
Abstract
Hepatic fibrosis emerges upon exposure of liver to various chemicals and if not treated, it develops various diseases such as cirrhosis and cancer. Carbon tetrachloride (CCl4) is a widely used toxin in animal models to develop hepatic fibrosis. Accumulation of unfolded proteins in cells causes stress in the endoplasmic reticulum (ER) and various mechanisms are involved in the cell to reduce the damage caused by these unfolding proteins. The most well known of these is the unfolded protein response. Further, autophagy works to remove these proteins if the damage cannot be repaired and is permanent. In our study, we investigated the effects of naringenin (NRG), a flavanon abundant in citrus fruits, on ER stress and autophagy in CCl4-injured rat liver. The animals were given 0.2 mL/kg of CCl4 for 10 days and treatment group was administered 100 mg/kg of NRG for 14 days. Histopathological examination was performed to show liver damage and to determine the therapeutic properties of the active substance. Transmission electron microscopy (TEM) analysis was carried out to establish cell level damage and effect of treatment. In addition, levels of ER stress and autophagy markers of liver were measured. According to our findings, TEM demonstrated positive effect of NRG and histological examinations reported ameliorative effects. In addition, NRG reduced levels of ER stress markers and inhibited autophagy significantly compared to CCl4-treated group. As a result, NRG significantly reduced damage in hepatocytes and provided a significant amelioration.
Introduction
The prevalence of chronic liver diseases, which lead to serious impairment of liver function, increases in young and old populations due to exposure to viruses, drugs, and toxins. 1 Hepatic fibrosis is one of the most important disease related to liver toxicity and reversible character of it would help development of new antifibrotic therapeutic approaches. Candidate agents should be investigated for having any antifibrotic effects so that it can prevent, stop, or reverse liver fibrosis, but still there is a lack of effective clinical therapies. 2
Carbon tetrachloride (CCl4) metabolism catalyzed by liver microsomal cytochrome P450 rapidly produces free radicals that consume hepatic glutathione and induce chain reaction of lipid peroxidation of hepatocyte membrane. 3 This CCl4 metabolism in endoplasmic reticulum (ER) leads to an unfolded protein response (UPR) as an indicator of loss of ER function. This defect triggers ER stress and it has been shown in many liver diseases. It has been observed that the UPR occurs in fatty liver, hepatic insulin resistance in obesity, ischemia-reperfusion injury, hepatitis B and C, alcohol-induced liver damage, and many toxic agent-borne diseases including acetaminophen toxicity. 4
Immunoglobulin heavy-chain-binding protein (BIP)/glucose-regulating protein 78 (GRP78) detects misfolded protein accumulation in ER and hence triggers UPR, which is a process that increases protein folding capacity, chaperone production, and also slows down entering of new proteins into ER. ER stress-related proteins inositol requiring enzyme 1 (IRE1), pancreatic ER kinase (PERK), and activating transcription factor 6 (ATF6) are activated upon chaperone BIP production by unfolded protein depositon in ER. Phosphorylation of initiation factor 2 alpha (eIF2-α) by PERK causes general mRNA translation weakening in the cell. At the same time, ATF4 boosts of numerous mRNA expressions such as antioxidant enzymes, chaperones, and apoptosis-related genes including CCAAT-enhancer-binding protein homologous protein (CHOP). IRE1 autophosphorylation is activated for the production of active transcription factor spliced X-box binding protein 1 (XBP1s). Cleavage of ATF6 in Golgi results with transformation of its active form and transfer to nucleus. All of these are adaptations to solve protein folding defects and increase cell survival. If stress becomes constant, this adaptation will fail and lead to deterioration of ER functions. All this eventually triggers apoptosis by causing to activate proapoptotic transcription factor CHOP, downstream target of UPR, and produce reactive oxygen species. 2,4,5
Autophagy is a highly conserved cellular recycling process that degrades damaged organelles and macromolecules and serves to protect cellular homeostasis. This process is based on the principle of transferring these macromolecules or damaged organelles to lysosome with double-membrane-bound vesicle called autophagosome. 6,7 Autophagosome formation is initiated by phosphoinositide 3 kinase (PtdIns3K), vacuolar protein sorting 34 (Vps34), and Beclin-1 and this process is controlled by protein kinase ULK1, autophagy-related proteins Atg13 and Atg17. Phosphatidylinositol 3-phosphate (PtdIns3P), formed by Beclin1/Vps34, is required to bring other Atg proteins necessary for maintenance of autophagosome structure to the site. The initiation phase is followed by formation of Atg5–Atg12–Atg16 complex, which leads to conversion of microtubule-associated protein 1 light chain 3-I (LC3-I) to LC3-II and LC3-II attaches to the membrane by activation of Atg7 and Atg3. 2,8 In addition, UPR tries to overcome the harm caused by misfolded proteins and organelles by stimulating autophagic destruction. 9 Apoptosis is triggered if UPR-mediated autophagy fails to provide homeostasis in the cell, 10 so fate of an ER stressed cell depends on balance between UPR and autophagy. 11
Recent studies have shown that autophagy and ER stress significantly increase in CCl4-induced liver injury and fibrosis. 2 It has been shown that inhibition of autophagy and reduction of autophagy-related protein expression such as Atg7 may prevent formation of fibrogenesis. 12
Flavonoids, which are commonly found in vegetables and fruits, may prevent the damage caused by free radicals in many ways mostly by scavenging superoxide or peroxynitrite. 13 Naringenin (NRG, 4, 5, 7-trihydroxy-flavonone), a bio-flavonoid commonly found in citrus fruits, tomatoes, and cocoa, has a strong radical scavenging effect. Several studies have reported a slightly protective effect of NRG against increased hepatic lipid peroxidation and oxidative stress in rats exposed to several toxic agents. 14 –17 In addition, many studies have shown that NRG has anti-inflammatory effects against CCl4-induced toxicity. 18 Besides, it has been shown to play an effective role in the liver by increasing expression of tumor suppressors against hepatocellular carcinoma. 19 Furthermore, it was found to be effective against hepatitis C by decreasing viral protein synthesis in liver. 20 However, effects of NRG on CCl4-induced ER stress and autophagy as a result of this stress in hepatocytes have not been elucidated yet.
Materials and Methods
Animals and study design
This study was carried out in accordance with Eskisehir Osmangazi University Faculty of Medicine Ethics Committee (approval no: 111, meeting no: 600 on May 23, 2017) and recommendation of Guide for the Care and Use of Laboratory Animals of the National Institutes of Health were followed at all stages. Female Wistar albino rats weighing 250–300 g were used in this study. Rats were housed in a room of Eskisehir Osmangazi University Medical Faculty Medical Surgery Research Center (TICAM) with an average temperature of 22 ± 2°C and a humidity of 45–50% with 12/12 h light/dark conditions during the experiment and fed with standard laboratory chow and tap water ad libitum. After a week of acclimatization five experimental groups were formed in each group to include seven rats (Table 1).
The Substances Applied to Groups
CCl4, carbon tetrachloride; SD, standard diet; i.p., intraperitoneal; NRG, naringenin; i.g., intragastric.
Carbon tetrachloride ([0.2 mL/kg] Sigma-Aldrich; Product code: 289116, density:1.594 g/mL) was dissolved in olive oil (1:1) and administered peritoneally (intraperitoneal [i.p.]) for 10 days, twice a day. Following CCl4 administration, 100 mg/kg NRG (Sigma-Aldrich; Product code: N5893) dissolved in water was applied to the treatment group for 14 days intragastrically. Olive oil (0.2 mL/kg) and NRG (100 mg/kg) alone administrated groups were determined as sham control group. After last treatment, animals were sacrificed under anesthesia with ketamine/xylazine (90/10 mg/kg, intramuscular [i.m.], i.p.) and liver tissues were collected.
Transmission electron microscopy
For transmission electron microscopy (TEM) liver tissues were placed in primary fixative in 2.5% glutaraldehyde containing 0.1 M phosphate buffer at 4°C for 24 h. Secondary fixations were achieved in 1% osmium tetroxide containing 0.1 M phosphate buffer at room temperature in rotator for 2 h, then tissues were washed three more times with phosphate buffer. Samples were treated with increasing ethyl alcohol gradient series (30%, 50%, 70%, 90%, 96%, and 100%) twice for dehydration at 4°C and were kept in propylene oxide for 30 min (twice). After standing in rotator for 2 h in a propylene oxide-araldite mixture at a ratio of 1:1, tissues were taken into pure araldite and were kept in the rotator overnight. Next day, they were embedded in araldite and polymerized at 60°C for 48 h. Ultra thin sections (60 nm) were obtained from blocks by ultramicrotome (Leica Ultracut R). These sections were taken to 300 mesh copper grids, stained with uranyl acetate-lead citrate, and then examined by TEM (JEOL JEM 1220).
Histopathological examination
The liver samples were fixed in 10% formalin for 2 days. Tissues were then dehydrated gradually in 70%, 80%, 90%, and 96% alcohol series respectively for 45 min. After dehydration, samples were kept in xylene for 20 min. Tissues were paraffinated at 65°C and 5 μm sections were taken from paraffin blocks with a microtome. Samples were treated with xylol for 1 h for deparaffinization and sections were stained with hematoxylin–eosin. Then, sections were dehydrated with alcohol again and were kept in xylol twice for 30 min. After they were coated with entellan histological evaluations were performed with Olympus BH-2 light microscope.
Determination of autophagy and endoplasmic stress markers
For the preparation of homogenates, liver tissue samples were weighed and washed in phosphate-buffered saline (pH 7.4); then, homogenized in ultrasonic homogenizer according to the manufacturer's instructions (Shangai YL Biotech Co., Ltd.). Total amounts of protein in tissues were calculated using Sigma, Total Protein Kit (Product code: TP0100). The markers of ER stress: CHOP (Cat no: YLA1471RA), GRP78 (Cat no: YLA0246RA), ATF6 (Cat no: YLA1403RA), ATF4 (Cat no: YLA1404RA), IRE1 (Cat no: YLA1405RA), PERK (Cat no: YLA1406RA), and XBP1s (Cat no: YLA1407RA) and autophagy markers Beclin1 (Cat no: YLA0731RA), Vps34 (YLA1420RA), Atg5 (YLA1418RA), Atg7 (YLA1419RA), and ve LC3-II (YLA1417RA) were detected in accordance with Shanghai YL Biotech Co., Ltd. ELISA kit protocols.
Statistical analysis
Data analysis was performed with using IBM SPSS 21 program. Descriptive statistics of the variables were shown as mean ± standard deviation. The normality of variables was assessed by Shapiro–Wilk test. For the normally distributed data, comparisons of the groups were done with one-way analysis of variance (ANOVA) analysis. For significant results of ANOVA, multiple comparisons were evaluated with Bonferroni test. P < .05 was considered as statistically significant.
Results
TEM demonstrated positive effect of NRG
According to TEM investigations, normal hepatocyte structure was observed in control group samples, hepatocyte nuclei were round or oval shaped, chromatin was also homogeneous and euchromatic. Mitochondria and ER were in a complete integrity and in normal appearance. In addition, rough ER structures appeared to be organized and abundant. Normally shaped homogeneous nuclei were observed in NRG group. ER structures generally appeared similar to control having healthy morphology. No dense vacuolizations were detected. The most significant finding in the CCl4 group was observation of advanced vacuolizations containing cell debris. These autophagic vacuoles appeared in large quantities, especially near mitochondria. Number of ER decreased considerably, fractures and dilations were also detected in a small number of ER. The most obvious finding observed in mitochondria is that electron density was very high. Although mitochondria did not show any loss of crista, atypical appearance and deterioration in double membrane were detected. In addition, chromatin condensation was found at periphery of nuclei. In the treatment group (CCl4 + NRG), nuclei were normal, homogeneously distributed, and regular membrane structure was determined. ER and granular ER structures were generally in a healthy morphology and ER fractures and swelling were much less identified. In this group, less vacuolization were observed compared to the CCl4-treated group, mitochondria and ER damage also were negligible (Fig. 1).
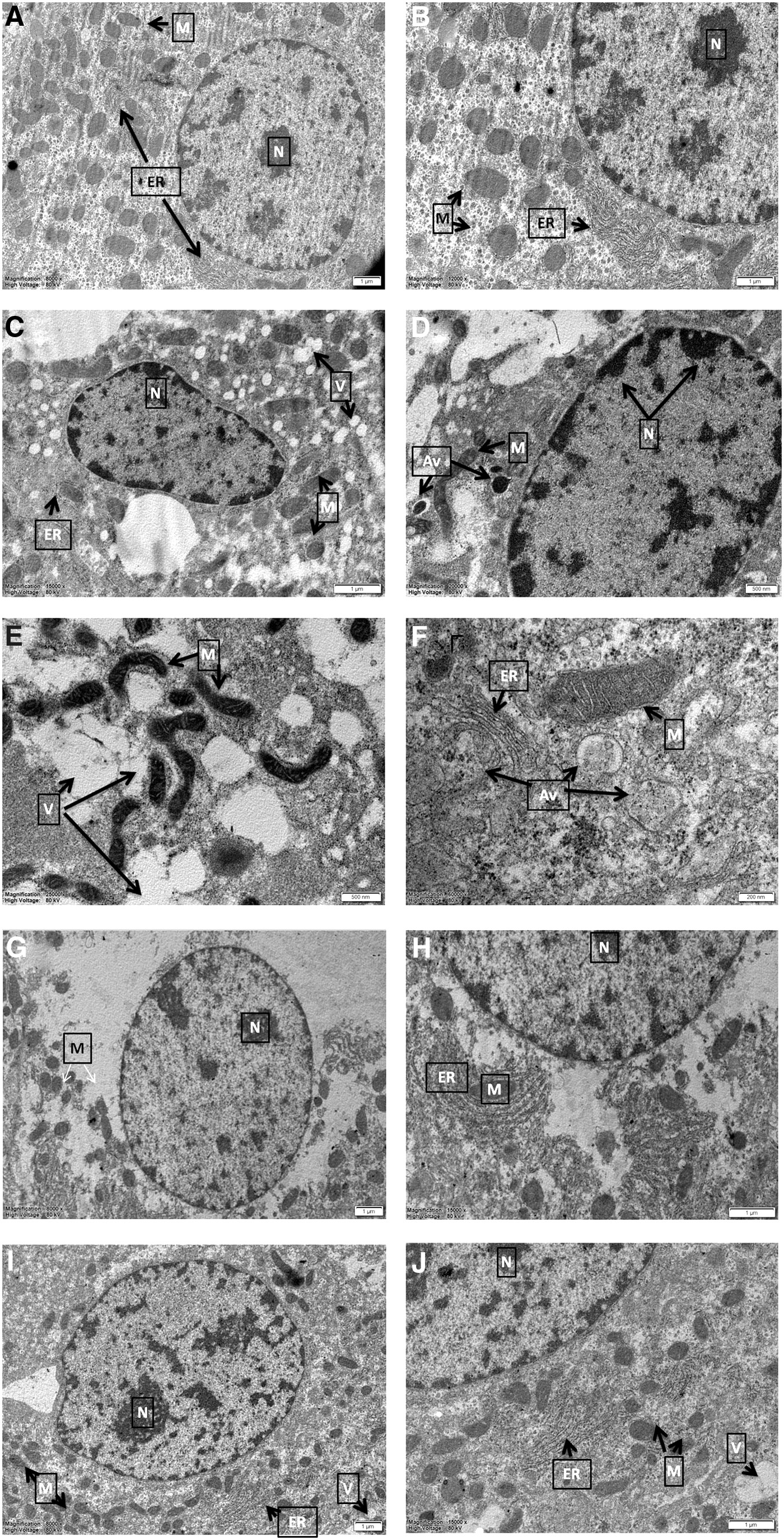
Transmission electron microscope detection of hepatocytes. Normally shaped and euchromatic nucleus (N) were detected in the control group. Mitochondria (M) and ER were in their normal structure and showed complete membrane integrity
Histological examinations reported ameliorative effect of NRG
Histological examinations of the groups were given in Figure 2 and dense vacuolization especially in pericentral regions, cellular degeneration, congestion in central vein, and partial cellular infiltration were observed in liver of rats in the CCL4 group. Although damage to pericentral regions of livers in CCL4 + NRG group appeared to be minimal, cellular damage and vacuolization were decreased compared to the CCL4 group. It was also detected that congestion of central vein continues. Other groups were found to have livers in normal histological structures.
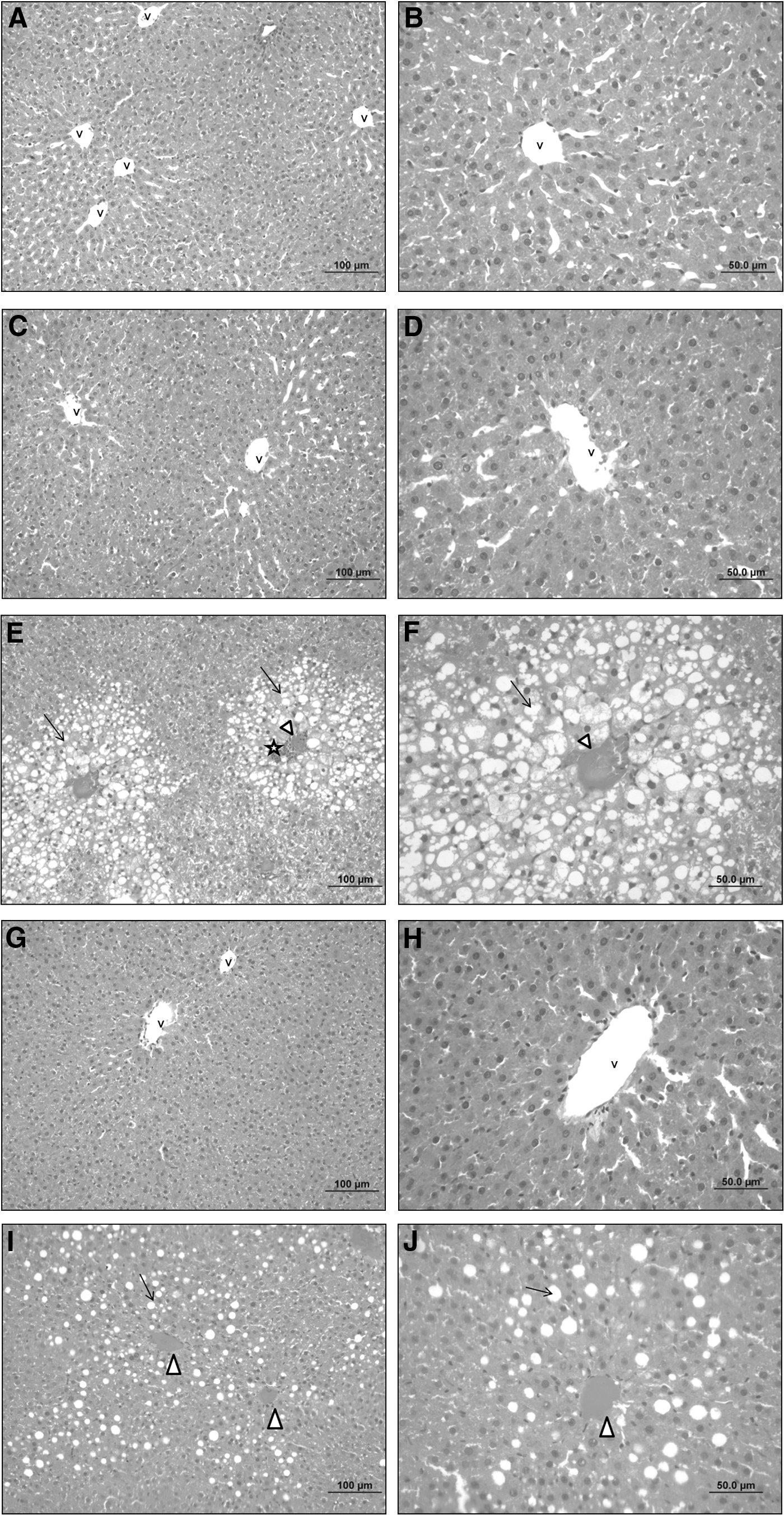
Histological examinations of liver tissues. Normal histological liver structure of Control
NRG reduced levels of ER stress markers and inhibited autophagy
Hepatocurative effect of NRG against CCl4-mediated ER stress was shown in Table 2. According to the results, CCl4 application significantly increased protein levels of ER stress markers CHOP, GRP78, ATF6, ATF4, IRE1, PERK, and XBP1s compared to the other groups in liver (P < .001). NRG treatment (CCl4 + NRG) was highly effective on these markers and downstream targets responsible for UPR and reversed CCl4-induced ER stress significantly (P < .001). In addition, a significant difference in IRE1 levels was found between the control group and NRG group (P < .01). Moreover, XBP1s protein levels were significantly decreased in NRG group compared to the control (P < .05). Furthermore, no difference was found between control and olive oil groups in all UPR markers.
Effects of Naringenin on Endoplasmic Reticulum Stress Markers
Data are presented as mean ± SD (n = 7).
Significant differences were found: * P < .05, ** P < .01, *** P < .001 compared with control group; ### P < .001 compared with CCl4 group.
CHOP, CCAAT-enhancer-binding protein homologous protein; GRP78, glucose-regulating protein 78; ATF6, activating transcription factor 6; ATF4, activating transcription factor 4; IRE1, inositol requiring enzyme 1; PERK, pancreatic ER kinase; XBP1s, spliced X-box binding protein 1; SD, standard deviation.
Results presented in Table 3 showed that autophagy-associated protein levels were significantly increased in CCl4 group (P < .001), supporting ER stress marker changes. Significant decreases were found in all marker levels in CCl4 + NRG group compared to CCl4 group (P < .001). There were no significant differences in autophagy marker levels between control and olive oil and between control and NRG groups.
Effects of Naringenin on Autophagy Markers
Data are presented as mean ± SD (n = 7).
Significant differences were found: *** P < .001 compared with control group; ### P < .001 compared with CCl4 group.
ATG5, autophagy-related protein 5; ATG7, autophagy-related protein 7; LC3-II, microtubule-associated protein 1 light chain 3-II; VPS34, vacuolar protein sorting 34.
Compatible with TEM analysis and histological examinations critical abrogation of these markers was also reported with NRG treatment (P < .001).
Discussion
Liver diseases are crucial problems in the world and many environmental toxins cause liver damage and development of these diseases. 21 Despite recent advances in hepatology, treatment methods used in liver diseases are insufficient to solve toxin-borne pathologies. Elimination of toxicity in liver, which is the most important organ affected by toxic substances at cellular level, will be beneficial for liver diseases. 22,23
CCl4 is bioactivated to trichlromethyl free radical (CCl3) by cytochrome P450 enzymes in hepatocytes, of which P450 2E1 isoform found in liver ER has been shown to be a crucial enzyme for this bioactivation and also for CCl4-caused pathological changes. Cytochrome P450 enzyme system requires an electron for its activation that is transferred between NADPH-P450 reductase and P450. A leak of electron from the transfer system occurs during the transfer that results in production of free radicals from redox cycle. 24,25 Many studies have reported that these free radicals cause ER stress. 2,26 In a study investigating whether ER stress was induced by CCl4, similar results were observed in tunicamycin-triggered ER stress, known to be an N-acetyl-glycosylation inhibitor. 26
Inflammation and cell death pathways that work against liver damage stimulate hepatocytes for liver regeneration. However, in cases of persistent and severe toxicity, hepatic stellate cells undergo myofibroblast-like activation and synthesize more extracellular matrix proteins. 27,28 In this case, the need for advanced protein synthesis and hence increase in the protein folding process requires excessive work of ER. 2,29 Although there are not many studies on the mechanism of the relationship between ER stress and autophagy in stellate cells, one study reported that blocking of IRE1 inhibits the activation of autophagy and prevents the formation of fibrosis. 30 Recently, CCl4 has been shown to cause swelling of ER in hepatocytes, providing increased expression of GRP78 and XBP1, a remarkable fact of ER stress. 31 Three ER-transmembrane proteins: IRE1, PERK, and ATF6 have been described as ER stress key sensors. UPR, especially triggered by secretory proteins, attenuates translation by PERK activation followed by phosphorylation of eIF2-α to reduce accumulation of proteins in ER. Furthermore, in response to ER stress, cells expanded protein folding ability by upregulating ER chaperones. 26,31,32
The therapeutic potential of natural herbal products has given plants an important role in using them in the fight against various diseases for centuries. 33 Many potential antioxidant composites were tested for protective activity against CCl4 toxicity. 34,35 NRG has prevented free radical accumulation caused by cytochrome P450 enzymes and has demonstrated curative effect against CCl4-induced hepatotoxicity in rat liver. 36 It was also found to be effective during hepatic inflammation by regulating Nrf-2 pathway. 37 In addition to these properties of NRG, which was shown to increase p38 tumor suppressor expression in breast cancer cells, 38 it may also be considered that it is an active agent to activate the UPR and to pull the trigger to overcome stress in ER. Therefore, in our study, we have hypothesized that NRG can be used to alleviate ER stress and autophagy at cellular and molecular levels against hepatic damage.
Previous studies have indicated that CCl4 toxicity causes significant damage to liver tissue such as tissue infiltration, regional necrosis, and degeneration. 13,35 In a study by San-Miguel et al., toxic effects of CCl4 were detected in TEM examinations in mouse hepatocytes and increased vacuolization, damaged organelles, and autolysosomes containing damaged organelles were reported. 2 Many histological examinations and TEM analysis demonstrated that NRG contributes greatly to elimination of damage to these tissues and repair of cellular damage caused by toxicity. 13,37 In our study, in accordance with previous studies, CCl4 toxicity caused a significant decrease in ER number at cellular level, swelling of ER, and formation of fractures in ER membrane. In addition, atypical appearances in mitochondria and disturbances in membrane structure indicated malfunction in energy metabolism. Besides, focal necrotic areas around nucleus were also noteworthy. Compatible with study of San-Miguel et al. intensive vacuolizations and autophagic vacuoles were noticed in CCl4 group. 2 Both histological examinations and TEM revealed that NRG was highly effective at eliminating liver damage induced by CCl4 at tissue and cellular levels. Organelle losses, degeneration in nucleus morphology, and vacuolization were largely eliminated at cellular level and degeneration and infiltration in tissues were prevented.
In an in vitro study of HepG2 cells, response of resveratrol treatment to CCl4-induced apoptosis and ER stress was investigated. A significant increase in mRNA expression levels of XBP-1s, CHOP, IRE1, ATF6, ATF4, eIF2α, PERK, and protein levels of GRP78, IRE1, and PERK were detected in cells treated with CCl4 and in resveratrol treated cells a significant reduction in expression of these key proteins were reported. 39 In another study, effects of melatonin treatment on ER stress and autophagy in CCl4-induced hepatic fibrosis model were researched. PERK, ATF6, and IRE1 protein expressions were significantly increased in fibrosis group and melatonin administration showed a significant decrease in levels of these markers. 2 According to our findings, there was a significant increase in protein levels of ER stress markers in liver in the CCl4 group compared to control. According to our findings, CCl4 caused ER stress in hepatocytes and formed a UPR that could not be overcome. Supporting our hypothesis, treatment of NRG in rats with CCl4-induced toxicity was very successful in preventing triggered ER stress and provided a downregulation of stress markers.
The formation of ER stress stimulates the cells to try to overcome stress by activating various signaling pathways. One of these signaling pathways induces transcription of autophagy genes and establishes relationship between UPR and autophagy. 40 It has been determined that autophagy, which is shown to be highly effective in liver fibrogenesis, was stimulated by the IRE1α-Xbp1 pathway and contributes to cell homeostasis. 30 In addition, knockdown of BIP, one of the key factors responsible for protein folding, was found to inhibit autophagosome formation in mammalian cells. 41 Furthermore, PERK-mediated phosphorylation of eIF2-α triggered by unfolded proteins was shown to induce formation of Atg5–Atg12–Atg16 complex and LC3 transformation. 42 Several studies have demonstrated that CCl4 derived liver fibrosis triggered UPR-mediated autophagy. 2,43 Similar to other studies, in our study, there was a significant increase in Vps34 and Beclin-1 levels, which are leading proteins in initiation of autophagic vacuole formation and LC3-II, a key marker for conjugation step, and Atg5 and Atg7 in CCl4 group. Recent researches have reported that inhibition of autophagy was very effective in reversing damage in hepatocytes. 2,44 In addition, a study examining the hepatoprotective effect of betaine in nonalcoholic fatty liver disease-induced mice was reported to increase the expression of autophagy activators along with antioxidant and anti-inflammatory properties of the natural compound. 45 In the present study, in agreement with TEM investigations, NRG prevented progression of autophagy by alleviating damage to organelles and modulating ER stress.
In addition to regulating many metabolisms such as fat, protein, and carbohydrate metabolism, liver plays a crucial role in detoxification of toxic materials entering organism. Because of its numerous functions, various metabolic, infective, circulatory, neoplastic, and toxic disorders can affect liver. ER stress and UPR-mediated autophagy occur in many acute and chronic liver diseases. Moreover, considering that a more severe and chronic administration of hepatotoxic (CCl4, lipopolysaccharide etc.) agents may cause inhibition of autophagy, studies are needed to determine the dosages and durations of toxic agents that will cause a permanent hepatic damage (or a condition in which cell death pathways is induced). It is important to suppress these protein folding defects and cellular mechanisms that cause these defects (such as free radical accumulation and inflammatory pathways) for therapy. The results obtained from the study are important in terms of the use of new natural active substances in treatment and contributing to future studies.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by the Commission of Scientific Research Project Support Bilimsel Araştırma Projeleri Komisyonu (BAP) Eskisehir Osmangazi University (Turkey).