
Abstract
Obesity predisposes people to a variety of chronic metabolic diseases. Identification of natural factors that prevent the development of obesity is likely to be the most successful means of ameliorating the current obesity epidemic. Patchouli alcohol is a sesquiterpene alcohol found in Pogostemon cablin and possesses health benefit activities. This study was designed to examine if patchouli alcohol affects adipogenesis, and investigates the underlying mechanisms whereby patchouli alcohol exerts antiobesity effect. 3T3-L1 adipocytes were differentiated with treatment of different concentrations of patchouli alcohol. An in vivo study was performed to test the effect of patchouli alcohol gavage on a high-fat diet (HFD)-induced obesity. Treatment of patchouli alcohol reduced lipid accumulation in 3T3-L1 adipocytes in a dose-dependent manner without toxicity. Regarding mechanism, treatment of patchouli alcohol reduced expression of peroxisome proliferator-activated receptor-gamma (PPARγ) and CCAAT-enhancer-binding protein-alpha (C/EBPα) and increased expression of total and active β-catenin in 3T3-L1 adipocytes. Oral gavage of patchouli alcohol led to a significant reduction of body weight and fat accumulation in the mice fed with HFD. Transcriptome analysis indicates that smad7 is most highly activated gene in patchouli alcohol-treated 3T3-L1 cells. Patchouli alcohol possesses health benefit effect through inhibiting adipogenesis and fat tissue development.
Introduction
Incidence of obesity sharply increased in the 21st century and is the second most preventable cause of death in the United States. Obesity is highly correlated with incidence of type 2 diabetes and a primary risk factor for diverse metabolic diseases such as cardiovascular risk, stroke, and cancer. Recently, the demand to inhibit these disorders using diverse plants-based compounds has been increasing and related mechanisms are required.
Essential oils have plenty of health benefits with antioxidant, antinociceptive, and antimicrobial activities. 1 Recently a few studies demonstrated that essential oil may prevent metabolic syndrome and could be considered as pharmaceutical agent for prevention and management of obesity. 2 –6 For example, supplementation of lime essential oil led to weight loss. 2 Arteinisia annua L. essential oil decreased accumulation of lipid droplets with downregulation of adipogenic genes such as peroxisome proliferator-activated receptor-gamma (PPARγ), CCAAT-enhancer-binding protein-alpha (C/EBPα), sterol regulatory element-binding protein-1c (SREBP-1c), fatty acid synthase (FAS), and acetyl Co-A carboxylase (ACC) in 3T3-LI cells. 3 Ginger essential oil diminished obesity caused by high-fat diet (HFD) and showed antihyperlipidemic effects by lowering serum-free fatty acid, triglyceride, and total cholesterols. 4 The essential oil from Pinus koraiensis leaves showed antidiabetic effect in streptozotocin-treated mice and pancreatic β cells in vitro 5 and suppressed the expression of obesity- and diabetes-inducing proteins such as SREBPs, 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR), FAS, and glycerol-3-phosphate acyltransferase (GPAT) in HepG2 cells. 6 These data propose the possibility that essential oils could be useful in the treatment of metabolic syndrome, including obesity and diabetes.
Patchouli alcohol (patchoulol) is a sesquiterpene and major component of patchouli. 7 Patchouli alcohol shows anti-inflammatory, 8 –11 antiviral, 12 antiulcer, 13 antiaging, 14 and antidepressant 15 activities. For the first time, our group reported that patchouli alcohol showed anti-inflammatory and anticancer activities in colon cancer cells. 16 Patchouli alcohol ameliorated DSS-induced mice acute colitis by suppressing inflammation 17 and attenuated atherosclerosis, possibly by inhibiting macrophage infiltration and its inflammatory responses. 18 However, the effect of patchouli alcohol on obesity has not been explored so far. This study was designed to investigate effect of patchouli alcohol on obesity, and propose reliable mechanisms focusing on modulation of the β-catenin pathway.
Materials and Methods
Materials
Patchouli alcohol was purchased from Santa Cruz Biotechnology (Dallas, TX, USA) for in vitro study and from Carbosynth (Oxford, UK) for in vivo study (Fig. 1). 3T3-L1 murine preadipocytes were purchased from American Type Culture Collection (Manassa, VA, USA). 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) was purchased from Sigma Aldrich (St. Louis, MO, USA). Antibodies for C/EBPα (cst-8178), PPARγ (cst-2435), β-catenin (cst-9582), and active (nonphosphor) β-catenin (cst-19807) were purchased from Cell Signaling (Danvers, MA, USA). Antibody for actin (sc-1616) was purchased from Santa Cruz Biotechnology. All other chemicals were purchased from Fisher (Waltham, MA, USA) or VWR (Radnor, PA, USA), unless otherwise specified.

Chemical structure and toxicity of patchouli alcohol.
Differentiation of 3T3-L1 preadipocytes and treatment of patchouli alcohol
The 3T3-L1 cells were grown using Dulbecco's modified Eagle's medium (DMEM) containing 10% bovine calf serum at 37°C under an atmosphere of 5% CO2 with replacement of new media every other day. After 2 days postconfluence, 3T3-L1 cells were incubated with media containing dexamethasone-methylisobutylxanthine-insulin (DMI) cocktail (DMEM containing 10% fetal bovine serum (FBS), 1 μg/mL insulin, 1 μM dexamethasone, and 0.5 mM isobutylmethylxanthine) to induce differentiation for 2 days and then incubated with maintenance media (DMEM containing 10% FBS and 1 μg/mL insulin) supplemented with different concentrations of patchouli alcohol for indicated days in figure legends. Patchouli alcohol was dissolved in dimethylsulfoxide (DMSO) and treated in the media as 0.1% (v/v).
Determination of fat accumulation in 3T3-L1 adipocytes
After washing with ice-cold phosphate buffer saline twice, the cells were fixed with 10% buffered formalin for 10 min and washed twice with distilled water to remove formalin. The cells were stained with oil-red O (ORO) solution (0.2% final concentration) for 10 min at room temperature and washed with distilled water four times to remove unstained ORO. The ORO-stained cells were observed under microscope and ORO dye was dissolved in 0.5 mL isopropanol and absorbance of the isopropanol extracts was measured at 500 nm using a microplate reader (Bio-Tek Instruments, Winooski, VT, USA).
Cytotoxicity assay
The cell viability was tested using MTT assay was performed as previously described. 19 In brief, the cells were incubated with MTT solution for 3 h at 37°C and the absorbance was measured in a microplate reader (Bio-Tek Instruments).
Western blot
The cells were lyzed in containing protease- and phosphatase inhibitors. The proteins (30 μg) were separated on SDS-PAGE and transferred to nitrocellulose membranes (Osmonics, Minnetonka, MN, USA). The membranes were incubated with a specific primary antiserum in Tris-buffered saline (TBS) containing 0.05% Tween-20 (TBS-T) and 5% nonfat dry milk at 4°C overnight. After three washes with TBS-T for 30 min, the blots were incubated with HRP-conjugated IgG for 1 h at room temperature, visualized using and quantified by Gel Doc 2000 system (Bio-Rad Laboratories, Hercules, CA, USA).
In vivo study using HFD-induced mouse model
Total 48 C57BL/6J male mice (5-week old) were purchased from Jackson's Laboratories (Bar Harbor, ME) and housed in a temperature- and humidity-controlled environment with 12-h light–dark cycle with food and water available ad libitum. The mice were randomly assigned into four groups and fed with following diets during whole experimental periods; (1) normal diet (control), (2) HFD, (3) HFD with low dose of patchouli alcohol (25 mg/kg body weight), and (4) HFD with high dose of patchouli alcohol (50 mg/kg body weight). The control group (n = 12) received a standard chow diet (F4031; Bioserv, Flemington, NJ) and HFD groups were fed for 12 weeks with diet that 60% kcal is provided from fat. This diet is composed of 36.0% fat, 20.5% protein, 35.7% carbohydrate, and 3.5% ash, and major ingredients are lard, casein, matodextin, sucrose, vitamin and mineral mix,
RNA sequencing for 3T3-L1 treatment of patchouli alcohol
The 3T3-L1 cells, which were treated with DMSO (untreated control), 50, and 100 μM patchouli alcohol, were subjected to RNA sequencing analysis following method described in earlier report.
20
In brief, total RNA was isolated using the RNeasy Plus Mini Kit (Qiagen, Hilden, Germany) and then the quality of isolated RNA was assessed RNA Integrity Number (also known as RIN) using the RNA R6K assay for the Agilent 2200 TapeStation (Agilent Technology, Santa Clara, CA, USA) in which RINs were ranging between 8.5 and 9.5. RNA sequencing library preparation was carried out at the Research Technology Support Facility of Michigan State University (East Lansing, MI, USA). Transcriptome (n = 3 per group) was analyzed through a 1 × 50 bp single-end read using an Illumina HiSeq system (Illumina, Inc., San Diego, CA, USA). The RNA sequence FASTA files mapped to the mouse reference genome (assembly GRCm38.p6;
Statistical analysis
Statistical analysis was performed using one-way ANOVA, followed by Duncan's multiple comparison test. Data are presented as the means ± standard deviation.
Results
Patchouli alcohol does not cause toxicity in undifferentiated and fully differentiated adipocytes
Initially, to see the potential toxicity of patchouli alcohol in undifferentiated and differentiated adipocytes, we treated undifferentiated 3T3-L1 preadipocytes with different concentrations of patchouli alcohol for 24 h and measured cell viability. The results indicate that short-term treatment of patchouli alcohol to preadipocytes is not toxic to the cells (Fig. 1B). Next, we tested long-term cytotoxic activity by treating the different concentrations of patchouli alcohol in differentiated 3T3-L1 adipocytes for 8 days. No toxicity was observed, but slight increase of cell viability was detected in the differentiated adipocytes treated with 75 and 100 μM patchouli alcohol (Fig. 1C).
Patchouli alcohol inhibits differentiation of preadipocytes and triglyceride accumulation in a dose-dependent manner
Differentiation of 3T3-L1 adipocyte is initiated by exposure to DMI cocktail and undergoes three distinct stages, including the early stage, the postmitotic intermediate stage, and the terminal stage. 21 First, to examine if patchouli alcohol affects adipogenesis and triglyceride accumulation at early stage of differentiation (with treatment of DMI cocktail), 3T3-L1 preadipocytes were exposed to different concentrations of patchouli alcohol during both differentiation (2 days) and postdifferentiation stage (8 days) for 10 days (Fig. 2A) and then triglyceride content was compared with ORO staining (Fig. 2B). As a result, triglyceride accumulation was significantly reduced in the cells treated with 25, 50, 75, and 100 μM patchouli alcohol (An inhibition of 40.9%, 38.0%, 47.7%, and 71.4% from the cells treated with 25, 50, 75, and 100 μM patchouli alcohol, respectively). Next, to examine if patchouli alcohol affects adipogenesis and triglyceride accumulation postdifferentiation stage (2 days after treatment of DMI cocktail), 3T3-L1 preadipocytes were differentiated for 2 days with DMI cocktail first and exposed to different concentrations of patchouli alcohol for 8 days (Fig. 3A) and then triglyceride content was compared with ORO staining (Fig. 3B). Significant decrease of triglyceride accumulation was observed in the cells treated with 12, 25, 50, 75, and 100 μM patchouli alcohol (An inhibition of 12.9%, 13.3%, 52.7%, 78.4%, and 80.4% from the cells treated with 12, 25, 50, 75, and 100 μM patchouli alcohol, respectively).

Patchouli alcohol treatment with differentiation inhibits triglyceride accumulation and altered expression of PPARγ, C/EBPα, total- and active β-catenin.
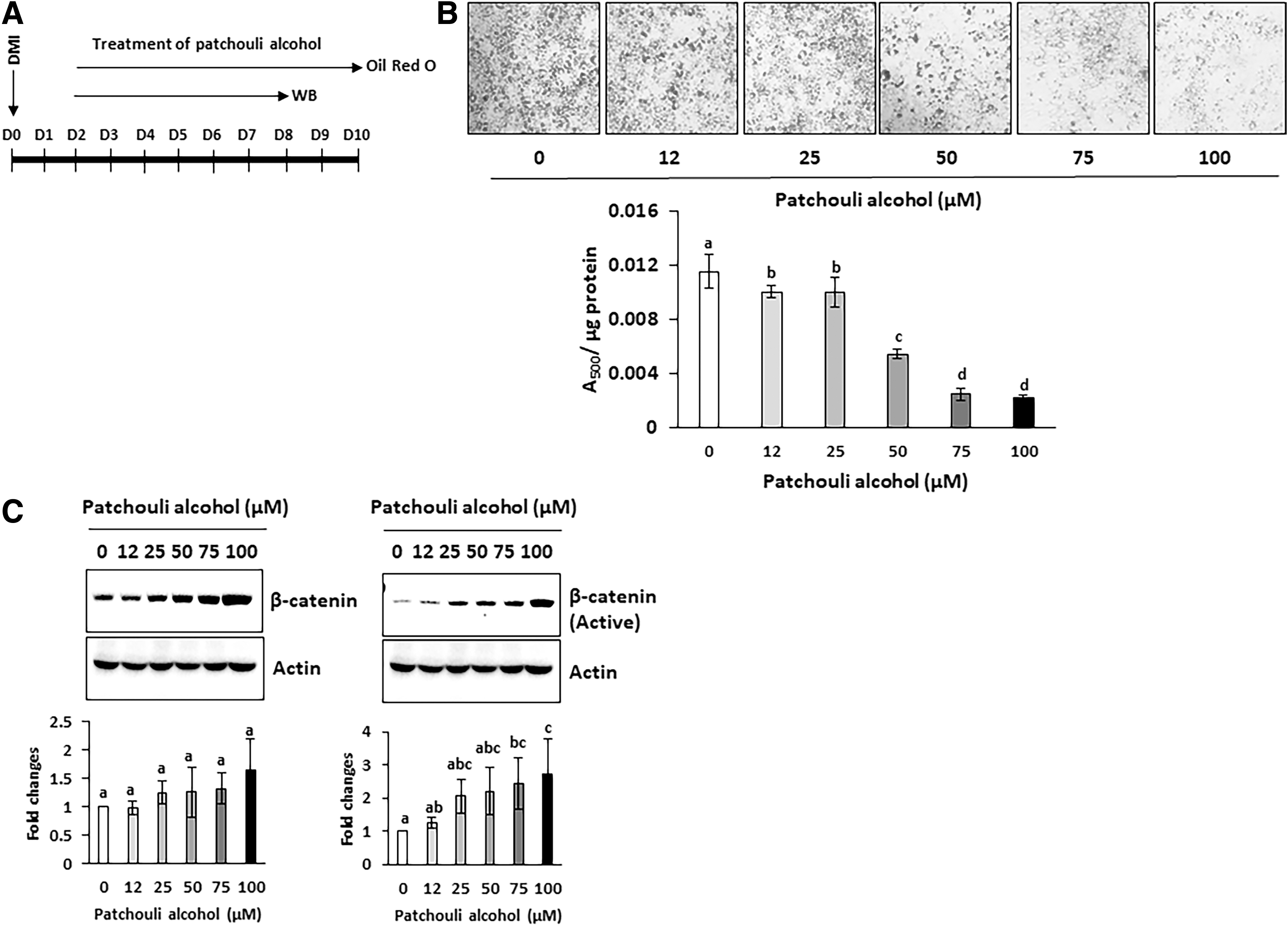
Patchouli alcohol treatment after differentiation inhibits triglyceride accumulation and altered expression of total- and active β-catenin.
Patchouli alcohol downregulates C/EBPα and PPARγ, and upregulates β-catenin in differentiated 3T3-L1 cells
The PPARγ and C/EBPα are most well-defined transcriptional triggers of adipocytes differentiation during early time points (within 2 days). To elucidate the molecular mechanisms for inhibition of adipogenesis, we measured expression of these protein in the cells exposed to DMI with or without patchouli alcohol for 3 days (Fig. 2A). As shown in Figure 2C, the marked decrease of C/EBPα and PPARγ were observed in differentiated adipocytes treated with patchouli alcohol, compared with nontreated control group (31.5%, 27.3%, and 58.8% inhibition of PPARγ; 45.6%, 48.6%, and 77.3% inhibition of C/EBPα at 50, 75, and 100 μM patchouli alcohol-treated cells). These changes in molecular event by patchouli alcohol could be a mechanism for suppression of adipocyte differentiation.
In contrast, Wnt signaling is negatively associated with adipogenesis 22 and β-catenin is a major mediator linking the upstream Wnt signaling to target gene expression. To investigate if treatment of patchouli alcohol affects amount of total and biologically active β-catenin, we performed Western blot using antibodies against total and active β-catenin (nonphospho at Ser45). Expression of total β-catenin was significantly increased in the 75 μM (24.6% increase) and 100 μM (36.6% increase)-treated cells, whereas active β-catenin tends to increase by the treatment of patchouli alcohol (Fig. 2C). Similar trends are observed in Figure 3C.
Patchouli alcohol decreases body weight and fat accumulation in HFD-induced obese mouse model
We investigated if patchouli alcohol possesses antiobesity activity using HFD-induced obese model because HFD is known to promote obesity in mice. 23 As shown in Figure 4A, the mice were exposed to normal diet or HFD for 12 weeks (from 5 to 17 week) and patchouli alcohol was fed to HFD-fed mice through oral gavage three times per week for 8 weeks (from 9 to 17 week). There were no significant weight differences between the groups before the start of the experiments (5-week age). A dramatic increase of body weight was observed in the mice fed with HFD compared with those fed with normal diet for 12 weeks. Treatment of patchouli alcohol resulted in a significant decrease of body weight in dose-dependent manner. The statistical significance was observed from 16 to 17 weeks. To determine whether reduced body weight by patchouli alcohol treatment resulted from reduced fat accumulation, we compared abdominal fat accumulation between control and patchouli alcohol-treated mice. Accumulation of abdominal fat was dramatically reduced in patchouli alcohol-treated mice compared with control mice (Fig. 4B). Epididymal, retroperitoneal, and brown fat pad were isolated and their weights were compared. An analysis of individual fat pad weights (expressed as milligram fat pad weight per gram body weight) revealed a significant decrease of epididymal and retroperitoneal fat pads in patchouli alcohol-treated mice, whereas brown adipose tissue was not significantly altered (Fig. 4C).
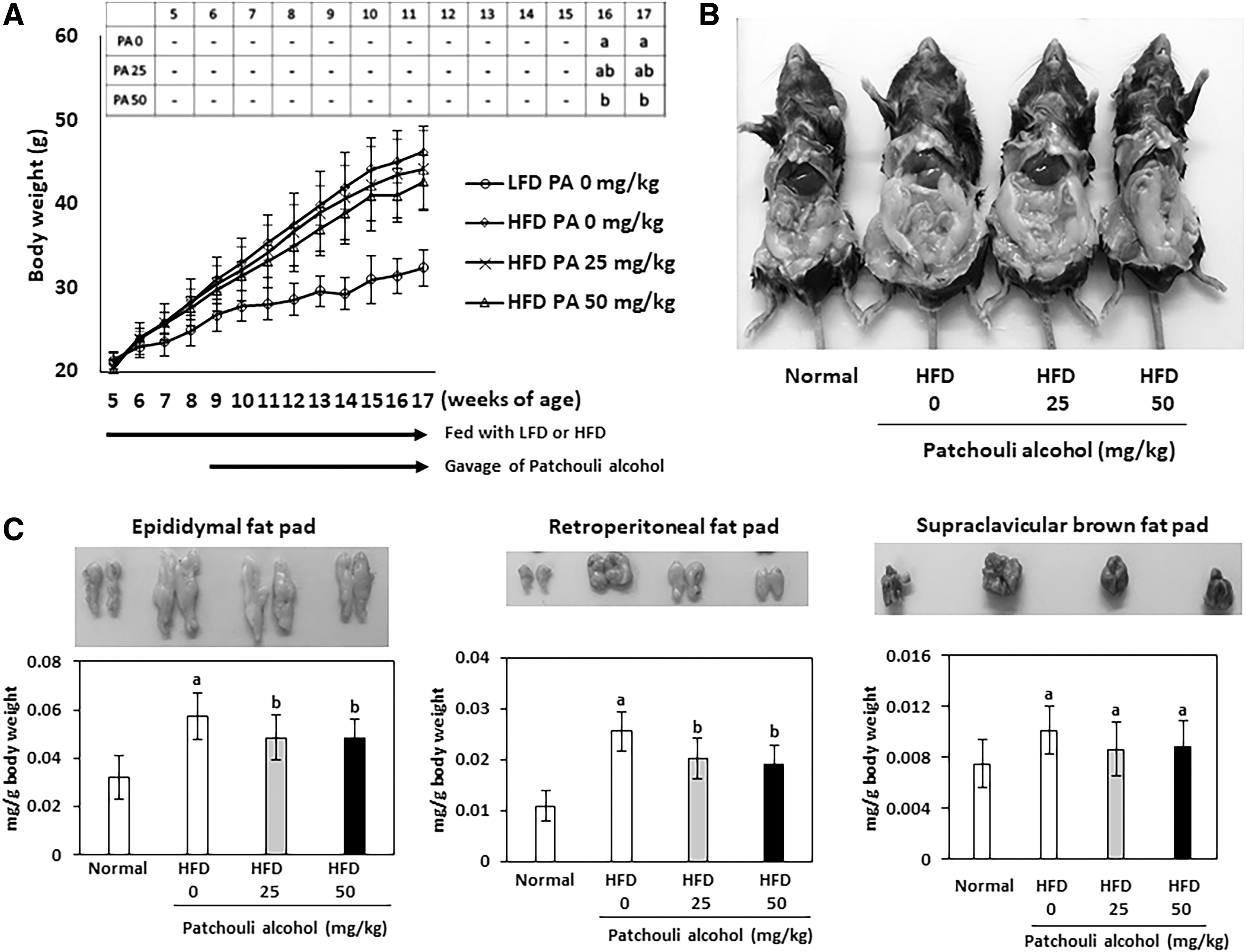
Patchouli alcohol decreases body weight and fat accumulation HFD-induced obesity mouse model.
Transcriptome analysis of 3T3-L1 cells treated with patchouli alcohol
To identify transcriptomic differences in 3T3-L1 cells, which were treated with patchouli alcohol, RNA sequencing analyses conducted. At the treatment of 100 μM patchouli alcohol (PA100), 18 genes were DE at FDR <0.05 and log2-fold change (FC) >1 or log2FC <-1 compared with untreated group (Con) (Table 1). Interestingly, all 18 DE genes in PA100 showed the less degree of fold changes in the treatment group of 50 μM patchouli alcohol (PA50), although all FDR values were not in significant levels in PA50 group.
Differentially Expressed Genes in 3T3-L Cells Treated with 100 μM Patchouli Alcohol (PA100)
Differential expressions compared with untreated group (Con) were indicated with log2-FC values. Values of log2FC and FDR derived from the group of 50 μM patchouli alcohol treatment (PA50) for designated genes were displayed.
FC, fold change; FDR, false discovery rate.
Discussion
Essential oils are natural products that were derived from aromatic plants and traditionally used all over the world for diverse purposes such as disinfection, anti-inflammation, relaxing, and potential exploitation in clinical medicine. 24 Pogostemon cablin (also called patchouli) is a well-known essential oil in Chinese medication and its proposed therapeutic effects include antiviral, antioxidative, analgesic, anti-inflammatory activities, and intestinal barrier function protection. 25 One of the most recently proposed health benefit properties by essential oils is metabolism and energy balance. 2 –6
Patchouli alcohol is a sesquiterpene alcohol (patchoulol), which is the characteristic component of Pogostemon cablin 7 (Fig. 1A). Previously, our research group reported that patchouli alcohol repressed inflammatory signaling in LPS-stimulated RAW264.7 and tumor necrosis factor-alpha-stimulated HT-29 cells through repressing IκB-α degradation, p65 nuclear translocation, and ERK1/2 pathway. 8 However, no study has examined the association between patchouli alcohol and obesity. In this study, for the first time, we investigated if patchouli alcohol influences body weight regulation and development of obesity. Our data indicate that patchouli alcohol possesses antiobesity activity in HFD-induced obese mouse model and in vitro model using differentiated adipocytes.
We designed two in vitro studies to see if treatment of patchouli alcohol at different time point (before and after differentiation) affects triglyceride accumulation. As shown in Figures 2A and B and 3A and B, similar inhibition of fat differentiation was observed from the cells treated with patchouli alcohol between earlier (D0) and later time point (D2). This indicates that antiadipogenic activity of patchouli alcohol is not associated with the change of molecular event that occurs with treatment of DMI cocktail during 48 h.
Since feeding HFD results in obesity and associated metabolic syndrome, we adopted this model to explore the possibility that patchouli alcohol possesses suppressive activity of adipose tissue development. Significant inhibition of body weight and fat accumulation was observed in patchouli alcohol-treated group (Fig. 4C), implying adipose tissue could be one of the primary organ associated with patchouli alcohol benefit.
Activated Wnt/β-catenin signaling is associated with inhibition of preadipocyte differentiation 22 and PPARγ suppresses Wnt/β-catenin signaling during adipogenesis. 26 Recently several studies reported that suppression of adipogenesis by bioactive compounds could be mediated by increased expression of β-catenin. 27 –33 The current results also indicate that patchouli alcohol stimulates expression of total- and active β-catenin in dose-dependent manner and suppressed adipogenesis (Fig. 2C, 3C). Thus, we speculate that antiadipogenic activity of patchouli alcohol may be due to inhibition of the signaling pathway from Wnt and β-catenin.
Interestingly, we identified smad7 as a novel molecular target of patchouli alcohol (Table 1). Smad7 expression was significantly increased in dose-dependent manner by treatment of patchouli alcohol. We also found that mRNA of β-catenin was not changed by treatment of patchouli alcohol (data not shown), indicating that patchouli alcohol does not lead to transcriptional upregulation, instead increases the stability of β-catenin. This speculation is supported by a strong interaction between smad7 and β-catenin that were reported recently. Smad7 is required for TGF-β1-induced accumulation of β-catenin, whereas repression of smad7 expression prevented TGF-β-induced activity of β-catenin. 34 And smad7 stabilizes β-catenin and increase binding to E-cadherin complex, thus affecting cell–cell adhesion. 35 Moreover, smad7 and β-catenin are cooperatively recruited to promoter region of their target gene. 36 These findings imply that increased expression of smad7 and subsequent stabilization of β-catenin might contribute to patchouli-alcohol-induced prevention of obesity.
In conclusion, this study demonstrated that patchouli alcohol suppressed adipogenesis and fat accumulation in adipocytes. These beneficial effects might be mediated by increased expression and active β-catenin.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study is supported by NIFA Hatch grant [MD-HNFS-9897] from the USDA National Institute of Food and Agriculture.