
Abstract
Polyacetylenes in the bark of Dendropanax morbifera trees have been reported to promote immune cell proliferation and to strengthen the innate immune system. The immunomodulatory potential of D. morbifera branch water extract (DBW) was evaluated by determining its effect on cell viability and the expression of cytokines and immune effector molecules in mouse RAW264.7 macrophages and splenocytes. Production of nitric oxide (NO), inducible nitric oxide synthase (iNOS), and cytokines (interleukin [IL]-1β, IL-2, and IFN-γ) in RAW264.7 macrophages increased after treatment with DBW. The activation of components of the NF-κB signaling pathway, including the phospho-IκBα and the expression and translocation of p65, a subunit of NF-κB, were also increased in RAW264.7 mouse macrophage cells after treatment with DBW. In addition, when mice were orally administered DBW, splenocyte cytokines and NO production were increased in a dose-dependent manner relative to control-treated mice. Furthermore, natural killer cell activity in DBW-treated mice was determined by lactate dehydrogenase (LDH) release assay. LDH release also increased in response to DBW treatment. Taken together, these results indicate that D. morbifera extract enhances innate immunity by promoting NF-κB signaling, leading to increased expression of proinflammatory cytokines and effector molecules. DBW therefore has potential therapeutic use in the context of immune stimulation.
Introduction
In healthy individuals, transformed or cancerous cells continuously arise as a result of errors during cell division and are continuously removed by innate immune cells, including macrophages and natural killer (NK) cells. 1 If this balance is disrupted due to weakened immune responses, cancer can arise. 2 The immune system can become weakened due to many factors, including environmental toxins, irregular eating habits, stress, and lack of sleep, in addition to cardiometabolic syndromes such as hypertension, diabetes, and hyperlipidemia. 3 In healthy individuals, age is an important cause of weakened immune responses and can lead to increased susceptibility to infections and cancer. 4
Recently, there has been a growing interest in the medicinal uses of the Dendropanax morbifera tree, also known as the ginseng tree, which is native to Korea. As the interest and demand for D. morbifera products has increased in China, competition to secure the raw material for these products is gradually intensifying. The medicinal effects of D. morbifera trees are mentioned in historical texts such as the Donguibogam and the Uibangyuchwi, and recent publications have reported the immunostimulatory properties of D. morbifera extracts. 5,6 The polyacetylenes contained in D. morbifera trees promote the proliferation of immune cells and strengthen innate immune responses. D. morbifera extracts also have cytoprotective and skin whitening effects, suggesting the presence of inhibitors of tyrosinase and melanin formation. In addition, recent studies have reported anticancer effects, antimicrobial activity, and benefits in patients with diabetes and hyperglycemia. D. morbifera extracts contain numerous physiologically active substances and have been shown to protect against oxidative stress. 6 –10
To increase the utility of D. morbifera seedlings as an industrial material, extracts were prepared from the leaves and branches of 3–5-year-old or 15-year-old seedlings, and these extracts were evaluated for immunostimulatory effects. In vitro and in vivo experiments were carried out to evaluate the effects of the D. morbifera extracts on the function of mouse macrophages.
Materials and Methods
Cell culture
RAW264.7 cells were maintained in Dulbecco's modified Eagle's medium (DMEM; Hyclone/Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 100 U/mL penicillin, 100 mg/mL streptomycin, and 10% fetal bovine serum. Cells were grown at 37°C with 5% CO2.
Preparation of D. morbifera leaf, branch, sapling, and mixed extracts
D. morbifera leaf, branch, sapling, and mixed (DL, DB, DS, and DM) extracts were prepared by adding materials to 600 L water (DLW, DBW, DSW, and DMW) or 20% or 50% ethanol (DLE20 or DLE50, DBE20 or DBE50, DSE20 or DSE50, and DME20 or DME50) and incubating at 80°C for 3 h. Subsequently, extracts were filtered and concentrated to 60 L before freeze-drying; dried extracts were then dissolved in water for 10 min. The mixed extract consisted of a 2:8 ratio of leaf and branch extracts. Solutions were then centrifuged at 17,000 g for 10 min and supernatants were collected.
Preparation of lipopolysaccharide, red ginseng extract, and dextrin
Lipopolysaccharide (LPS) was prepared from Escherichia coli strain O127:B8 (L4516; Sigma-Aldrich, St. Louis, MO, USA). Red ginseng extract (RGE; Korea Ginseng Corporation, Daejeon, Korea) was purchased as a 30% solution, and then filtered with a 0.2 μM syringe filter, and diluted to a 5% solution in water. Dextrin (Dex; Samyanggenex, Incheon, Korea) was diluted to 100 μg/mL in water. Rhodiola Rosea extract (Honggyeongcheon [HONG]; Jinyong, Suwon, Korea) was diluted to 200 μg/mL in water.
Preparation of mouse splenocytes
Mice were sacrificed, spleens were removed, and splenocyte suspensions were prepared. Splenocytes were washed three times with phosphate-buffered saline (PBS) and suspended in complete DMEM. The cells were then seeded in culture dishes at a density of 1 × 106 cells/mL. After 24 h, the viability was measured and cytokine expression was evaluated by quantitative polymerase chain reaction (qPCR).
Cell viability
The cytotoxic effects of each extracts and LPS were evaluated by WST-1 assay. WST-1 solution was added directly to the culture media (10 μL per well) and cells were incubated for 3.5 h. After 3.5 h, the optical density of the supernatants was measured at 450 nm.
Cytokine expression
Quantitative real-time polymerase chain reaction (qRT-PCR) was used to evaluate cytokine expression levels in LPS- or extract-treated cells or in the splenocytes of mice treated with DBW (500, 1000, or 2000 mg/kg body weight), Dex (100 mg/kg body weight), or RGE (200 mg/kg body weight). Total RNA from splenocytes of drug-treated mice was prepared according to the manufacturer's protocol (iNtRON Biotechnology, Inc., Korea). All samples were normalized to the levels of GAPDH and expressed as the fold change. All reactions were done in triplicate. The relative expressions and standard deviations (SDs) were calculated using the comparative quantification method (Supplementary Fig. S1).
Nitric oxide production
RAW264.7 cells (1 × 106 cells/mL) were plated for 18 h, and then extracts of DL, DB, DS, and DM (25, 50, and 100 μg/mL) and LPS (1 μg/mL) were added and the cells or splenocytes were incubated for an additional 24 h. The concentration of nitrite in the culture supernatants was measured by adding 100 μL Griess reagent (1% sulfanilamide and 0.1% N-(1-naphthyl)-ethylenediamine dihydrochloride in 5% phosphoric acid) to 100 μL culture media.
Western blot analysis
RAW264.7 cells or splenocytes from mice treated with DBW were lysed in lysis buffer (50 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1% NP-40, 10 mM NaF, and 10 mM sodium pyrophosphate) and incubated for 30 min on ice. Lysates were centrifuged at 20,000 g for 10 min at 4°C and total proteins were separated by SDS-PAGE on 8% or 12% polyacrylamide gels and transferred to nitrocellulose membranes. Membranes were blocked by incubating for 2 h or overnight at 4°C in 5% nonfat Difco skim milk (BD Biosciences, San Jose, CA, USA) in PBST. The following antibodies were used: anti-p-Akt (4070s; Cell Signaling Technologies, Danvers, MA, USA), anti-Akt (9272s; Cell Signaling), anti-p-ERK (9101s; Cell Signaling), anti-ERK (9102s; Cell Signaling), anti-p-IκBα (2859s; Cell Signaling), anti-IκBα (9242s; Cell Signaling), anti-p-NFκB (3033s; Cell Signaling), anti-NFκB (6956s; Cell Signaling), and anti-β-actin (A5441; Sigma-Aldrich). After washing three times with PBST, membranes were incubated with HRP-conjugated antibodies for 1 h. Protein signals were then visualized.
Luciferase assays for NF-κB transcriptional activity
To measure NF-κB transcriptional activity, TLR4-expressed 293T cells were transiently transfected into a reporter construct encoding firefly luciferase driven from the NF-κB promoter. Renilla luciferase reporter plasmid was included as an internal control. Then, luciferase activity was measured according to the manufacturer's instructions (Promega, Madison, WI, USA). Firefly luciferase activity was normalized relative to the Renilla luciferase activity and presented as the mean (±SD) of three independent experiments.
Treatment of animals with DBW
Animal protocols were approved by the Institutional Animal Care and Use Review Committee of the University of Ulsan College of Medicine (2017-02-148; Seoul, South Korea). Male 6-week-old BALB/c mice were distributed into five or six groups of five to six mice per group. The highest dose of extract used was 2000 mg/kg body weight; control groups were given 500, 1000, and 2000 mg/kg body weight of the vehicle. Mice were dosed orally once a day for a total of 8 days (short-term) or once every other day for 30 days (long-term).
Plasma biochemistry
Plasma was obtained by centrifuging blood with anticoagulant and the levels of alanine aminotransferase (ALT, K752-100; BioVision, Milpitas, CA, USA), aspartate aminotransferase (AST, K753-100; BioVision), creatine, and urea were spectrophotometrically measured at absorbances of 570nm or 450nm.
Measurement of lactate dehydrogenase activity
Lactate dehydrogenase (LDH) activity was measured in serum using an LDH Enzymatic Assay Kit (XpressBio, USA, 3460-04). Whole blood was collected and incubated at 37°C for 10 min. Samples were centrifuged at 9,600 g for 10 min, and the serum layer was collected. LDH Reagent Mix was combined with 5 μL of sample or standard in microplates, and the absorbance was measured at 340 nm. A second reading was taken at the same absorbance after 5 min.
Statistical analysis
Statistical analyses were performed using SPSS (SPSS, Inc., Chicago, IL, USA). Differences between treatment groups were identified using ANOVA and were considered significant when P < .05.
Results
Effects of D. morbifera extracts on cell viability
D. morbifera (DL, DB, DS, and DM) extracts were prepared from water and ethanol, and the viability of RAW264.7 macrophages treated with each extract was tested by WST-1 assay. First, RAW24.7 cells were treated with DLW, DLE20, DLE50, DBW, DBE20, DBE50, DSW, DSE20, DSE50, DMW, DME20, and DME50 at 25, 50, and 100 μg/mL for 24 and 48 h. Cell viability did not decrease in response to treatment with the DL, DB, DS, or DM extracts at 24 h (Fig. 1A) or 48 h (Fig. 1B). These data confirm that D. morbifera extracts do not induce cytotoxicity within at 48 h in concentrations of up to 100 μg/mL.

Cell viability effects of DL, DB, DS, and DM extracts during 24 and 96 h in RAW264.7 cells. Except for the Dendropanax morbifera leaves extracts, the remaining extracts showed no toxicity at various concentrations.
D. morbifera extracts increased cytokine expression and nitric oxide production
Next, cytokine expression was measured by qPCR in RAW264.7 cells treated with D. morbifera. DL or DB extracts increased the expression of interleukin (IL)-2 in a dose-dependent manner (Fig. 2A). LPS was used as a positive control. These results suggest that D. morbifera extracts can improve innate immune function. 11 Next, COX-2 expression was examined in response to treatment with DL extracts and DB extracts. 12 DB extracts induced higher expression of COX-2 than DL extracts (Fig. 2B). Finally, nitric oxide (NO) activity was measured in RAW264.7 cells. DL, DS, and DM extracts did not induce NO activity, although the DB extracts did (Fig. 2C). These results suggested that DBW extracts had the strongest immunostimulatory properties of the four preparations. 13 –16

DB extracts enhanced cytokine expression and NO activity in RAW264.7 cells. The efficacy of the extracts on cytokine expression and NO activity was confirmed.
DBW induces NF-κB activity
To investigate the mechanism of DBW-mediated immune enhancement, 17 activation of the AKT and NF-κB pathways in response to LPS or DBW was evaluated in RAW264.7 cells by western blot analysis. AKT and IκBα were phosphorylated after exposure of the cells to LPS or DBW (Fig. 3A). We also measured the transcriptional activity of NF-κB following treatment of 293T cells with LPS, HONG extracts, RGE extracts, 18 or DBW by luciferase assay. DBW and RGE increased NF-κB transcriptional activity to a comparable degree (Fig. 3B). These results suggest that DBW induces immunostimulatory effects by regulating NF-κB signaling. 19 –21 These results also suggest that the immune-stimulatory effects of DBW are comparable to those of HONG and RGE.
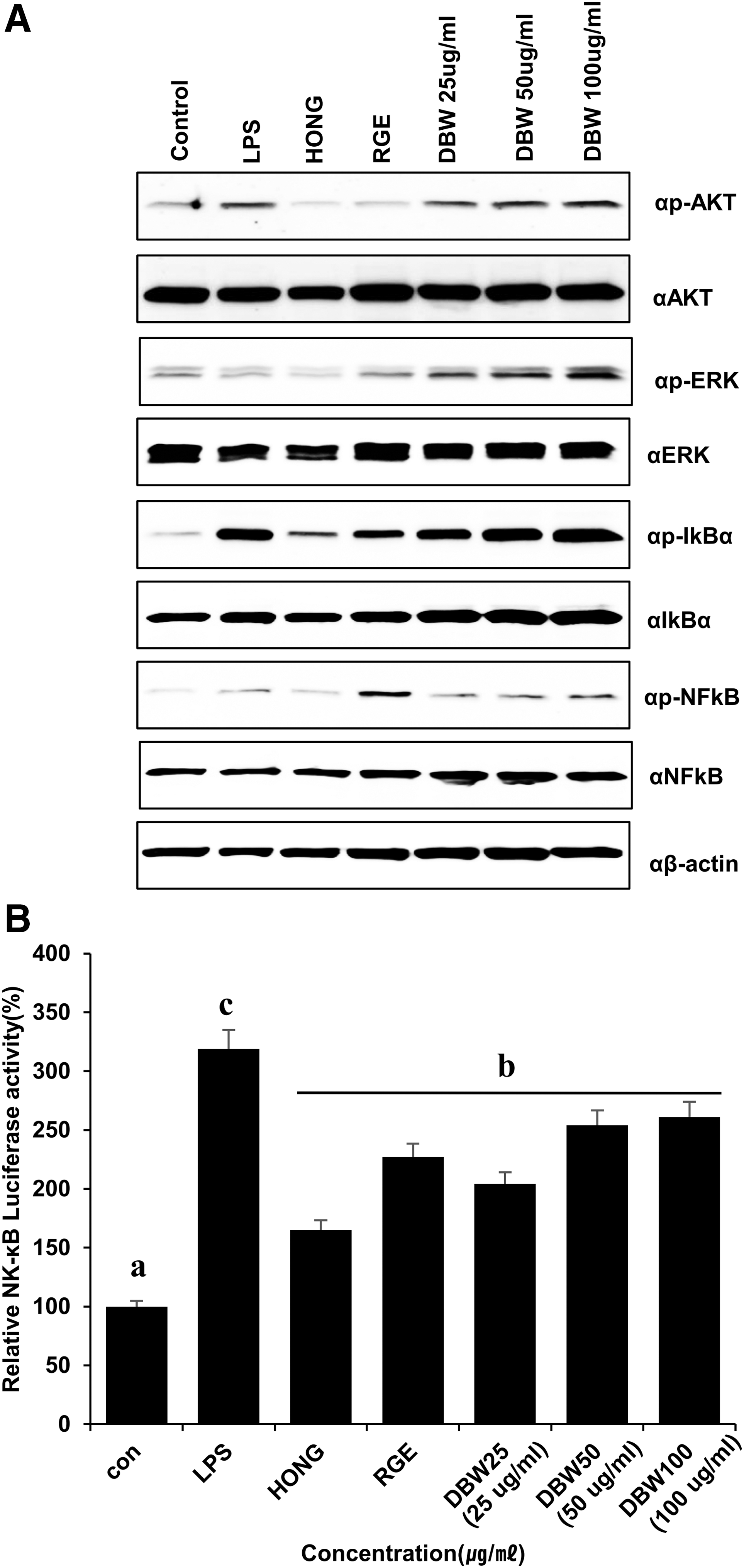
DBW induces the expression of transcription factors. DBW enhances the p-IκBα level and NK-κB promoter activity in RAW264.7 cell lines.
In vivo toxicity of DBW
To investigate the in vivo toxicity of DBW, body weight loss was measured in BALB/c mice treated with dextrin, RGE, or DBW for 8 days (short-term in vivo toxicity tests, Fig. 4A) or 30 days (long-term in vivo toxicity tests, Fig. 4D). As shown in Figure 4A and D, the weights of mice treated with DBW (500, 1000, or 2000 mg/kg) did not change compared to control- or dextrin-treated mice. Furthermore, DBW did not change the absolute or relative weights of the internal organs at any concentration (Supplementary Fig. S2). Furthermore, to confirm that DBW was not toxic to the mouse splenocytes, splenocytes were isolated from mice following short-term or long-term treatment with DBW. The splenocytes were cultured and the viability was evaluated after 48 h (Fig. 4B, E). In these results, DBW did not induce toxicity in mouse splenocytes. 22,23 In addition, the plasma ALT and AST levels were measured in serum of mice after short-term (Fig. 4C) or long-term (Fig. 4F) DBW treatment. These liver function assays showed fluctuations in AST and ALT levels within the normal range. The mean AST and ALT levels were not significantly different between untreated and DBW-treated mice. The mean creatine and urea levels in untreated and DBW-treated mice were also not different. Thus, DBW did not alter the liver function. Finally, to assess liver toxicity, liver tissue was extracted and hepatic toxicity was assessed by hematoxylin and eosin staining of liver tissues (Fig. 4G). DBW did not result in liver toxicity after either short-term or long-term treatment.

In vivo toxicity of DBW. DBW was not affecting the short-term (8 days) and long-term (30 days) toxicity in vivo mouse.
DBW increases NK cell activity by increasing the expressions of cytokines and production of NO and LDH
Primary splenocytes from mice treated with DBW (1000 and 2000 mg/kg body weight) showed increased expression of inducible nitric oxide synthase (iNOS) and IL-6 mRNA after short-term (8 days) treatment (Fig. 5A). 2,12,24 In splenocytes from mice after long-term (30 days) treatment with DBW, iNOS, IL-6, IL-1β, and IL-10 levels were increased (Fig. 5B). 25 –27 The increased expression of IL-6 was also confirmed at the protein level by western blot (Fig. 5C). Next, the effects of DBW on NO and LDH production were evaluated after long-term treatment with DBW. In mice treated with DBW, NO levels in the blood increased in a dose-dependent manner compared to control-treated mice (Fig. 5D). This result was consistent with the production of NO from DBW-treated macrophages in vitro. These results suggest that DBW is an effective mediator of immune signaling. Furthermore, the modulation of NK cell activity by DBW was evaluated by LDH release assay. As shown in Figure 5E, LDH levels increased in mice treated with DBW. Taken together, these results demonstrate the immune-stimulatory effects of DB extracts in vivo.
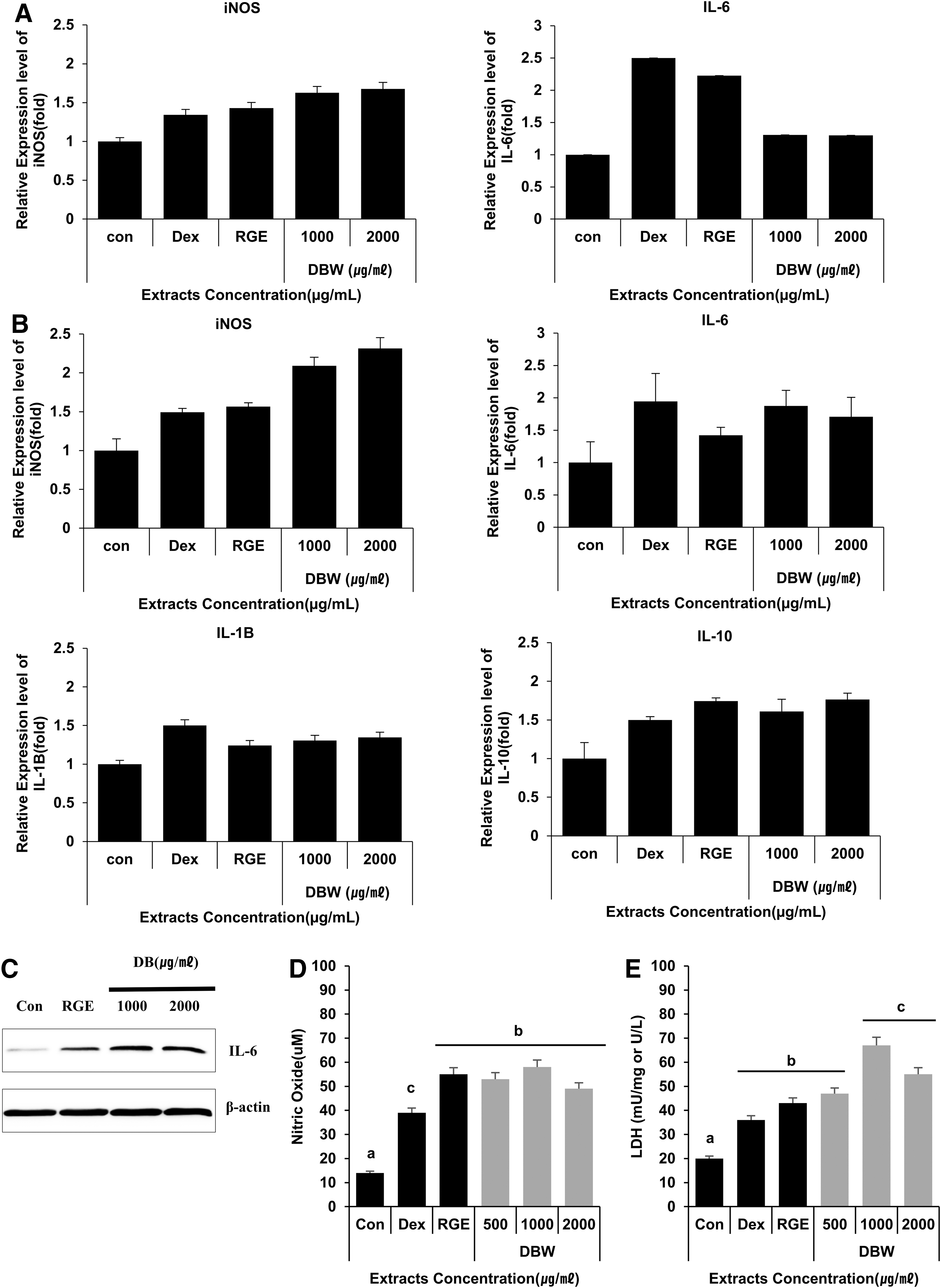
In vivo induction of cytokines by DBW in mouse primary splenocytes. Oral administration of DBW enhanced secretion of cytokines in mouse primary splenocytes.
Discussion
Recently, there has been a growing interest in the medicinal use of D. morbifera extract. D. morbifera extract contains numerous physiologically active compounds and has been reported to have anticancer and antimicrobial properties, in addition to beneficial effects in patients with metabolic syndromes and related diseases such as diabetes. In this study, we examined the ability of DBW extract to enhance innate immune activity in cultured macrophages and in mice. 6,11,19,28
The main organ of immunity in the human body is the spleen, and increases in cytokine production in splenocytes are a key indicator of immune reactivity. 22,29,30 For this reason, we tested the immunomodulatory effects of D. morbifera extracts in vitro on RAW264.7 cells and in vivo by oral administration to mice. DBW enhanced both NO production and cytokine (IL-1β, IL-2, and IFN-γ) expression. This is the first study to demonstrate the nontoxic, immune-stimulating effects of D. morbifera extract in vitro and in vivo. The differences in the biochemical profile of mice treated orally with DBW daily for 8 days were examined. In addition, we investigated the toxicity of DBW as a part of a safety test. DBW was administered orally to mice at two dose levels of DBW (1000 and 2000 mg/kg body weight). Oral DBW did not elicit any notable toxicity, such as changes in body or organ weights, at either dose. Taken together, the current study demonstrates that DBW has immune-stimulating effects, enhancing NO, IL-1β, IL-2, IFN-γ, and iNOS expression, and is nontoxic even at high doses.
In this study, DBW did not induce cytotoxicity against RAW264.7 macrophages at concentrations of 25–100 μg/mL. In fact, DBW increased the cell viability at 24 h within this concentration range. It also increased the expression of immunostimulatory cytokines (IFN-γ, IL-6, and IL-10). 25 –27 Transcription factors such as NF-κB alter cytokine expression. NF-κB activation, in turn, is regulated by the IκB complex. phospho-IκB was increased in response to DBW treatment, suggesting that DBW induces the secretion of immunostimulatory cytokines by activating the NF-κB pathway. The production of NO and LDH were also increased in DBW-treated mice. All living cells contain LDH, but it does not penetrate the cell membrane. When transformed or infected cells are attacked by NK cells, LDH is released. Therefore, LDH release is an indicator of NK cell activity. 31
In conclusion, we found that DBW enhanced cytokine expression in cultured macrophages and in mice splenocytes. We further examined whether immune activation in response to DBW occurs via the NF-κB pathway. 19,21 DBW induces the production of proinflammatory cytokines such as IFN-γ and IL-6 by increasing the phosphorylation of IκBα and thereby inducing NF-κB transcriptional activity. 32 DBW therefore has potential in the future development of treatments to promote immune activation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, and Forestry (IPET) through the Agri-Bio Industry Technology Development Program, funded by the Ministry of Agriculture, Food, and Rural Affairs (MAFRA) (116093-3).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.