced insulin levels but was not capable of significantly reverting IR, probably because of the short exposure to UA reversal treatment. HOMA-IR has been widely used for the estimation of IR in humans and experimental animal models. 60 –62 Moreover, increased levels of adiponectin were observed in UA treated animals, which has been reported to diminish the activity of NF-κB and levels of TNF-α and IL-6 in obese patients reducing the risk for developing IR, hyperinsulinemia, and T2D. 16,63 –66 Based on HOMA-IR and insulin levels, UA effectiveness to improve IR and hyperinsulinemia in obese rats was demonstrated in our study.
Hypertrophic adipocytes accumulated in obese adipose tissue secrete MCP-1 promoting monocytes recruitment and their eventual differentiation to macrophages expressing proinflammatory adipokines IL-1β, IL-6, and TNF-α, propagating the inflammatory response locally within VAT and systemically. 65,67,68 Upregulation of MCP-1 gene expression in VAT is strongly associated with IR in humans irrespective of body mass index. 69 MCP-1 expression in adipose tissue and plasma MCP-1 levels have been found to positively correlate with obesity degree. 11 Chronic inflammation characterizing T2D involves IL-1β, an adipokine progressively destroying insulin-producing β cells or rendering the β cells nonfunctional. IL-1β production is tightly regulated by caspase-1, as shown in macrophages isolated from human fat. 70 IL-6 stimulates insulin secretion and increases SOCS-3 (suppressor of cytokine signaling 3) protein levels in the liver, impairing insulin signaling by decreasing IRS-1 tyrosine phosphorylation, contributing to IR and hyperinsulinemia. 15,71 In this study, the anti-inflammatory activity of UA was exhibited by negatively regulating MCP-1, IL-1β, and IL-6 expression. This effect concurs with the improvement of HOMA-IR, hyperinsulinemia, and adiponectin.
Reduction of IL-1β, IL-6, and TNF-α in melanoma cells treated with UA through the inhibition of NF-κB activation have been documented.
72
Similarly, UA treatment lowered IL-1β, IL-6, TNF-α expression, and NF-κB translocation in mice with
UA dose of 50 mg/kg p.o. daily was selected from a previous dose–response analysis in an animal model showing the most significant activity to counteract experimental inflammation with no toxicity traits or mortality. 29,52 Based on the body surface area normalization method, a human weighing ∼60 kg may benefit from UA metabolic effects by consuming ∼500 mg daily, the equivalent dose of 50 mg/kg daily administered to our animals, 73 and probably might provide better results in patients suffering from metabolic syndrome than clinical trials using lower doses. 74 Our results suggest UA as a suitable diet component that could prevent IR and hyperinsulinemia, or could be useful to revert some of the metabolic abnormalities already exhibited in obese patients. We propose performing additional studies to further elucidate the underlying mechanisms of UA observed in the preventive and reversal assays because several molecular pathways have been described in the onset and development of IR and hyperinsulinemia. Our results open the possibility to evaluate UA effects in other diseases characterized by IR, hyperinsulinemia, inflammation, or hypoadiponectinemia.
In conclusion, UA prevented IR and hyperinsulinemia, and also reversed hyperinsulinemia induced by obesity, probably by decreasing adiposity, entailing a negative modulation of proinflammatory adipokines expression and adiponectin increase (FIG. 3). Our results provide additional evidence of UA potential use in the treatment, prevention, and reverting hyperinsulinemia underlying IR, T2D, and other comorbidities in obese patients.

Proposed mechanism of UA on IR and hyperinsulinemia. Excess of visceral adipose tissue leads to a proinflammatory state through different mechanisms inducing hyperinsulinemia and IR, the main processes contributing to the development of obesity comorbidities. UA, a carboxylic acid with pentacyclic triterpenoid structure, is a major component in a wide variety of foods of plant origin effective to prevent hyperinsulinemia and IR through its antiadiposity and anti-inflammatory activity. IR, insulin resistance.
Footnotes
Acknowledgments
A.S.G.-G., J.G.-B., and B.E.B.-R. conceptualized and designed the research, all authors contributed with the experimental performance; J.G.-B. and B.E.B.-R. wrote the article and contributed equally as corresponding authors.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by a postgraduate fellowship from Consejo Nacional de Ciencia y Tecnología to A.S.G.-G. (No. 224997) and by the research grants from University of Guadalajara to B.E.B.-R. (REC/747/2016, REC/164/2017, REC/CINV/325/2018, REC/CINV/RG/025/2019).