
Abstract
Histone acetyltransferase (HAT) activity is well established to regulate inflammatory responses. In contrast, the mechanisms by which natural nutritional extracts influence epigenetic mechanisms to regulate inflammation have not yet been thoroughly investigated. Thus, in the present study, we observed that the anti-HAT activity exerted by an ethanol extract of Ligularia fischeri (ELF) inhibited inflammation. Specifically, we used a cell-free system to show that ELF attenuates HAT activity. We also demonstrated that ELF decreases lipopolysaccharide (LPS)-induced HAT mRNA and protein expression levels in Raw 264.7 cells, and thereby attenuates inflammation-induced patterns of hyperacetylation at nonhistone and histone-H4 proteins. Interestingly, we found that ELF blocked p65 translocation in LPS-stimulated Raw 264.7 cells by attenuating acetylation at lysine residue 310 of p65. Finally, we investigated whether ELF reduces the inflammatory cytokines, IL-6, IL-1β, and TNFα, using its HAT inhibitor activity. Taken together, these results suggest that ELF negatively regulates inflammatory responses by inhibiting HATs and HAT activity.
Introduction
Inflammation is a major biological process that is affected, mediated by both the organism itself and environmental and dietary stimuli, 1 causing many infectious and noninfectious diseases. 2 Indeed, nutrition affects most chronic inflammatory diseases, including obesity and diabetes, 3 demonstrating that nutrients are able to modulate inflammation. 4,5 Thus, food has recently emerged as an important factor in inflammation-related diseases.
Protein acetylation, which is regulated by histone acetyltransferases (HATs) and histone deacetylases (HDACs), influences a broad set of cellular processes implicated in human diseases, including inflammatory diseases. 6,7 The acetylation of histone has been reported to be associated with the transcription of inflammation-related genes. 8 For example, corticosteroids recruit HDAC2 to reverse histone acetylation patterns and thereby suppress the transcription of multiple inflammatory genes. 9,10 Furthermore, the acetylation of nonhistone proteins has also been shown to mediate inflammation by regulating mRNA stability, protein localization and degradation, and protein/protein interactions of various target genes and proteins. 6,11
The ubiquitously expressed nuclear factor-κB (NF-κB) is required for, and critically regulates, inflammation. 12 Following the engagement of a Toll-like or cytokine receptor by lipopolysaccharide (LPS) or tumor necrosis factor α (TNF-α), respectively, IkappaB kinase (IKK) is phosphorylated. An activated IKK then phosphorylates serine residues 23 and 19 of inhibitory kappaB (IκB) 13 and thus promotes IκB polyubiquitination and degradation. 14 Consequently, NF-κB, which is constitutively bound to IκB in the cytosol, enters the nucleus and activates the transcription of genes that encode various cytokines such as interleukin (IL)-1β, IL-6, and TNF-α. 15,16 The most abundant form of NF-κB is a heterodimer of p50 and p65. 17 Notably, p65 has been shown to be a key transcriptional activator. 18 Chen et al. showed that the p65 lysine (K) residues 218, 221, and 310 are acetylated by p300 and HAT, 19 and the acetylation of K310 modulates the nuclear translocation of cytosolic NF-κB. 20 Thus, regulating p65 acetylation is essential to treat inflammatory diseases.
Ligularia fischeri (LF) belongs to the Compositae family of perennial vegetable plants that are distributed throughout Europe and Asia. 21 LF leaves have been used to treat inflammatory diseases such as diabetes mellitus and rheumatoid arthritis. 22 Many investigators have demonstrated that LF exerts antiobesity, 23 antihepatotoxic, 24 and anti-inflammatory effects. 25 Furthermore, the LF derivative has been reported to reduce inflammation by suppressing COX-2. 26
However, while the beneficial anti-inflammatory effects of LF are well established, the epigenetic mechanisms that underlie them have not been elucidated. Thus, the present study demonstrated that LF abrogates nuclear NF-κB translocation by exerting anti-HAT activity and thereby inhibits inflammation.
Materials and Methods
Cell culture
Raw 264.7 cells were purchased from the American Type Culture Collection (Manassas, VA, USA). Cells were cultured using high-glucose Dulbecco's modified Eagle's medium (DMEM). Cells were pretreated with 70% ethanol extract of LF (ELF) for 18 h, and 1 ng of LPS (Sigma, St. Louis, MO, USA) was added for 6 h before cell harvest.
Preparation of LF extract
LF was purchased from a local market (Seongnam, Republic of Korea) and extracted in a 10-fold volume of 70% ethanol, and the precipitate was removed by centrifugation at 8000 × g for 30 min. The supernatants were then freeze dried.
Cell toxicity
After reaching ∼70% confluence, cells were treated with ELF at 10, 25, 50, or 100 μg/mL. After a 24-h incubation, cell toxicity was measured by WST-1 assay (Enzo Life Sciences, Inc., Farmingdale., NY, USA).
Quantitative real-time polymerase chain reaction
Total RNA was isolated using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using an I Cycler iQ system (Bio-Rad, Hercules, CA, USA). PCR amplification was carried out using the following primers: for mouse IL-6, forward 5′-CCT CTG GTC TTC TGG AGT ACC-3′, reverse 5′-ACT CCT TCT GTG ACT CCA GC-3′; for mouse IL-1β, forward 5′-; for mouse TNFα, forward 5′-ATG AGC ACA GAA AGC ATG A-3′, reverse 5′-AGT AGA CAG AAG AGC GTG GT-3′; for mouse p300, forward 5′-GTT GCT ATG GGA AAC AGT TAT GC-3′, reverse 5′-TGT AGT TTG AGG TTG GGA AGG-3′; for mouse CBP, forward 5′-GAC CGC TTT GTT TAT ACC TGC-3′, reverse 5′-TCT TAT GGG TGT GGC TCT TTG-3′; for mouse PCAF, forward 5′-TGT CAT TGG TGG TAT CTG T-3′, reverse 5′-ATA TGT GAG GAA GTT GAG GAT-3′, and for mouse βActin, forward 5′-CTA AGG CCA ACC GTG AAA G-3′, reverse, 5′-ACC AGA GGC ATA CAG GGA CA-3′.
Immunoblot assay
Following treatment, the cells were lysed with radioimmunoprecipitation assay buffer (RIPA) (Elpis, Daejeon, Republic of Korea). Total protein was measured using the bicinchoninic acid assay (BCA) method and loaded on 10% Bis/Tris gels for electrophoresis. The membranes were blocked for 1 h and incubated with primary antibodies against histone H3K9, histone H4K8Ac, histone H4K12Ac, histone H4K15 (Cell Signaling, Danvers, MA, USA), and β-actin (Enzo Life Sciences, Inc.) overnight. After washing, membranes were incubated with secondary antibodies for 1 h. After washing, the protein bands were detected by chemiluminescence.
Immunofluorescence assay
Cells, 1 × 104, were cultured on coverslips and treated with LPS with or without ELF at an indicated concentration for 2 h. Cells were fixed in 100% methanol for 30 min and then treated with 0.25% Triton X-100 in phosphate-buffered saline for 5 min at room temperature. For detection of endogenous p65, cells were incubated with the anti-p65 antibody and then stained with the goat anti-rabbit fluorescein isothiocyanate (FITC; Invitrogen) at 37°C for 1.5 h. Nucleus was revealed with Hoechst 33258 (Invitrogen) staining. Cells were imaged using a confocal microscope.
Nuclear fraction
Cells were lysed in lysis buffer [10 mM Tris (pH 7.4), 10 mM NaCl, 3 mM MgCl2, 0.5% NP-40, and protease inhibitor cocktail (Roche)]. The supernatants were used as cytosolic fractions and the pellets were used as nuclear fractions.
Histone extraction
Cells were lysed according to the manufacturer's protocol (Abcam, Cambridge, MA, USA). The cells were lysed with a prelysis buffer to isolate the nuclear fraction. The nuclear pellet was resuspended in lysis buffer, incubated for 30 min on ice, and centrifuged at 9,600 × g for 5 min at 4°C. The supernatant was transferred to a clean tube and Balance-Dithiothreitol buffer was added.
In vitro HAT assay
HeLa cell nuclear extracts were assessed for HAT activity according to the manufacturer's protocol (BioVision, Inc., Militas, CA, USA). IC50 values were calculated using Prism 5 software (version 5.03; GraphPad Software, Inc., CA, USA).
Statistical analyses
Data are expressed as mean ± standard deviation (SD). The statistical analyses were conducted using SPSS (ver. 20; SPSS, Inc., Chicago, IL, USA). Statistical differences between mean values were evaluated by either Student's t-test or one-way analysis of variance (ANOVA) followed by Bonferroni post hoc test. A value of P < .05 was considered to be statistically significant.
Results
ELF is a specific inhibitor of HATs
While screening a library of natural substances, we observed that ELF exhibited a potent anti-HAT activity. As shown in Figure 1A, ELF significantly inhibited HAT activity (left panel), and the IC50 value for HAT inhibition by ELF was 5.33 mg/mL (right panel). To confirm this finding, we next examined HAT activity using nuclear fractions that were isolated from Raw 264.7 cells, as indicated in Figure 1B. Consistent with the results shown in Figure 1A, HAT activity was significantly increased in LPS-treated cells; however, this activity was effectively inhibited by ELF (Fig. 1B). As shown in Figure 1C, ELF did not cause obvious cytotoxicity. These data suggest that ELF has the potential as a novel HAT inhibitor (HATi).

Inhibitory effects of ELF on HAT activity.
ELF is a general HATi
We examined the enzyme specificity of ELF by measuring the p300, CREB binding protein (CBP), and P300/CBP-associated factor (PCAF) in the presence or absence of ELF. As shown in Figure 2, ELF was an effective inhibitor of p300 acetyltransferase activity, with an IC50 value of 6.54 mg/mL (upper panel). Under similar conditions, ELF also inhibited CBP (middle panel) and PCAF activity (lower panel), with IC50 values of 7.54 and 9.36 mg/mL, respectively, indicating that ELF may be an HATi.

Inhibitory effect of TA on global HAT. Enzyme-specific anti-HAT activities of ELF against p300, CBP, and PCAF were measured using purified enzymes. A colorimetric assay was performed to measure enzyme activities in the presence of ELF at the indicated concentrations. The results are presented as percentages relative to the control sample. The IC50 value of ELF inhibition of enzyme activities was calculated using Prism software (right panels). The values presented are the mean ± SD of three independent experiments. Means with different superscript letters are significantly different, P < .05. CBP, CREB binding protein; PCAF, P300/CBP-associated factor.
ELF decreases both mRNA and protein expressions of HATs
We next performed qRT-PCR and Western blot assays to evaluate whether ELF inhibits HAT activity by regulating mRNA and protein expression of HATs. Expectedly, LPS increased the expressions of p300 and CBP compared with those observed in the control group, and these effects were significantly inhibited by ELF treatment (Fig. 3A, B). While PCAF mRNA expression was not increased by LPS stimulation, it was significantly reduced following ELF treatment. PCAF protein expression did not show any change after either treatment (Fig. 3B). Together, these findings suggest that while ELF likely inhibits p300 and CBP activity by modulating their expression at the transcriptional level, it may regulate PCAF activity via a different mechanism.

ELF regulates mRNA and protein expressions of HATs.
ELF attenuates LPS-induced hyperacetylation of histone and nonhistone proteins
The acetylation of nonhistone and histone proteins is well known to stabilize protein levels and the transcriptional activation of various inflammation-related genes, respectively. Therefore, we next examined whether ELF-mediated HAT inhibition affected acetylation patterns at K (Lys) residues in nonhistone and histone proteins in Raw 264.7 cells. The results showed that while LPS treatment increased the acetylation of Lys residues in nonhistone proteins, this LPS-induced hyperacetylation was significantly attenuated by ELF treatment (Fig. 4A). Similarly, LPS treatment also increased histone acetylation; however, ELF treatment selectively reduced the histone hyperacetylation of H4K8, H4K12, and H4K15, but did not affect H3K9 histone acetylation (Fig. 4B). These data suggest that ELF-mediated HAT inhibition affects the acetylation of both nonhistone and histone proteins that mediate inflammatory responses. 27,28

ELF ameliorates LPS-induced hyperacetylation of nonhistone and histone proteins.
ELF blocks p65 translocation by abrogating its acetylation at K310
As discussed, p65 critically mediates inflammatory responses. Specifically, its acetylation at K310 by p300/CBP induces it to be transported from the cytosol to the nucleus, where it then induces transcription of inflammation-related genes. 20 Therefore, we next tested whether ELF abrogates LPS-induced p65 translocation by reducing K310 acetylation. As shown in Figure 5A, p65 was found to be stabilized in the cytosol and to be translocated to the nucleus following LPS stimulation. Interestingly, ELF treatment inhibited this LPS-induced translocation and also attenuated K310 acetylation. To confirm this observation, we performed an immunofluorescence assay, which similarly showed that LPS treatment dramatically induced p65 stabilization and transportation, and that these effects were completely abrogated by ELF treatment (Fig. 5B). Together, these findings indicate that ELF negatively regulates p65 by reducing K310 acetylation and thereby obstructing p65 nuclear translocation.

ELF prevents p65 translocation following stimulation of inflammatory signaling.
ELF decreases the mRNA level of inflammatory cytokines
Finally, to determine whether preventing p65 nuclear translocation downregulates the inflammatory cytokines, we examined the mRNA expression of IL-6, IL-1β, and TNF-α via qRT-PCR. As expected, these genes were markedly increased after LPS stimulation, but markedly decreased in response to ELF treatment. Taken together, these findings show that ELF attenuates IL-6, IL-1β, and TNF-α mRNA levels by negatively regulating p65, and thereby likely ameliorates inflammatory responses (Fig. 6).
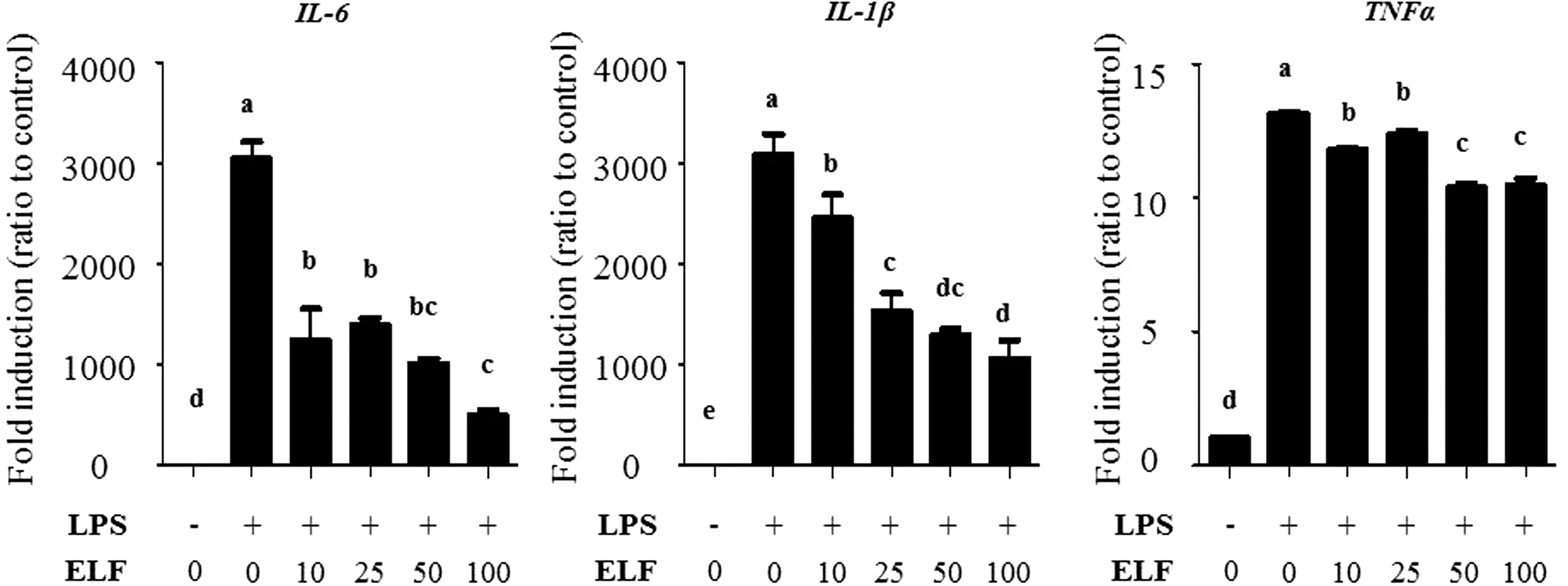
ELF decreases mRNA expressions of inflammatory cytokine genes.
Discussion
In general, proinflammatory action leads to various human diseases. 29 While this relationship between inflammation and disease is well established, the mechanisms that underlie it remain obscure. Thus, we demonstrated a novel epigenetic mechanism that underlies the regulation of inflammatory responses. Recent research has demonstrated the importance of personalized nutrition for maintaining general health; thus, insights into nutritionally modulated epigenetic mechanisms that control inflammation are likely to have important clinical implications.
The present study revealed that ELF is a novel HATi (Figs. 1 and 2). HATis have been shown to mediate the pathogenesis of several human diseases that involve inflammation, including cancer, Alzheimer's disease, diabetes, and hyperlipidemia, through inhibiting catalytic HAT activity. 30 Furthermore, many studies have demonstrated that food and/or food-derived phytochemicals ameliorate inflammatory responses by inhibiting HAT activity. 31 –33
As expected, our data showed a dramatic increase in histone acetylation following LPS treatment, including that of histones H3K9, H4K8, K4K12, and H4K15 (Fig. 4B). HAT-induced histone acetylation is well established to drive transcriptional activation. Specifically, increased acetylation neutralizes the positive charge of histones H3 and H4, and thus renders the chromatin less compact, thereby allowing transcription factors to more easily access the promoter regions of their target (in this case, proinflammatory) genes. 11 Thus, our results are consistent with previously published studies that have reported inflammation-induced patterns of histone hyperacetylation. 34 –36 Interestingly, ELF may specifically attenuate histone H4 acetylation; however, we did not identify the mechanism by which it does so. Similar epigenetic effects have been shown to affect the expression of transcription factors and to consequently regulate inflammatory target genes. 37 Together, these findings support the importance of considering epigenetic mechanisms when investigating inflammation regulation.
It is well known that the natural components of food and/or their derivatives exert epigenetic effects to mediate inflammation. For example, epigallocatechin gallate has been shown to inhibit Epstein–Barr virus-induced B lymphocyte transformation 20 and endothelial cell inflammation 38 by reversing chromatin-HAT binding and inducing histone H3 hypoacetylation. Likewise, gallic acid suppresses LPS-induced inflammatory responses by attenuating HAT activity. 32 Interestingly, both of these substances affect inflammatory phenotypes by epigenetically regulating NF-κB. Consistent with these previous findings, our data show that ELF ameliorates inflammatory responses by controlling NF-κB by epigenetic regulation. Specifically, ELF was herein shown to inhibit NF-κB translocation by blocking acetylation of the p65 K310 residue, which, as discussed, is required to initiate the nuclear translocation of p65 in response to inflammation. 20 Notably, it is also essential to facilitate full p65 transcriptional activity, 39 because acetylated K310 binds to the bromodomains of the Brd4 coactivator to stimulate NF-κB activity, and thereby, NF-κB-dependent inflammatory responses. 40 Indeed, multiple coactivators have been reported to be necessary for p65 acetylation and NF-κB transcriptional activation 41 ; thus, it is possible that ELF may impede the transportation or binding of HAT coactivators to the p65 K310 residue, thereby blocking NF-κB translocation and inhibiting the proinflammatory genes such as IL-1β, IL-6, and TNFα.
LF leaves contain CQA derivatives (comprising 3,4-di-O-CQA, 3,5-di-O-CQA, 4,5-di-O-CQA, 5-O-CQA, 3-O-CQA, and 3-p-O-coumaroyl) as major phenolic constituents, 24,42 and while their anti-inflammatory effects have been well documented, 43,44 few reports have investigated whether and how they modulate epigenetic mechanisms under inflammatory conditions. Nevertheless, the present study is the first to demonstrate that ELF induces anti-inflammatory effects by attenuating HAT activity.
Although our finding supports that ELF has a potential agent that prevents inflammation, this study has a couple of limitations; first, we did not confirm the mechanisms by which ELF disrupts binding between HATs and acetylated K310, and second, how LF CQA derivatives impact the epigenetic regulation of inflammatory responses was not identified. Therefore, in-depth studies should be further conducted.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Main Research Program (E-0150301-05) of the Korea Food Research Institute (KFRI), funded by the Ministry of Science and ICT.