
Abstract
The maintenance of a healthy skin barrier is crucial to prevent and treat atopic dermatitis (AD) lesions and avoid infections. Glycomacropeptide (GMP) is a bioactive peptide that has demonstrated promising results as an anti-inflammatory and antipruritic therapy for experimental AD. This study aimed to analyze the effect of GMP on impaired cutaneous barrier-related signs in a rat model of AD lesions. AD-like dermatitis was induced on the skin by repeated topical applications of 2,4-dinitrochlorobenzene, and animals were orally administered GMP before or after AD induction. The expression of skin structural proteins and antimicrobial peptides (AMPs) was evaluated by immunoblot or immunohistochemistry, epidermal thickening was evaluated by histochemistry, the level of IFN-γ and changes in the microbiota were evaluated by quantitative polymerase chain reaction, and the quantity of fecal short-chain fatty acids (SCFAs) was evaluated by gas chromatography. GMP administration significantly increased filaggrin, β-defensin 2, and cathelicidin-related AMP expression in AD-like lesions. Involucrin expression was not modified. In GMP-treated animals, epidermal thickening and IFN-γ expression were strongly reduced in damaged skin. GMP treatment impacted the skin microbiota and prevented Staphylococcus aureus colonization, which is associated with AD. In addition, high levels of Bifidobacterium were detected in the feces of GMP-treated animals, and the acetic acid and butyric acid contents increased in animals prophylactically administered GMP. These results suggest that GMP markedly prevents or reverses skin barrier damage in rat AD-like lesions through a bifidogenic effect that induces fecal SCFA production with prolonged treatment. Our findings provide evidence that GMP may represent an optimum strategy for the therapy of the dysfunctional cutaneous barrier in AD.
Introduction
Atopic dermatitis (AD) is a chronic inflammatory skin condition that affects ∼10%–20% of children in developed countries and up to 5% of adults. 1,2 AD has a complex etiology, with interactions between genetic and environmental factors that induce abnormalities in the epidermal barrier and immune system. 3
Evidence supports that the pathogenesis of AD is initiated by an impaired epidermal structure and function that later causes inflammation, T-helper (Th)1/Th2 cell dysregulation, immunoglobulin E production, and mast cell hyperactivity, which further provokes secondary barrier abnormalities. 4 The most common defect in skin barrier is a decrease in the production of filaggrin or other proteins of the stratum corneum, which contributes to the induction of dry skin and the susceptibility to infections. 5,6 Besides, skin bacterial colonization is augmented by the reduced production of antimicrobial peptides (AMPs). 7
Bioactive peptides derived from dietary proteins are beneficial to human health. Milk is one of the main sources of bioactive peptides, and among them, glycomacropeptide (GMP) is broadly studied. 8 It is derived from κ-casein by enzymatic activities, is composed of 64 amino acids, and is highly glycosylated with units of N-acetyl neuraminic (sialic) acid. 9 Because it is safe, is nonimmunogenic, and has nutritional properties, it is used in the food and pharmaceutical industries. 9 –11
Different experimental approaches have highlighted the prebiotic, anti-inflammatory, and immunoregulatory capacities of GMP. 12,13 In our previous studies on experimental AD, we showed that administering GMP to Wistar rats decreased inflammation, pruritus, and the Th2 response. 14 However, it is known that anti-inflammatory therapy is not sufficient in the management of AD and that the restoration of skin barrier function is essential to avoid future complications, such as opportunistic infections. 15 In addition, the anti-inflammatory effect of natural substances in experimental AD is not always linked to the improvement of the altered skin barrier. 16 Thus, to propose this bioactive peptide as a potential therapy in the prevention or effective management of AD, its effect on skin barrier alteration must be analyzed.
In this study, we evaluated whether oral GMP administration influenced the dysfunction in the epidermal barrier associated with 2,4-dinitrochlorobenzene (DNCB)-induced AD-like lesions in rats.
Materials and Methods
Animals
Male Wistar rats (10-week old) were acquired from the Laboratory Animal Service of the Autonomous University of Aguascalientes. Animals were maintained under controlled temperature (22°C–24°C) and illumination (12-h light cycles) conditions. Food (Rodent Laboratory Chow 5001; Purina, Mexico) and tap water were provided ad libitum. All animal experiments were approved by the animal care committee of the Autonomous University of Aguascalientes (Approval date: August 11, 2014).
Rats were randomly divided into five groups and two independent protocols were developed (n = 4 rats/group/protocol): group administered water (control); AD group prophylactically administered water (AD-WP); AD group prophylactically administered GMP (AD-GP); AD group therapeutically administered water (AD-WT); and AD group therapeutically administered GMP (AD-GT).
Atopic dermatitis induction and treatment
To induce AD, animals were sensitized on day 1 with an intramuscular injection of 1 mg of dinitrophenyl-bovine serum albumin (Sigma-Aldrich, St. Louis, MO, USA) prepared in 7.8 mg of aluminum hydroxide gel (Thermo Scientific, Waltham, MA, USA), and a subcutaneous injection of 0.5 mL of Bordetella pertussis vaccine (Biofarma, Bandung, Indonesia) as an adjuvant. On day 14, 16, 18, 20, and 22, rats were resensitized with topical applications of 60 μL of 0.015 g/mL DNCB in the ear lobe (Sigma-Aldrich) prepared in acetone:olive oil solution (4:1), and on day 36, rats received a final DNCB challenge.
GMP (gifted by Arla Food Ingredients Group P/S, Viby, Denmark) was dissolved in tap water and administered using an esophageal catheter at a dose of 500 mg/kg/day. Control group was managed only with adjuvants and vehicles. 14 Prophylactic treatment started 3 days before intramuscular sensitization and therapeutic treatment on day 23, and both finished on day 36. Animals were euthanized by inhaled anesthesia on day 37, and ear samples were obtained.
Western blot analysis
Rat ear tissue was excised, frozen in liquid nitrogen, and homogenized with phosphate-buffered saline. Proteins were extracted using lysis buffer with protease inhibitors, and the supernatant was separated by centrifugation at 16,435 g for 20 min. Proteins were quantified by the Bradford method, and equal amounts of total proteins were separated by 10% SDS-PAGE and blotted onto a PVDF membrane. Membranes were incubated with monoclonal anti-involucrin (1:1000; Abcam, Cambridge, MA, UK) or anti-β-actin (1:1000; Sigma-Aldrich) antibodies overnight at 4°C, washed, incubated with an alkaline phosphatase-conjugated secondary antibody (1:5000; Zymed, San Francisco, CA, USA), and later detected using SIGMAFAST (Sigma-Aldrich). Densitometric data were expressed as levels relative to those of β-actin.
Histological analysis
Ear skin samples were fixed in formalin and embedded in paraffin. Sections 5 μm thick were stained with hematoxylin and eosin for epithelium thickness evaluation at 400 × in a microscope Carl Zeiss (Oberkochen, Germany).
In addition, sections of 3 μm thickness were mounted on poly-
Finally, slices were counterstained with Harris′ hematoxylin and photographed using a light microscope at 400 × magnification.
RNA purification and quantitative polymerase chain reaction
Total RNA was extracted from ear tissue using the SV Total RNA Isolation System (Promega, Madison, WI, USA). First-strand cDNA was synthesized using the oligo dT primer with the RETROscript Reverse Transcription kit (Thermo Scientific). Quantitative polymerase chain reaction (qPCR) was performed with the GoTaq qPCR Master Mix (Promega) in a StepOne Real-Time PCR System (Thermo Scientific). The mRNA expression of IFN-γ was quantified with the primers shown in Table 1. Relative quantification was determined with the 2−ΔΔCt method, normalizing data to β-actin as a housekeeping gene. 17
Primer Sequences Used for Relative and Absolute Quantitative Polymerase Chain Reaction
DNA isolation and quantitative polymerase chain reaction
To determine the change in Staphylococcus aureus and Staphylococcus epidermidis, we employed an approach based on a described protocol with adaptations. 18 Briefly, genomic DNA was extracted from skin samples with the ZymoBead Genomic DNA Kit (Zymo Research, Irvine, CA, USA). For qPCR, reactions were mounted with the GoTaq qPCR Master Mix (Promega) in a StepOne equipment. For absolute quantification, amplicons from PCR targeting the femA gene of S. aureus and S. epidermidis were cloned using the InsTAclone PCR Cloning Kit (Thermo Scientific). The resulting plasmids, pTZ57R-FemA-SA and pTZ57R-FemA-SE, were purified with the PureYield Plasmid Maxiprep System (Promega) and used for 10-fold serial dilutions of femA gene copy numbers.
For the fecal contents of Lactobacillus/Bifidobacterium/Bacteroides, fecal DNA was purified from 100 mg of feces using the ZR Fecal DNA Miniprep kit (Zymo Research), and 16S gene copies of Lactobacillus/Bifidobacterium/Bacteroides were determined by qPCR with a previously reported method. 19 Primers used for absolute quantifications of skin and fecal microbiota are listed in Table 1.
Determination of short-chain fatty acid levels
Fecal samples (100 mg) were suspended in 200 μL sterile deionized water by vortexing. After centrifugation at 16,000 g for 10 min, the supernatant was collected and measured in a gas chromatograph 6850 Network GC System (Agilent Technologies, Santa Clara, CA, USA) with an HP-5MS column coupled to a mass spectrometer 5975C VL MSD with a triple-axis detector. The initial temperature in the oven was 100°C with an increment rate of 15°C/min. The injector and detector temperatures were both 270°C, and helium was used as the carrier gas at a 1.5 mL/min flow rate. Individual acetic acid and butyric acid concentrations were determined by comparing peaks with a standard curve established with the WSFA-2 standard (Sigma-Aldrich).
Statistical analysis
Data from each group were expressed as mean ± standard error of the mean (SEM) and statistically analyzed by multicomparative Bonferroni test using GraphPad Prism 5.01 statistical software. Data of the abundances of S. epidermidis and S. aureus were analyzed by Kruskal–Wallis tests and Dunn's multiple comparison post-test. Significance was set at P < .05.
Results
Effect of glycomacropeptide on stratum corneum protein expression
To estimate the effect of GMP administration on skin barrier proteins, we analyzed the levels of filaggrin by immunohistochemical analysis (Fig. 1A). Morphometric assessment showed that compared to control animals, AD animals showed an average reduction of 69.13% (P < .001) in filaggrin immunoreactivity in skin tissue. GMP used prophylactically avoided the loss of filaggrin in AD lesions, as filaggrin level was similar to that in control group. Therapeutic GMP administration recovered 1.84-fold (P < .001) filaggrin expression (Fig. 1B).
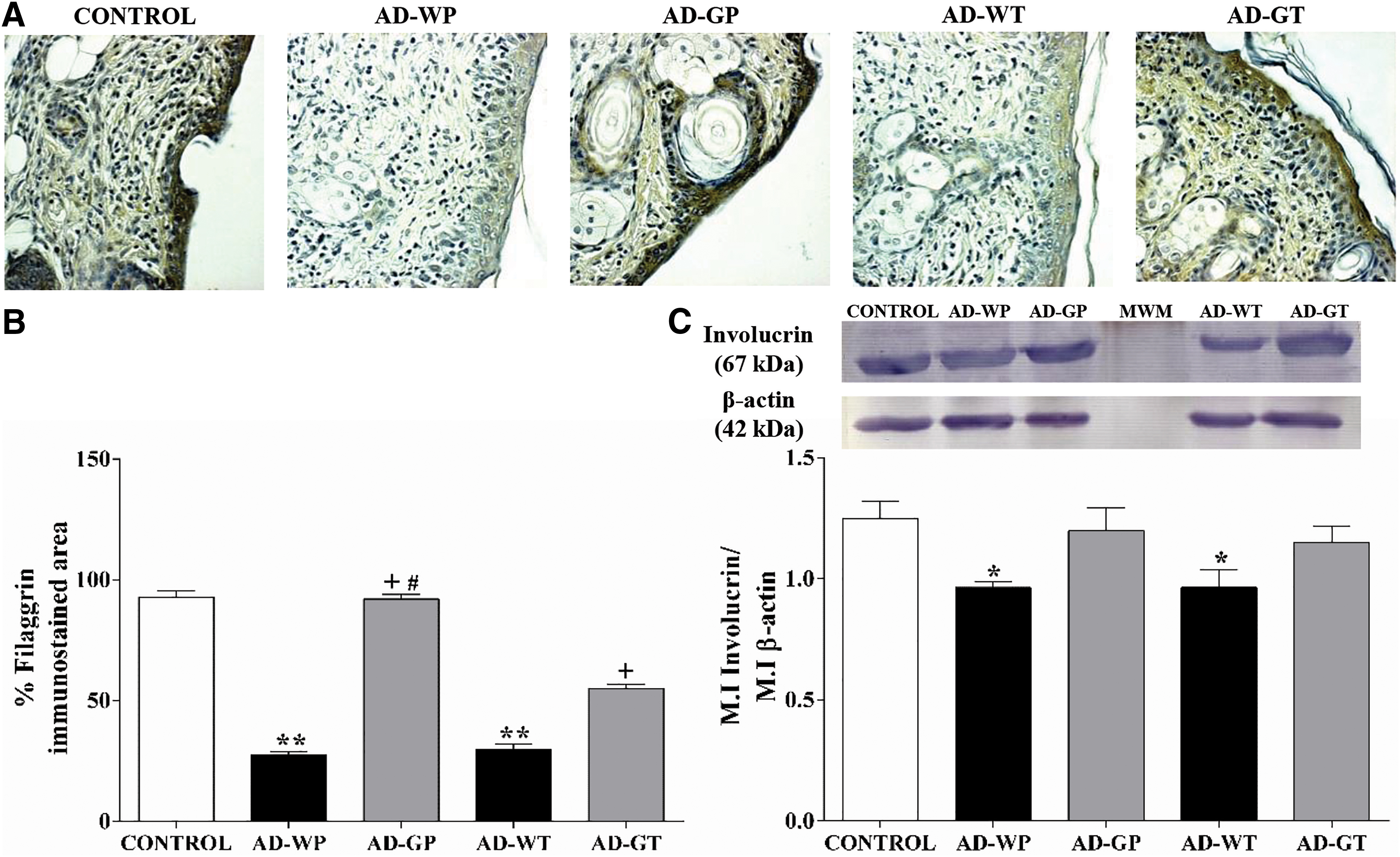
Effect of GMP administration on filaggrin and involucrin expression in AD-like lesions.
We analyzed the skin level of involucrin, a structural protein of the cornified envelope, by Western blot. The involucrin levels in animals with DNCB-induced skin lesions were reduced by 22.85% (P < .05). GMP treatments slightly increased involucrin expression, although it was not significant (Fig. 1C).
Stimulatory effect of GMP on antimicrobial peptide expression
AMPs produced by epidermal cells are part of the skin barrier. When skin is disrupted as a consequence of AD, AMP production is downregulated. 20 To determine whether the oral intake of GMP could prevent the downregulation of AMP expression, we detected BD-2 and CRAMP (Fig. 2A) by immunohistochemistry in the skin of experimental animals.
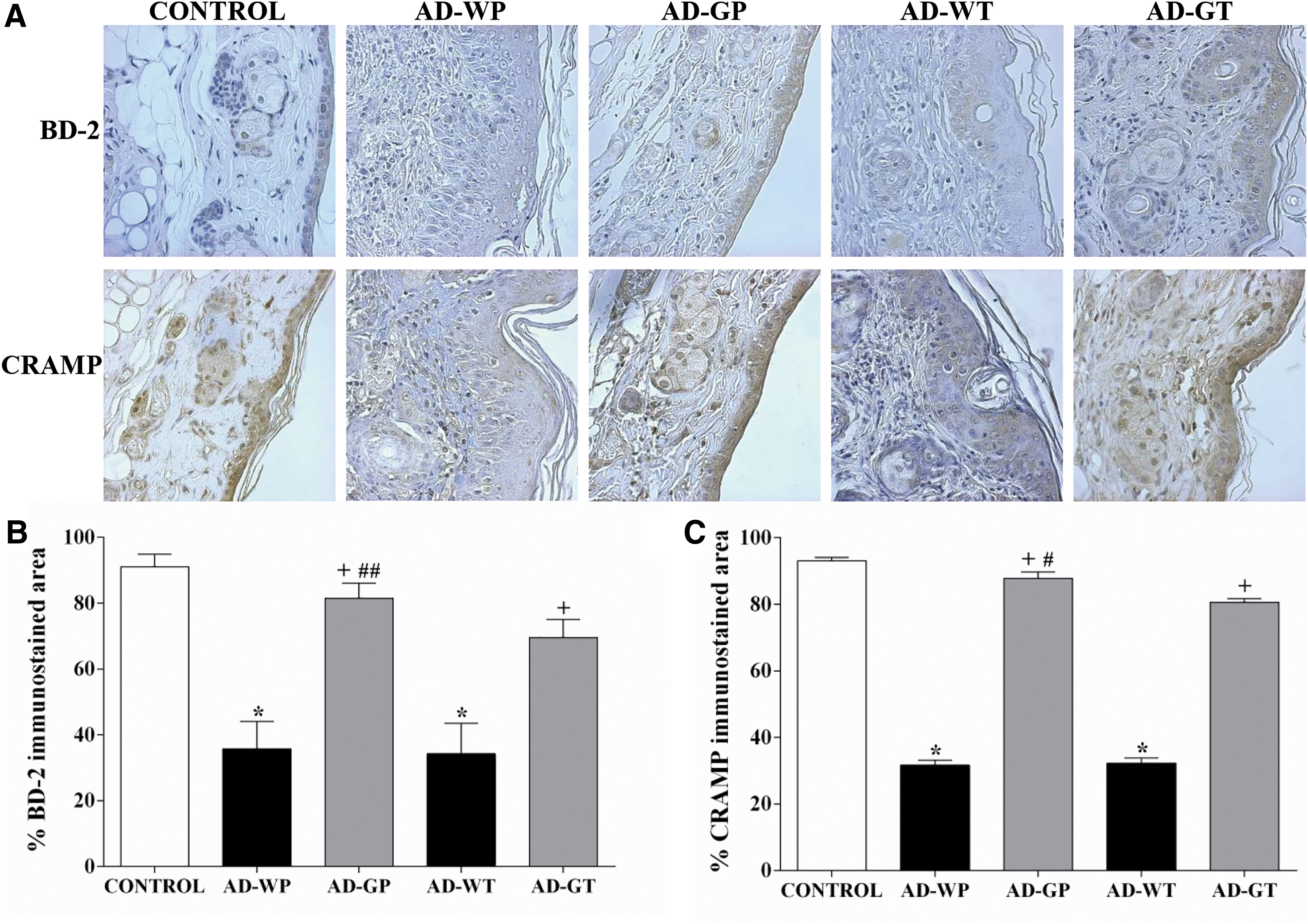
Effect of GMP administration on BD-2 and CRAMP expression in AD-like lesions.
As shown in the immunostaining analysis (Fig. 2B, C), compared to control group, the AD-WP and AD-WT groups exhibited reductions of 60.68% and 62.36% (P < .001) in immunopositivity to BD-2 and of 65.97% and 65.30% (P < .001) to CRAMP, respectively. However, animals pretreated with GMP prevented the downregulation of BD-2 and CRAMP expression in skin AD lesions, as immunostained areas were unchanged compared to those in control animals. Furthermore, compared to untreated animals, animals administered GMP in a therapeutic manner recovered 2.03- and 2.49-fold BD-2 and CRAMP expression (P < .001).
Inhibitory effect of GMP on epidermal thickness
Skin dermatitis is characterized by epidermal hyperplasia, 21 which occurred in our experimental model (Fig. 3A). Morphometric assessment revealed that the epidermis of AD-WP and AD-WT rats was 23.80 ± 0.99 and 26.86 ± 1.1 μm thick, whereas the epidermis of control rats was only 10.47 ± 0.62 μm (Fig. 3B). In the AD-GP and AD-GT groups, the epidermis was 46.51% and 28.96% (both P < .001) thinner than those in the AD-WP and AD-WT animals, respectively.

Effect of GMP administration on epidermal thickness and IFN-γ expression.
Because IFN-γ is related to epidermal thickening in AD lesions, 22 we investigated whether GMP modulates IFN-γ mRNA expression. We found that the AD-WP and AD-WT groups exhibited a 7.93- and 9.19-fold (P < .001) increase in IFN-γ expression. Interestingly, prophylactic GMP administration decreased IFN-γ expression by 88.27% (P < .001) and by 68.95% (P < .001; Fig. 3C) when it was therapeutically administered.
Effect of GMP on the S. epidermidis/S. aureus balance
An impaired epidermal barrier and altered innate immune response during AD contribute to changes in skin microbiota with susceptibility to S. aureus colonization. 23 We investigated the amount of S. epidermidis and S. aureus in injured cutaneous tissue in rats. In our AD model, the amount of S. epidermidis was similar among the five groups, with an average number of 169.35 femA gene copies/10 ng of DNA (Fig. 4). Interestingly, the number of copies of S. aureus increased in untreated AD animals, but it was only significant in the AD-WT group.
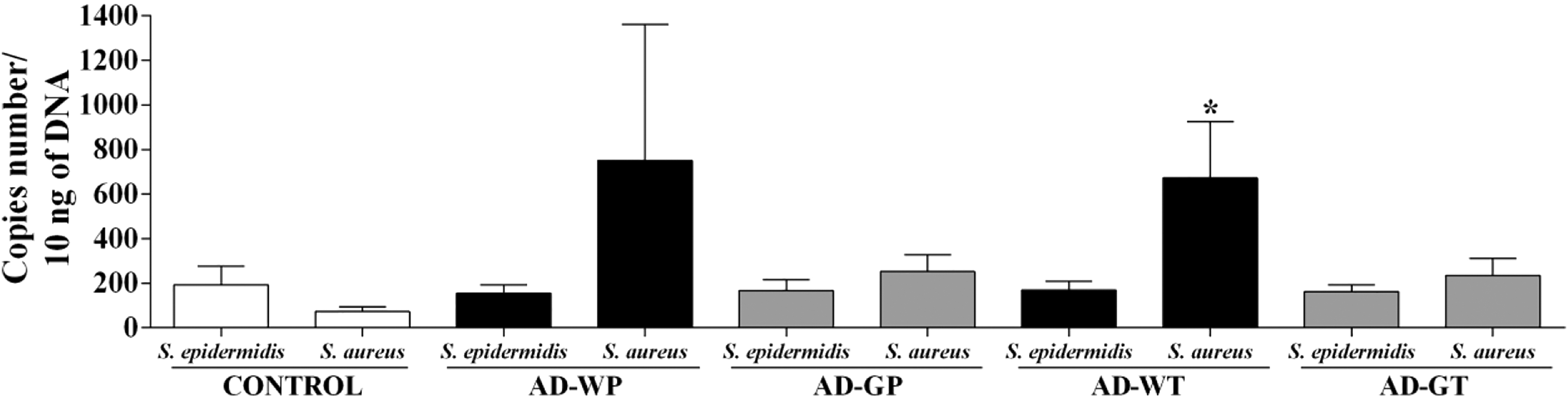
Effect of GMP on cutaneous amounts of Staphylococcus epidermidis and Staphylococcus aureus. Quantification of S. epidermidis and S. aureus was analyzed as the number of copies of each bacterial species in 10 ng of extracted DNA per rat. Data from each group are expressed as the mean ± SEM. *P < .05 versus control.
When we analyzed the ratio between S. aureus and S. epidermidis in each group, we observed that in control animals, S. aureus was 2.6 times less abundant than S. epidermidis, but in AD animals, the number of copies of S. aureus was 4.85-fold (AD-WP) and 3.96-fold (AD-WT) greater than those of S. epidermidis. GMP administration clearly showed a tendency to decrease the ratio of S. aureus/S. epidermidis in the AD-GP and AD-GT groups.
Effects of GMP on the intestinal environment
Due to the prebiotic activity of GMP, 24 we measured the amount of Lactobacillus, Bifidobacterium, and Bacteroides in feces. Lactobacillus was present at a higher concentration than Bifidobacterium and Bacteroides in the fecal microbiota of the control rats (Table 2). The levels of the three bacterial genera in AD rats showed no change compared to those in control rats. Logarithmic amounts of Bifidobacterium increased significantly by 2.43- and 2.85-fold in samples of AD rats prophylactically and therapeutically treated with GMP. Lactobacillus only increased 1.92-fold in the AD-GT group, and Bacteroides remained similar between treated and untreated AD animals.
Microbiota Measurement in Feces by Quantitative Polymerase Chain Reaction Targeted to 16 S Genes
P < .05 versus AD-WP, ++ P < .0001 versus AD-WT, and # P < .0001 versus AD-GP.
AD, atopic dermatitis; AD-GP, group with AD prophylactically administered GMP; AD-GT, group with AD therapeutically administered GMP; AD-WP, group with atopic dermatitis prophylactically administered water; AD-WT, group with AD therapeutically administered water; GMP, glycomacropeptide; SD, standard deviation.
We also evaluated the fecal short-chain fatty acid (SCFA) levels, as Bifidobacterium has been associated with increased acetate and butyrate production. 25,26 The concentrations of acetic acid and butyric acid in fecal samples were similar between the control and AD animals (Fig. 5A, B). However, the acetic acid and butyric acid levels in animals orally administered GMP for 39 days (AD-GP) increased 2.03-fold (P < .001) and 1.8-fold (P < .01) in the fecal content compared to those in the untreated animals. When animals received GMP in a therapeutic manner (14 days of intake), the fecal SCFA levels were unchanged.

Effect of GMP administration on intestinal SCFA levels. Fecal samples (100 mg) from each rat were analyzed by gas chromatography to quantify acetic acid
Discussion
AD is a chronic skin disease with severe negative effects on the quality of life and financial stability of patients and their families. Currently, the maintenance and repair of a healthy skin barrier have become crucial in the prevention and management of AD. 15 In this study, we demonstrated that oral GMP administration prevents skin barrier dysfunction by upregulating filaggrin and AMP expression, and by strongly reducing epidermal thickness and IFN-γ expression. Besides, GMP administration impacts the skin microbiota, by diminishing S. aureus colonization in injured skin without changing the S. epidermidis level. These beneficial effects are accompanied by the enhanced growth of Bifidobacterium and an increase in the fecal SCFA levels.
The amounts of filaggrin, involucrin, BD-2, and CRAMP in injured tissues were lower in our AD-like rats than in control animals, as previously reported in experimental models and in patients with AD. 5,27 During AD, the generated Th2 milieu has been proposed as the mechanism underlying the low expression levels of epidermal structural proteins and AMPs. 27 –29 It is known that GMP downregulates IL-4 and IL-13 expression in AD lesion 14 ; however, GMP administration increased filaggrin, BD-2, and CRAMP expression, but not involucrin expression. Thus, we consider that other factors, in addition to Th2 cytokines, might be involved in involucrin downregulation.
GMP might differentially modulate the factors participating in epidermal structural protein expression, as STAT3 is involved in the downregulation of filaggrin expression, 30 while STAT6 is implicated in involucrin expression. 31 In traditional Iranian medicine, one of the most common treatments for severe and chronic AD is the intake of milk whey with an aqueous extract of field dodder seeds, with demonstrated efficacy on skin moisture and elasticity in a clinical trial. 32 Our results are in agreement with these findings, as the oral administration of GMP, one of the main components of whey, increases filaggrin expression, a protein that is a natural skin moisturizing factor.
Thickening of the epidermis in patients with AD is due to changes in epidermal proliferation and differentiation. 2,33 In our experimental model, epidermal thickening was evident, and GMP administration before or after AD induction significantly reduced it. IFN-γ is involved in the hypertrophy of the skin in AD. 22 Therefore, the reduction in epidermal thickness is in agreement with the low IFN-γ mRNA expression in GMP-treated animals. Negative regulatory effects of GMP on IFN-γ production have been reported in other disease experimental models, such as colitis or phenylketonuria. 34 –36
Normal healthy skin is colonized by S. epidermidis, S. haemolyticus, and S. hominis. 37 These organisms compete for nutrients and space, thereby preventing the colonization of opportunistic pathogens, such as S. aureus, which arises in ∼5% of the healthy population and is greatly increased in patients with AD. More than 90% of AD patients have S. aureus colonization on their lesional and, to a lesser extent, nonlesional skin. 38
In our animal model, rats with AD showed an increase in S. aureus colonization in lesional skin, although the level of S. aureus recovered to control levels with GMP treatment; this finding is in accordance with the upregulation of filaggrin and AMPs, as the absence of filaggrin in keratinocytes is associated with epidermal S. aureus colonization, 39 and AMPs have anti-staphylococcal activity. 40,41 In addition, the Th2 downregulatory effect reported for GMP in AD skin lesions 14 may also be involved in the reduction in the amount of S. aureus, because IL-4 plays a crucial role in staphylococcal binding to skin by inducing the synthesis of fibronectin by skin fibroblasts, which allows S. aureus adherence. 42,43
AD development has been associated with alterations in the normal microbiota composition. In children with AD, some genera such as Bifidobacterium, Staphylococcus, Corynebacterium, and Bulleidia are completely absent from their feces. 44 Moreover, Bifidobacterium is decreased in fecal samples of adult patients with AD, and its abundance is significantly diminished in patients with severe skin symptoms. 45 In accordance with these findings, supplementation with Bifidobacterium and Lactobacillus species prevents the development of eczema and skin inflammation in humans and mice. 46,47 Because GMP intake mainly and strongly increased Bifidobacterium in our AD model, we suggest that GMP may exert a protective role in AD, at least in part, by its prebiotic action.
Bifidobacterium is an SCFA-producing genus of bacteria, 48 and a high intestinal abundance of butyrate-producing bacteria has been associated with a lower severity of cutaneous signs and symptoms in infants with atopic eczema. 49 We demonstrated that prophylactic GMP administration significantly increased acetate and butyrate in feces, indicating that these SCFAs may be involved in its effect. This result parellels previous observations that show an increased SCFA production in mice fed a GMP-enriched diet or by in vitro fermentation of GMP with fecal microbiota. 36,50 It is known that a portion of SCFAs absorbed by enterocytes reaches systemic circulation, 51,52 and thus, the preventive effect of GMP on a dysfunctional cutaneous barrier might be mediated by the direct action of acetic acid and butyric acid on skin cells. 53 –57
Moreover, an indirect effect of these SCFAs on AD-lesional skin through immune or neuroendocrine system regulation should not be disregarded. Strikingly, we did not observe any change in the fecal SCFA content when GMP was administered in a therapeutic manner. Longer schedules of treatment are likely needed to induce a detectable increase in SCFAs. It is important to highlight that upon GMP ingestion, some peptides with biological activities can be released. 58 In this context, systemic GMP or GMP-derived peptides may also directly modulate the activity of some cells, as demonstrated in immune cells. 12 Currently, our research focused on studying the direct effect of GMP, or products derived from its enzymatic digestion, on keratinocytes.
In conclusion, we found that the oral administration of GMP increases filaggrin, BD-2, and CRAMP expression in rat lesional like AD skin, leading to a decrease in S. aureus colonization. It also strongly inhibits skin thickening and IFN-γ production. The increase in the intestinal amount of Bifidobacterium by GMP intake may represent one of the mechanisms of action of the protective effect of GMP on AD. Thus, orally administered GMP is a good candidate for the prevention and management of an altered epidermal barrier in AD. Beneficial effects of GMP on damaged skin might prevent the development of other allergic pathologies, as AD is considered the beginning of the atopic march. However, further studies are needed to understand the full mechanisms underlying the protective effect of GMP and also its metabolism and tissue distribution.
Finally, this study provides new perspectives to analyze the potential therapeutic effect of GMP in other skin diseases with impaired barrier function.
Footnotes
Acknowledgments
Authors wish to thank Manuel Tinajero for excellent technical assistance and MVZ José Luis Ponce and MVZ Jesús Humberto Estrada for providing the animals for the study.
Author Disclorure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Grant PIBB 15–9N from the Autonomous University of Aguascalientes and the Grant 240921 from CONACYT.