
Abstract
Excessive expression of TNF-α worsens bacterial vaginosis (BV) and osteoporosis. Therefore, to understand whether probiotics could alleviate vaginosis and osteoporosis, we isolated anti-inflammatory Lactobacillus plantarum NK3 and Bifidobacterium longum NK49 from kimchi and human fecal lactic acid bacteria collection and examined their effects on Gardnerella vaginalis (GV)-induced vaginosis and ovariectomy-induced osteoporosis in female mice. Oral gavage of NK3 and/or NK49 significantly alleviated GV-induced vaginosis; these inhibited NF-κB activation and TNF-α expression in the vagina and uterus, and decreased the GV population in the vagina. Furthermore, treatment with NK3 and/or NK49 alleviated ovariectomy-induced osteoporosis and obesity; these increased blood calcium, phosphorus, and osteocalcin levels and suppressed body weight. GV-induced vaginosis and ovariectomy increased colonic myeloperoxidase activity, TNF-α expression, and fecal Proteobacteria population. NK3 and/or NK49 treatments reduced TNF-α expression and NF-κB activation in the colon. NK3 and NK49 treatment also restored GV- or ovariectomy-disrupted gut microbiota composition. In conclusion, NK3 and NK49 may simultaneously alleviate BV and osteoporosis by suppressing NF-κB-linked TNF-α expression through the regulation of gut microbiota population.
Introduction
The vagina, a unique environment for microbial colonization, dramatically changes over the course of a lifetime. 1,2 The vaginal microbiota of post-menarche females predominantly consist of Lactobacilli and anaerobes and recessively facultative and obligate anaerobes. The disruption of hormonal environment, as in menstruation and menopause, can suppress the growth of Lactobacilli and accelerate infections by pathogens such as Gardnerella vaginalis (GV) and Candida albicans. 3,4 GV frequently infects the vagina and simulates NF-κB activation and TNF-α expression, leading to bacterial vaginosis (BV). 5 –7
Post-menopause females frequently experience BV, Candidal vaginitis, osteoporosis, and obesity. 8,9 Menopause increases adipogenesis and osteoclast formation through the upregulation of proinflammatory cytokine expression with estrogen deficiency, resulting in obesity and osteoporosis. 8,9 To cure BV and osteoporosis, antibacterials such as clindamycin and metronidazole and female hormones are frequently used. 10 –12 However, these have severe side effects, such as drug resistance, superinfection, and breast swelling and tenderness. Therefore, many functional foods such as probiotics are frequently recommended for the treatment of BV, osteoporosis, and obesity. 13,14
Probiotics are safe functional foods that antagonize pathogens, modulate host immune system, and exhibit anti-inflammatory effects. 15 –17 Vaginal administration of Lactobacillus fermentum L23 attenuated BV by suppressing GV proliferation. 18 Oral administration of probiotics such as Lactobacillus acidophilus La-14 and Lactobacillus rhamnosus HN001 mitigated vaginosis in mice with GV-induced BV. 19 Lactobacillus johnsonii HY7042 also alleviated BV. 20 Strus et al. reported an interesting finding that, when a mixture of L. fermentum 57A, Lactobacillus gasseri 57C, and Lactobacillus plantarum was orally administered in healthy women, these probiotics were detected in the vagina. 21 Lactobacillus reuteri alleviated osteoporosis in ovariectomized rats. 22 L. reuteri increased bone density in healthy individuals and alleviated primary and secondary osteoporosis. 23,24 Probiotics attenuated obesity in high-fat-diet (HFD)-fed rodents. 25 Nevertheless, the beneficial effects of probiotics against female disorders such as BV, osteoporosis, and obesity have not been thoroughly reported.
Therefore, we obtained the anti-inflammatory L. plantarum NK3 from kimchi lactic acid bacteria collection, and Bifidobacterium longum NK49 from human fecal lactic acid bacteria collection and examined their anti-BV effects in GV-infected mice, as well as their anti-osteoporosis and anti-obesity effects in ovariectomized mice.
Materials and Methods
Materials
RPMI 1640, sodium dodecyl sulfate (SDS), tetramethyl benzidine, β-estradiol-3-benzoate (β-EB), peptidoglycan (PG), lipopolysaccharide (LPS), hydrogen peroxide, and sodium thioglycolate were purchased from Sigma (St. Louis, MO). Antibodies from inducible NO synthase (iNOS), p65, p-p65, cyclooxygenase (COX)-2, and β-actin were purchased from Cell Signaling Technology (Beverly, MA). Acrylamide (30%) for polyacrylamide gel electrophoresis (PAGE) was purchased from Biosesang (Seoul, Korea). GV KCTC5096 was purchased from the Korean Collection for Type Cultures (Daejeon, Korea).
Bacterial culture
GV was anaerobically cultured in a general anaerobic medium (GAM; Nissui Pharm Co, Tokyo, Japan) broth at 37°C for 48 h.
NK3 and NK49 were cultured in general media for probiotics such as deMan, Rogosa, and Sharpe (MRS) broth (BD, Radnor, PA). Cells were harvested and suspended in sterilized phosphate-buffered saline or maltose solution.
Animals
Female C57BL/6 mice (6 weeks old for GV-induced BV and 11 weeks old for ovariectomy-induced osteoporosis) were obtained from Orient Experimental Animal Breeding (Seoul, Korea). Mice were kept in polycarbonate cages at 20–22°C and 50% ± 10% humidity on a 12-h/12-h light/dark period, provided with regular rodent chow and water ad libitum. Mice were acclimatized for 1 week and used in experiments. The animal experiments were approved by the Committee for the Care and Use of Laboratory Animals in the Kyung Hee University [IACUC No. KHUASP(SE)-17-101 and KHUASP(SE)-18-117] and carried out according to the NIH and University Guidelines for the Care and Use of Laboratory Animals.
Preparation of macrophages
Two milliliters of 4% thioglycolate was injected in the intraperitoneal cavity of mice. Peritoneal cavity fluid was collected on the fourth day after intraperitoneal injection and centrifuged at 3000 g for 10 min. Collected cells were washed twice with RPMI 1640, suspended in RPMI 1640 containing 10% fetal bovine serum and 1% antibiotic-antimycotic, incubated at 37°C for 20 h, and washed with RFA thrice according to the method of Jang et al. 25 Attached cells were used as macrophages. Macrophages (1 × 106 cells per well) were treated with probiotics in the presence of LPS or absence of PG for 90 min (for NF-κB activation assay) or 20 h (for cytokine assay).
Dosage regimen of NK3 and NK49
To assay the anti-inflammatory activities of probiotics, isolated macrophages (1 × 106 cells/mL) were treated with probiotics including NK3 and NK49 (1 × 103 and 1 × 105 CFU/mL, suspended in phosphate-buffered saline) as previously reported. 19
To decide the dosage of NK3 or NK49 in mouse experiments, NK3 or NK49 (1 × 107, 1 × 108, and 1 × 109 CFU per mouse per day) was orally administered for 2 weeks in GV-infected mice as previously reported. 19 Of these, treatment with NK3 or NK49 at a dosage of 1 × 109 CFU per mouse per day most effectively suppressed myeloperoxidase activity in the vagina of mice with BV. Therefore, NK3 and NK49 were orally gavaged at a dosage of 1 × 109 CFU per mouse per day in mouse experiments.
Preparation of mice with BV
Mice were separated into eight groups (CON, ES, GV, GLP, GBL, GMX1:1, GMX4:1, and GPC) of seven mice each. Mice, except the CON group, were subcutaneously injected with β-EB (0.5 mg/0.1 mL) daily for 2 days before GV infection according to the method of Jang et al. 19 GV (1 × 106 CFU, suspended in 20 μL saline) was treated intravaginally. Test agents (GV, vehicle [1% maltose] alone; GLP, 1 × 109 CFU per mouse per day of NK3; GBL, 1 × 109 CFU per mouse per day of NK49; GMX, 1 × 109 CFU per mouse per day of NK3 and NK49 [1:1] mixture; GMX4:1, 1 × 109 CFU per mouse per day of NK3 and NK49 [4:1] mixture; GPC, 0.02 mL of 10% clotrimazole) were orally (for NK3 and NK49) or intravaginally (clotrimazole) administered for 2 weeks (6 days per week). The CON group was intravaginally injected with saline instead of GV suspension and β-EB, or 1% maltose instead of test agents. ES group treated with β-EB alone was treated with vehicle instead of GV suspension and test agents. Mice were anesthetized by CO2 asphyxiation. Vagina, uterus, and colon tissues were collected and stored at −80°C for assaying biochemical markers.
Preparation of mice with ovariectomy-induced osteoporosis
Mice were separated into eight groups of six mice each, anesthetized with ether, and bilateral ovaries were resected according to the method of Li et al. 26 Mice were not treated with antibacterials after the operation. Although the sham (SHA) group had surgery, their bilateral ovaries were not resected. Mice were treated with test agents on the eighth day after the operation. Test agents (OV, vehicle [1% maltose]; OLP, 1 × 109 CFU per mouse per day of NK3; OBL, 1 × 109 CFU per mouse per day of NK49; OMX1:1, 1 × 109 CFU per mouse per day of NK3 and NK49 [1:1] mixture; OMX, 1 × 109 CFU per mouse per day of NK3 and NK4 [4:1] mixture; OPC, 1 mg/kg of β-estradiol) were orally gavaged (for NK3 and NK49) or intraperitoneally injected (for β-estradiol) for 4 weeks in ovariectomized mice. Control and SHA groups were orally gavaged with 1% maltose instead of test agents. Mice were sacrificed 18 h after the final treatment with test agents. Uterus, femur, liver, and colon tissues were collected. These tissues were stored at −80°C for assaying biochemical markers.
Myeloperoxidase activity assay
Vagina, uterus, and colon tissues were homogenized in RIPA lysis buffer (300 μL; Biosesang) containing 1% protease and 1% phosphatase inhibitor cocktail and centrifuged (10,000 g, 20 min, and 4°C). 19 Myeloperoxidase activity of the supernatants was assayed in the reaction mixture, which consisted of 0.1 mM hydrogen peroxide, 1.6 mM tetramethyl benzidine, and 50 mM phosphate-buffered saline (pH 7.4), and incubated at 37°C. Absorbance at 650 nm was monitored over time.
ELISA and immunoblotting
For immunoblotting, tissue and cell homogenate supernatants were prepared according to the method of Jang et al. 19 Supernatant proteins were separated by SDS-PAGE, transferred to a nitrocellulose membrane, sequentially incubated with primary antibodies for iNOS, COX-2, p65, p-p65, and β-actin and a secondary antibody, and detected with an enhanced chemiluminescence reagent.
For ELISA, the concentrations of TNF-α, IL-10, and osteocalcin were measured in the supernatants of vagina, uterus, and colon tissue homogenates and bloods using ELISA kits (R&D Systems, Minneapolis, MN). 19
Assay of calcium and phosphorus
Calcium and phosphorus levels in the blood were assayed according to the manufacturer's protocols.
Quantitative polymerase chain reaction
Fecal DNA was purified using a DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany), and quantitative polymerase chain reaction (qPCR) analysis of Firmicutes, Bacteroidetes, δ/γ-Proteobacteria, and GV was carried out using SYBER premix Ex Taq II (TaKaRa, Shiga, Japan) according to the method of Lim et al. 27 Thermal cycling conditions were as follows: initial denaturation at 95°C for 30 sec, denaturation at 95°C for 5 sec, annealing at 63°C for 30 sec, extension at 72°C for 30 sec, and 35 cycles. Bacterial population levels were calculated by comparing with bacterial 16S rRNA. Primers are shown in Supplementary Data.
Reverse transcription qPCR for TNF-α, IL-10, arginase-1 (Arg1), Arg 2, and β-actin was carried out with RNA (2 μg) purified from the liver. 25,28 Thermal cycling conditions were as follows: initial denaturation at 95°C for 30 sec, denaturation at 95°C for 15 sec, annealing at 60°C for 30 sec, extension at 72°for 30 sec, and 40 cycles. Gene expression levels were calculated by comparing with β-actin. Primers are shown in Supplementary Data.
Statistical analysis
All experimental data were described as mean ± standard deviation. The significance was analyzed using one-way ANOVA followed by post hoc analysis using Dunnett's comparison tests (P < .05).
Results
Effects of NK3 and NK49 on NF-κB activation in LPS-stimulated macrophages
First, the anti-inflammatory effects of lactic acid bacterial strains isolated from kimchi and human feces were examined in LPS- or PG-stimulated macrophages. Of tested bacteria, NK3 and NK49 inhibited NF-κB activation the most (Fig. 1). NK3 and NK49 were identified as L. plantarum and B. longum, respectively, based on the results of Gram staining, 16S rDNA sequencing, and using the API 50 CHL Kit (bioMerieux, Seoul, Korea). These were acid-resistant and showed no hemolysis in 7% (v/v) sheep blood agar plate.

Effects of LP NK3 and BL NK49 on NF-κB activation in LPS- or PG-stimulated macrophages.
NK3, NK49, and their mixtures alleviated GV-induced BV in mice
To evaluate whether the oral administration of probiotics NK3 and NK49 alleviated BV, we orally gavaged NK3, NK49, and their (1:1 and 4:1) mixtures on mice with GV-induced BV and examined their anti-BV effects (Fig. 2 and Supplementary Fig. S1). GV increased myeloperoxidase activity and TNF-α expression in the vagina, resulting in BV. The oral administration of NK3, NK49, or their mixtures (M1:1 and M4:1) significantly suppressed GV-induced TNF-α expression and myeloperoxidase activity in the vagina, while IL-10 expression was increased. Furthermore, GV induced COX-2 and iNOS expression and NF-κB activation in the vagina. The oral administration of NK3 and/or NK49 suppressed GV-induced COX-2 and iNOS expression and NF-κB activation in the vagina. Furthermore, these treatments significantly inhibited GV-induced TNF-α expression and myeloperoxidase activity in the uterus, while IL-10 expression was increased (Supplementary Figs. S1B and S2). Next, we investigated the effects of NK3 and/or NK49 on macrophage polarization in the vagina (Fig. 3). These decreased the expression of Arg2 and TNF-α in the vagina, which are M1 macrophage markers, whereas the expression of Arg1 and IL-10, which are M2 macrophage markers, was increased. Furthermore, treatment with NK3 or/and NK49 significantly reduced the GV population (Supplementary Fig. S3).
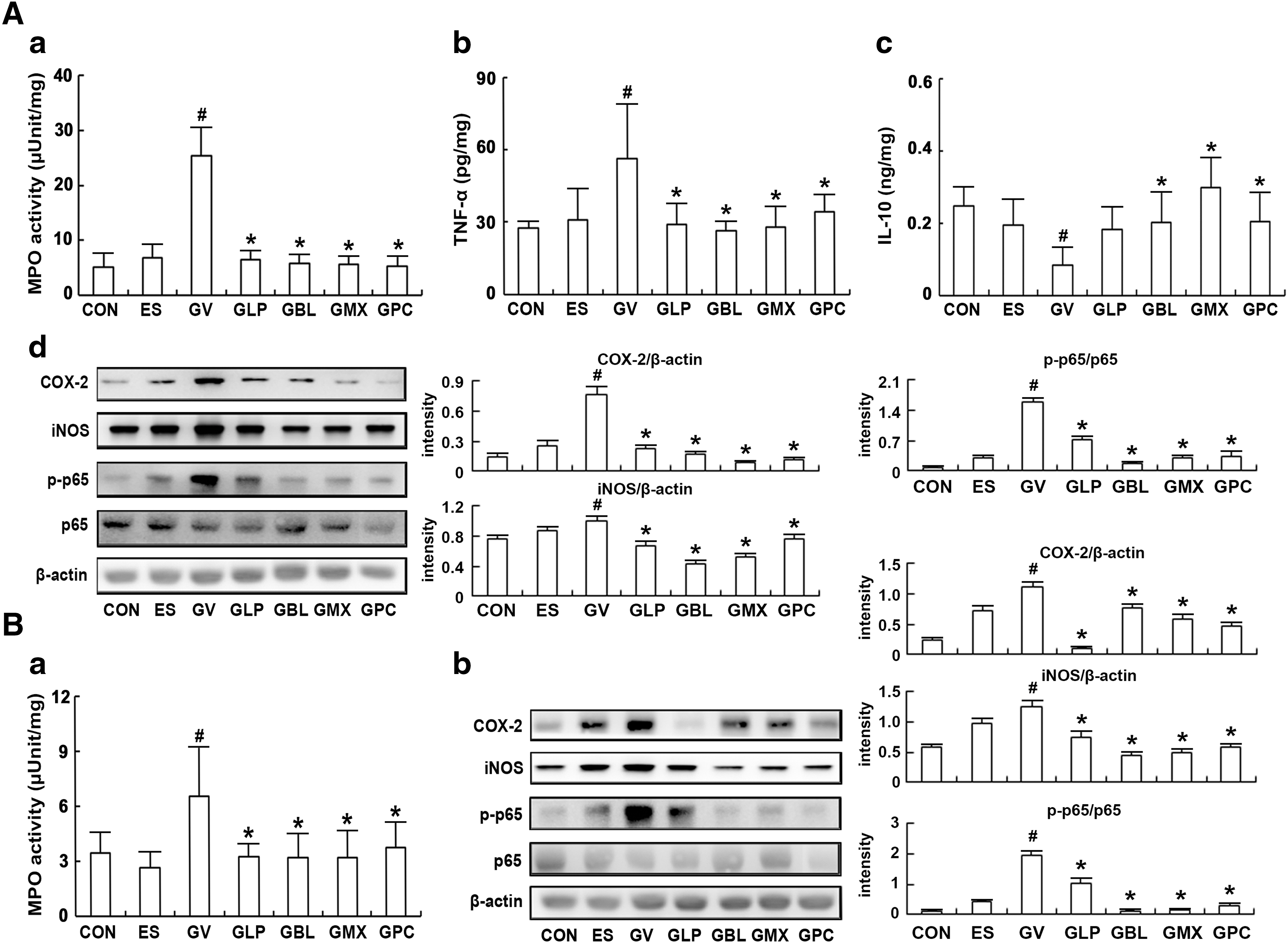
Effects of LP NK3 and BL NK49 on GV-induced vaginosis in immunosuppressed mice.
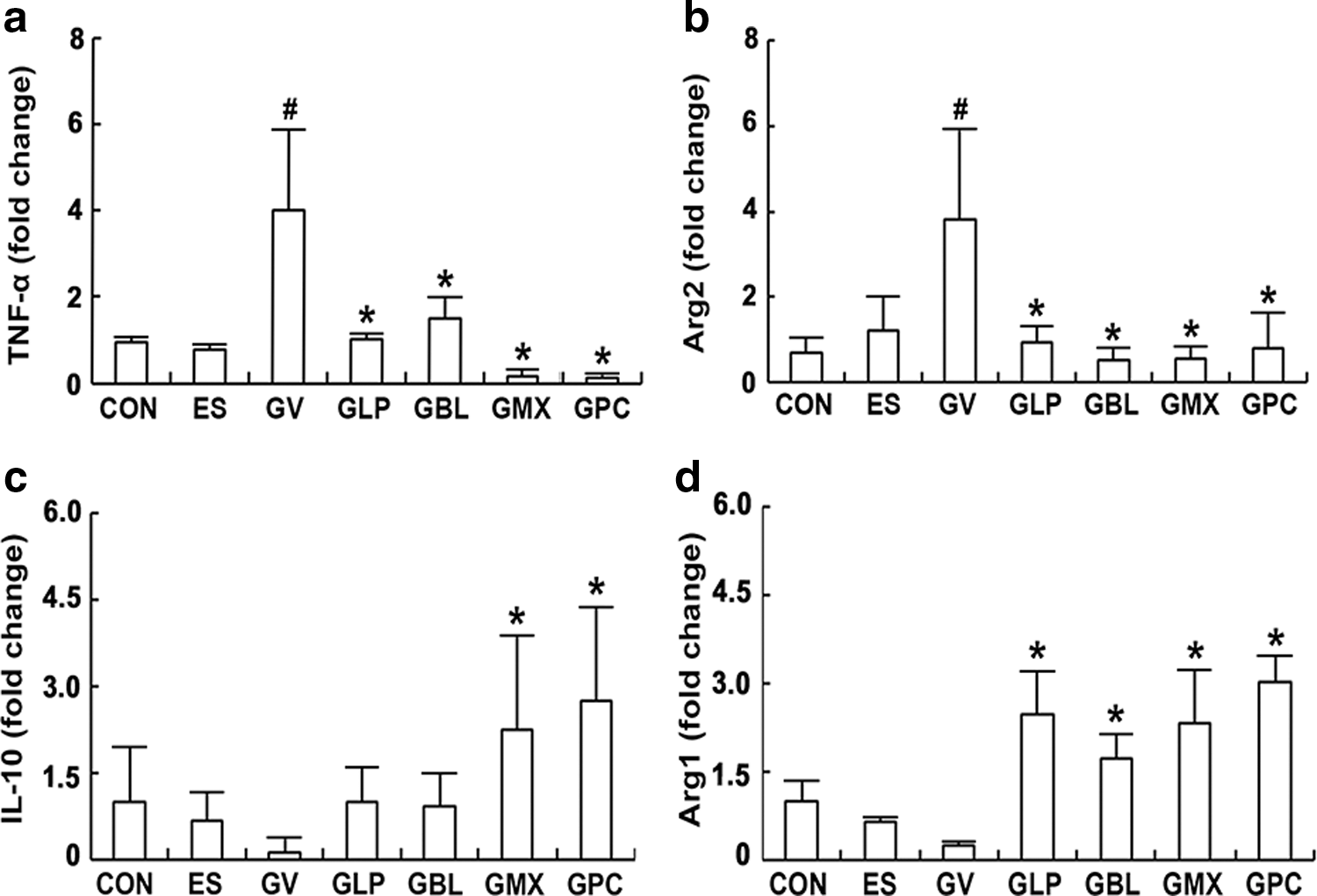
Effects of LP NK3 and BL NK49 on macrophage polarization in the vagina of mice with GV-induced vaginosis. TNF-α
To understand the relationship between vaginosis and gut immunity, we investigated the effects NK3 and/or NK49 on IL-10 and TNF-α expression in the colon of mice with GV-induced BV (Fig. 4A). GV infection significantly increased NF-κB activation, TNF-α expression, and myeloperoxidase activity in the colon, while IL-10 expression was decreased. The oral administration of NK3 and/or NK98 potently mitigated BV-induced colitis; these significantly downregulated NF-κB activation, TNF-α expression, and myeloperoxidase activity, and upregulated IL-10 expression. Next, we investigated fecal microbiota composition in mice with GV-induced BV using qPCR (Fig. 4B). GV significantly increased Proteobacteria population and suppressed Bacteroidetes population in the vagina. However, treatment with NK3 and/or NK49 significantly suppressed Proteobacteria population and increased Bacteroidetes population, thereby decreasing Proteobacteria/Bacteroidetes and Firmicutes/Bacteroidetes ratios. Furthermore, these inhibited gut microbiota LPS production.
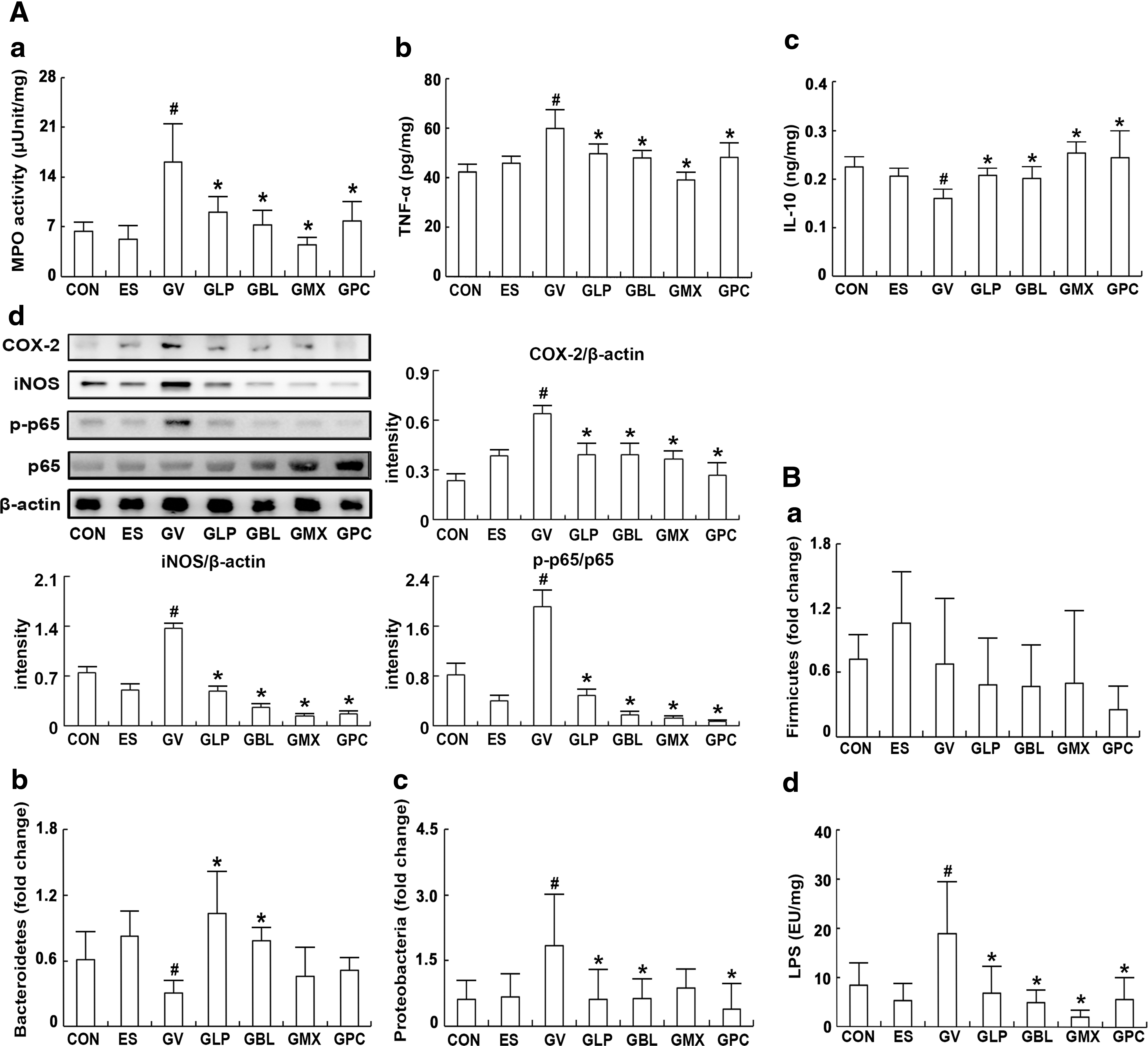
Effects of LP NK3 and BL NK49 on gastrointestinal inflammation and gut microbiota composition in mice with GV-induced vaginosis.
Effects of NK3 and/or NK49 on osteoporosis and obesity in ovariectomized mice
Next, the effects of NK3 and NK49 on osteoporosis and obesity was measured in ovariectomized mice (Fig. 5 and Supplementary Fig. S4). Ovariectomy significantly increased bodyweight and caused a loss of femur and uterus weights and bone density. Ovariectomy also decreased blood calcium, phosphorus, and osteocalcin levels. However, the oral administration of NK3, NK49, or their mixtures strongly restored calcium, phosphorus, and osteocalcin to pre-ovariectomy levels. Moreover, ovariectomy increased NF-κB activation and iNOS, COX-2, and TNF-α expression in the uterus (Fig. 6).
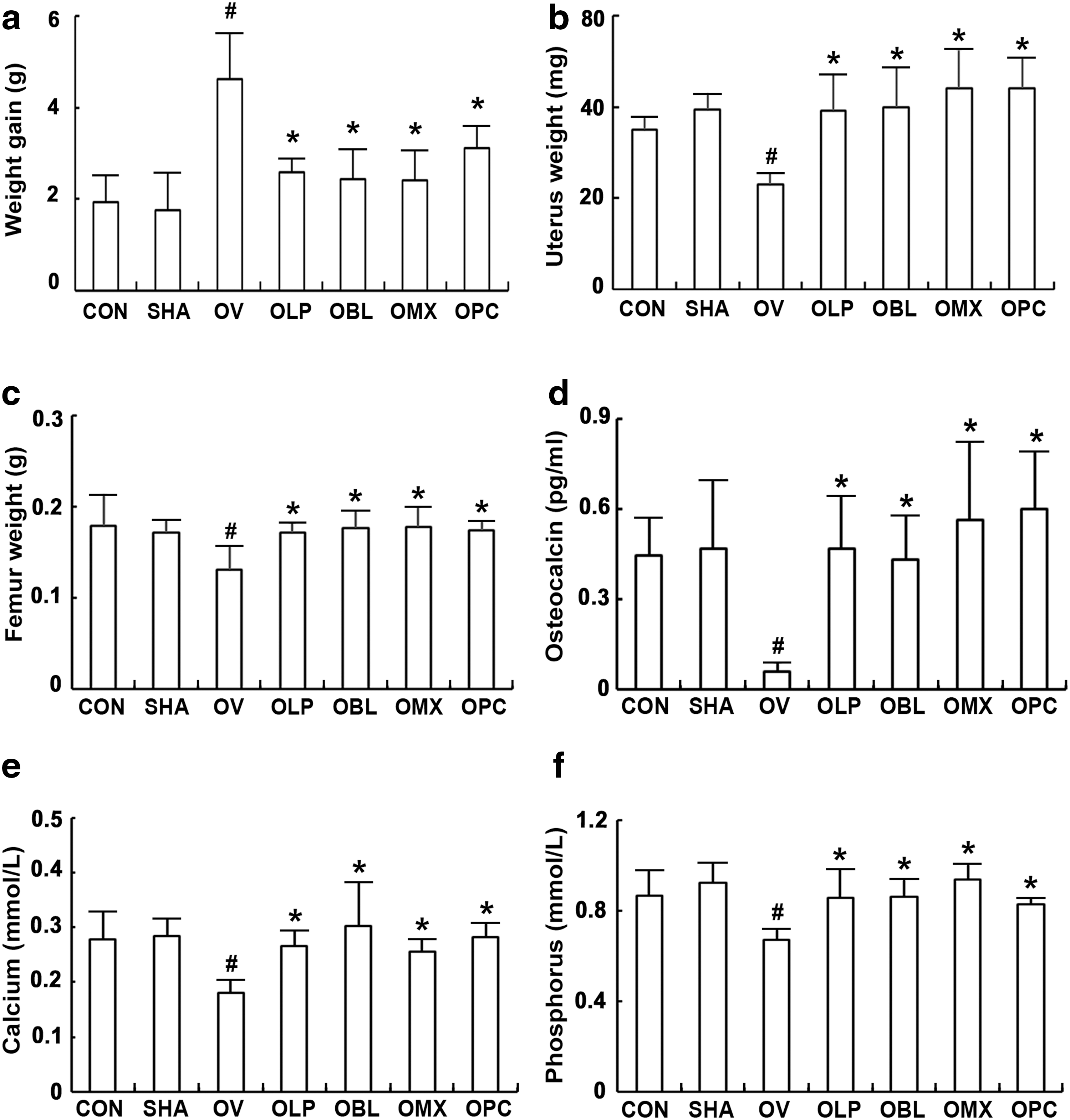
Effects of LP NK3 and BL NK49 on ovariectomy-induced osteoporosis in mice. Effects on body
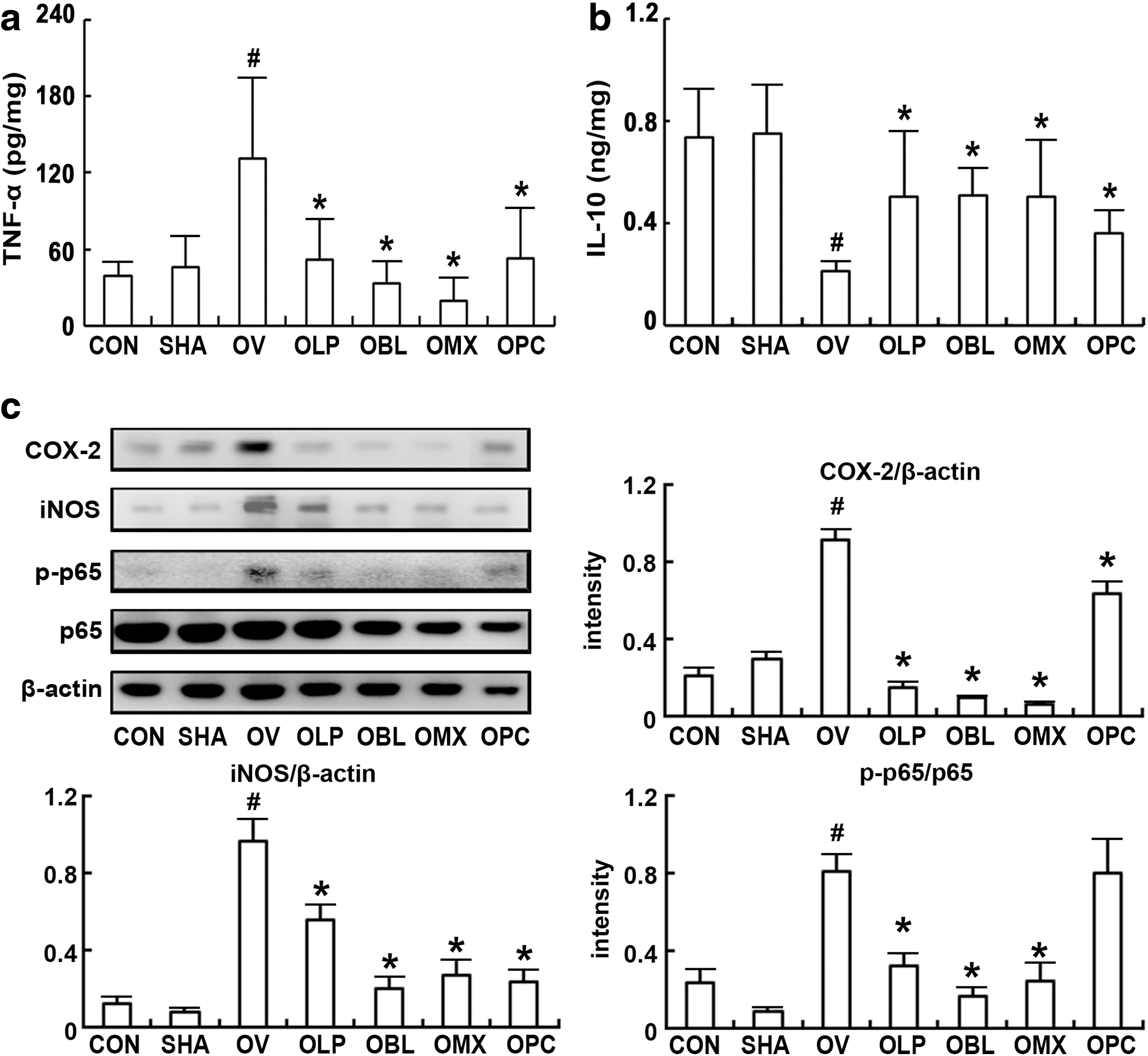
Effects of LP NK3 and BL NK49 on uterus inflammation in mice with ovariectomy-induced osteoporosis. Effects on TNF-α
We also examined the effect of ovariectomy on the colitis in mice (Fig. 7). Ovariectomy significantly increased TNF-α expression and decreased IL-10 expression in the colon. The oral administration of NK3 or/and NK49 suppressed NF-κB activation and TNF-α expression and induced IL-10 expression. Furthermore, ovariectomy disrupted the composition of gut microbiota; it increased the populations of Firmicutes and Proteobacteria, and suppressed the population of Bacteroidetes. Treatment with NK3 or/and NK49 restored ovariectomy-suppressed Bacteroidetes population and ovariectomy-enhanced Firmicutes and Proteobacteria populations. These treatments also suppressed ovariectomy-induced increase in the Proteobacteria to Bacteroidetes population ratio. Furthermore, these treatments inhibited gut microbiota LPS production.
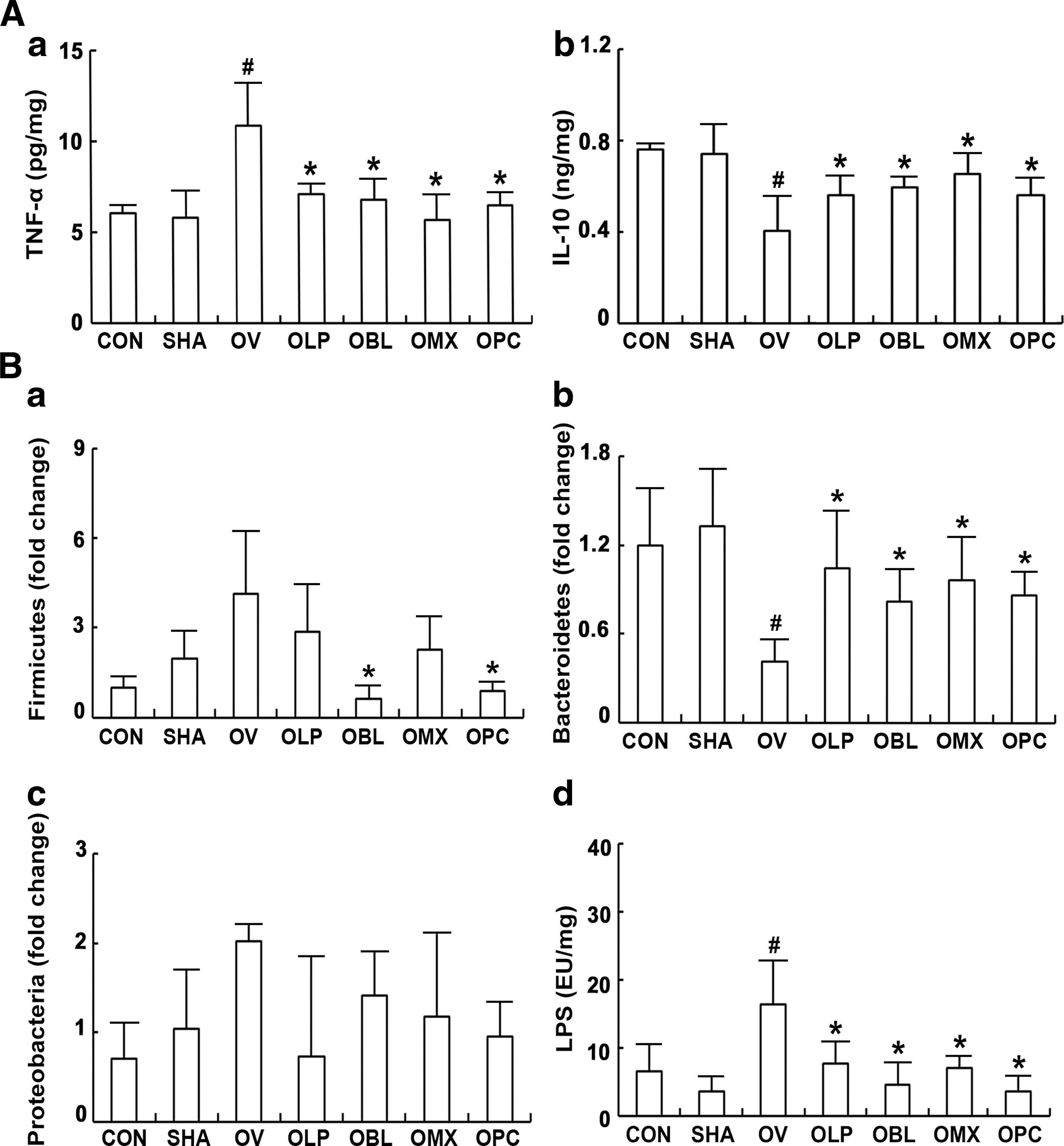
Effects of LP NK3 and BL NK49 on gastrointestinal inflammation and gut microbiota composition in mice with ovariectomy-induced osteoporosis.
Discussion
Osteoporosis, BV, obesity, and depression frequently occur in post-menopausal females. 8,9 Osteoporosis, a prevalent disease in menopausal women, is associated with the proinflammatory cytokine expression levels. Of these cytokines, TNF-α induces osteoclast formation through the stimulation of bone marrow stromal cells, and IL-10 suppresses osteoclastogenesis. 9,29,30 These cytokines are bone resorption stimulants, acting directly on osteoclasts and osteoclast precursors. BV, an inflammatory vaginal disorder, increases NF-κB activation and TNF-α expression in the vagina. 2,20,31
In the present study, GV infection and ovariectomy strongly increased NF-κB activation and TNF-α expression in the vagina, uterus, and colon. These results suggest that GV infection and menopause can stimulate systemic inflammatory responses; in particular, these can cause gastrointestinal inflammation, including colitis. Furthermore, GV infection and ovariectomy induced gut microbiota LPS production and caused gut dysbiosis. Thus, these increased the populations of Proteobacteria and Firmicutes and decreased the population of Bacteroidetes. These findings suggest that GV infection and menopause can cause systemic inflammatory responses as well as gut dysbiosis. Gut microbiota disruption by GV infection and menopause can cause gastrointestinal inflammation, leading to the deterioration of BV and osteoporosis. 32,33
We also found that probiotics NK3 and NK49 inhibited NF-κB activation in LPS- or PG-induced macrophages in vitro. NK3 and/or NK49 also inhibited TNF-α expression and NF-κB activation in GV-induced BV in mice, whereas IL-10 expression was increased. NK3 and/or NK49 alleviated BV-induced inflammation in the colon; these decreased TNF-α expression, NF-κB activation, and myeloperoxidase activity and increased IL-10 expression. Furthermore, these alleviated gut microbiota disruption by GV infection. In addition, the oral administration of L. acidophilus La-14 and L. rhamnosus HN001 attenuated BV in GV-infected female mice. 11 These findings suggest that probiotics can alleviate BV in vivo by regulating gastrointestinal inflammation and microbiota.
The oral administration of NK3 and/or NK49 alleviated ovariectomy-induced osteoporosis in mice; these increased ovariectomy-suppressed blood osteocalcin, phosphorus, and calcium levels and uterus and femur weights, as well as decreased bodyweight. These results suggest that NK3 and NK49 can alleviate osteoporosis in menopause women through osteoblast activation. L. rhamnosus HN001 induced mineral absorption and bone density in ovariectomized female rats. 34 L. reuteri suppressed bone loss in ovariectomized mice. 35 HFD induced obesity by regulating gut microbiota composition. 36 Lactobacillus sakei OK67 suppressed HFD-induced obesity by regulating gut microbiota LPS production. 28 These results suggest that NK3 and NK49 can alleviate osteoporosis and obesity in menopause women by suppressing gut bacterial LPS production. Furthermore, NK3 and NK49 mitigated anxiety/depression-like behaviors and IL-6 expression in mice exposed to immobilization stress (Supplementary Fig. 5). These findings suggest that NK3 and NK49 can simultaneously alleviate menopausal symptoms, including osteoporosis, obesity, BV, and depression.
In conclusion, NK3 and NK49 may interact with the epithelial cells and act as a barrier in the gastrointestinal tract to modulate gut microbiota and immune cells, such as macrophages, dendritic cells, and intestinal lymphocytes, leading to a reduction in TNF-α expression. The suppression of the proinflammatory cytokine expression and gut microbiota LPS production by NK3 and NK49 can mitigate vaginal inflammation and osteoclast formation through the alleviation of gut inflammation, leading to the attenuation of BV, osteoporosis, and obesity.
Footnotes
Acknowledgments
This research was supported by the Medical Research Program through the National Research Foundation (NRF) of Korea, funded by the Ministry of Science and ICT (NRF-2017R1A5A2014768).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.