
Abstract
Much research has indicated that alcoholic liver disease (ALD) is associated with oxidative stress and inflammation induced by ethanol, and that numerous antioxidants could effectively alleviate such injuries. Moreover, recent studies have identified andrographolide (AD) as having strong anti-inflammatory and antioxidant activities, which can block oxidative damage associated with nuclear factor kappa B (NF-κB)-mediated inflammation. However, the biological role and potential mechanism of AD in its protection against ALD have not been fully characterized. To observe the possible effect of AD, male C57BL/6J mice received ethanol through intragastrical gavage for 12 weeks in this study. The ethanol group was separated into five subgroups: (1) model group (n = 10); (2) silymarin group (0.1 mg/g body weight [BW], n = 10); (3) AD (0.05 mg/g BW) group (n = 10); (4) AD (0.1 mg/g BW) group (n = 10); and (5) AD (0.2 mg/g BW) group (n = 10). Mice in AD groups were treated orally by gavage once per day. The experimental results show that serum aminotransferase, liver lipids, lipid peroxidation, and antioxidant capacities were significantly changed in the model group after alcohol treatment, and the liver tissue histological findings showed pathological changes. Compared with the model group, treatment with AD improved serum aminotransferase, liver function, lipid accumulation, and hepatic reactive oxygen species levels. And AD decreased the hepatic NF-κB and tumor necrosis factor alpha (TNF-α) protein expression of ALD mice. This research demonstrated that AD can alleviate liver pathological injury and oxidative stress in mice exposed to ethanol by decreasing the expression of NF-κB and TNF-α.
Introduction
Alcoholic liver disease (ALD), one of the most serious consequences of excessive alcohol consumption, is one of the common diseases globally. 1 According to a recent WHO report, 3 million global deaths could be attributed to alcohol consumption in 2016, 2 and ALD has become a serious public health problem in many countries. 3,4
Although the pathogenic factor of ALD is single, its pathological process is very complicated. Cytokines, immune mediation, apoptosis, virus superposition, endotoxin, and genetic polymorphisms are all involved in the pathological process of ALD. 5 –10 The occurrence of ALD is usually accompanied by the generation of abundant inflammatory cytokines. As important inflammatory mediators, the function of tumor necrosis factor alpha (TNF-α), nuclear factor kappa B (NF-κB), and other proinflammatory cytokines has been comparatively and comprehensively studied in the etiopathogenesis of ALD. The transcription of many inflammatory-related genes, such as interleukin (IL)-1β, IL-6, and TNF-α, can be promoted by the activation of NF-κB. 11 Nevertheless, there is no specific clinical treatment for ALD so far. 12
In recent years, the clinical application of botanical drugs has gradually been recognized. Andrographolide (AD) is the main active ingredient of Andrographis paniculate, which is a kind of botanical drug. 13 Owing to its remarkable curative effect and few adverse reactions, AD-related preparations have been widely used as clinical drugs for bacterial infection and upper respiratory tract infection in Asia. 14,15 AD has strong anti-inflammatory and antioxidant abilities, which mainly act by blocking oxidative damage and NF-κB activation, thereby suppressing the release of TNF-α. 16,17 Therefore, it may be hypothesized that AD possesses potent anti-inflammatory properties that may be beneficial for the treatment of ALD. However, the effect of AD on ethanol-induced hepatic damage has not been adequately studied.
This study aimed to investigate the potential of AD as a novel therapy for ALD by evaluating its hepatoprotective effects on a mouse model of ethanol-induced liver injury, thereby providing preventive and therapeutic strategies for ALD, and developing the potential value of AD in clinical application.
Materials and Methods
Chemicals
The chemical reagents used in this article were as follows: silymarin (Novartis International AG, Basel, Switzerland) and AD (purity ≥99%; Beijing Solarbio Technology Co., Ltd, China).
Animals and hepatic injury treatments
Our studies were conducted in accordance with the protocols and with the approval by the Animal Ethics Committee of Jilin University approved by the Ministry of Health, China. The mouse license for the laboratory (no. SCXK 2015-0001) was issued by the National Science and Technology Department of Jilin Province. Eight-week-old male C57BL/6J mice (procurement at Changsheng Experimental Animal Company, Liaoning, China) weighing from 16 to 20 g were accommodated under laboratory conditions (controlled temperature 25°C ± 2°C, relative humidity 60% ± 10%, a 12-h light–dark cycle, room air circulation 12–18 times/h) with purified water and normal chow available ad libitum. The 60 mice were maintained throughout the 12-week experimental period after being randomly grouped (n = 10), each group was as follows: (1) blank group (physiological saline, 7.5 mL/kg BW, oral gavage); (2) AL group (alcohol solutions 52% [v/v], 7.5 mL/kg BW, oral gavage); (3) AD group (ALD model [AL] group + AD 0.05, 0.1, 0.2 mg/g BW); and (4) silymarin group (AL group + silymarin 0.1 mg/g BW). Twelve days later, all animals were sacrificed by anesthesia with a lidocaine (0.2 mg/g BW), and heart blood and liver were examined by serum biochemical values testing and histopathological analysis.
Biochemical values of serum, liver lipids, lipid peroxidation, and antioxidant capacities in mice livers
According to manufacturers' instructions as described previously, 18 biochemical values of serum, the levels of liver lipids, lipid peroxidation, and antioxidant capacities in mice livers were determined using liver biochemical assay kits (Jiancheng Corporation, Nanjing, China).
Liver histopathology, immunohistochemical analysis for liver NF-κB and TNF-α, and reactive oxygen species staining
Liver tissues were fixed in 10% formalin. Then, under an Olympus BX50 light microscope (Olympus Corp., Tokyo, Japan), the pathological morphology and the expression of hepatocytes' NF-κB and TNF-α of the hepatic sections were observed by hematoxylin–eosin and immunohistochemical staining (antibodies' procurement at Abcam, Inc., Cambridge, MA, USA). The mentioned experimental results were collected by the HMIAS-2000 medical imaging system. Dihydroethidium (DHE; Beyotime Biotechnology, Jiangsu, China) staining on the frozen hepatic sections was studied under a confocal laser scanning microscope (TCS-SP2 Laser Scanning Confocal Microscope; LEICA, German).
Western blot assays
Tissue samples were weighed, and 4°C RIPA lysis buffer (C1051; Applygen Technologies, Inc., Beijing, China) was added at a proportion of 1:100 for adequate cracking and then homogenization. After centrifugation, a bicinchoninic acid protein kit (Applygen Technologies, Inc.,) was used to assay the total protein content of the samples, and the protein concentration was adjusted to be the same according to the results. All of these steps were performed on ice.
The Western blotting test procedure was as follows: 20 μL of the protein and loading buffer was added per lane. Sodium dodecyl sulfate polyacrylamide gel electrophoresis was applied to the electrophoretical separation of proteins. A Trans-Blot SD Semi-Dry Electrophoresis Transfer Cell (Bio-Rad, USA) was applied to transfer the protein bands on a polyvinylidene difluoride (PVDF) membrane (Millipore, USA). Subsequently the membrane was blocked with 5% defatted milk and then incubated overnight with a primary antibody at 4°C. Finally, the PVDF membrane was incubated with secondary antibodies for 60 min at room temperature after being washed with tris-buffered saline tween-20 buffer three times for 10 min each time. The electrogenerated chemiluminescence reagent (Millipore) was used for 5 min for development, and a gel imaging system was used to observe and photograph the membrane. Primary antibodies employed in the mentioned steps are as follows: NF-κB, TNF-α, and glyceraldehyde-3-phosphate dehydrogenase (Abcam, Inc., Cambridge, MA, USA).
Statistical analysis
All values are expressed as the mean ± SEM, and the statistical significance at the probability level of less than 0.05 was set for all tests. All data were statistically analyzed using SPSS 24.0 (IBM Corp., Armonk, NY, USA). Multigroup comparisons of the means were performed by one-way analysis of variance with post hoc contrasts tested by the least significance difference test.
Results
Effects of AD on body weight, liver weight, and liver index
At the end of the 12-week experiment, no significant difference was found in average BW among all groups (Fig. 1A, P > .05). However, compared with those of the blank group, the AL group showed a significant increment in both liver weight (Fig. 1B) and liver index (Fig. 1C, P < .05). Chronic administration with AD effectively improved the general status of liver, especially at the concentration of 0.2 mg/g BW of AD.

Effects of AD on the general status of the mice given ethanol. The levels of
Effects of AD on biochemical values, liver lipids, lipid peroxidation, and antioxidant capacities in the ALD mice
The levels of serum aminotransferase (aspartate transaminase [AST], alanine transaminase [ALT], gamma glutamyl transferase [γ-GT]) of the ALD mice were all significantly higher (Fig. 2A–C; P < .05) than those of the blank group. In the AD groups, the levels of serum aminotransferase were significantly lower than in the AL group (P < .05). In addition, contents of total cholesterol (TC), triglyceride (TG) (Fig. 2D, E), and malondialdehyde (MDA) (Fig. 3A) were significantly increased (P < .05) in the AL group, whereas superoxide dismutase (SOD) and glutathione (GSH) (Fig. 3B, C) of the liver tissue were significantly decreased (P < .05).
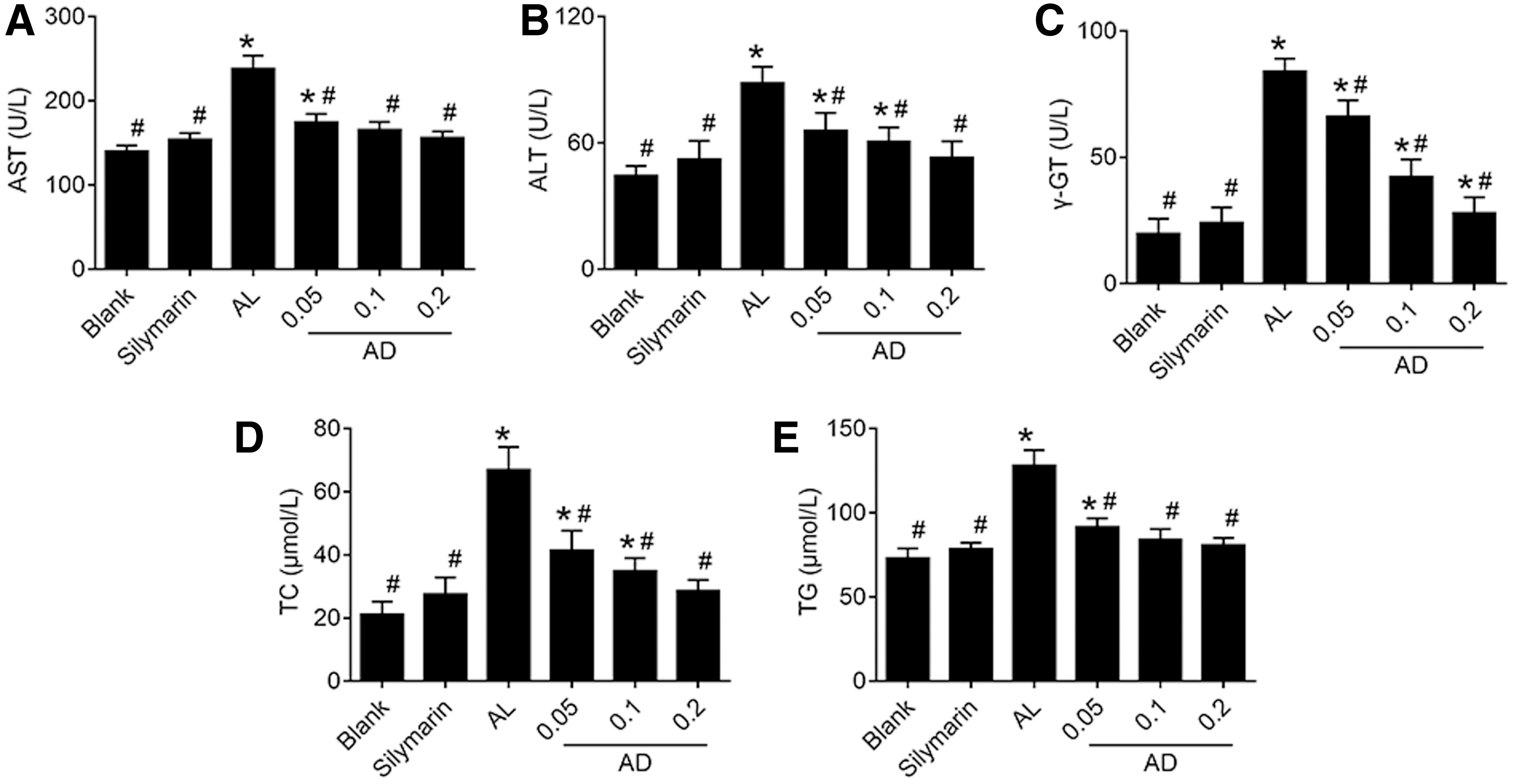
Effects of AD on serum transaminases and liver lipids in the mice given ethanol. The serum
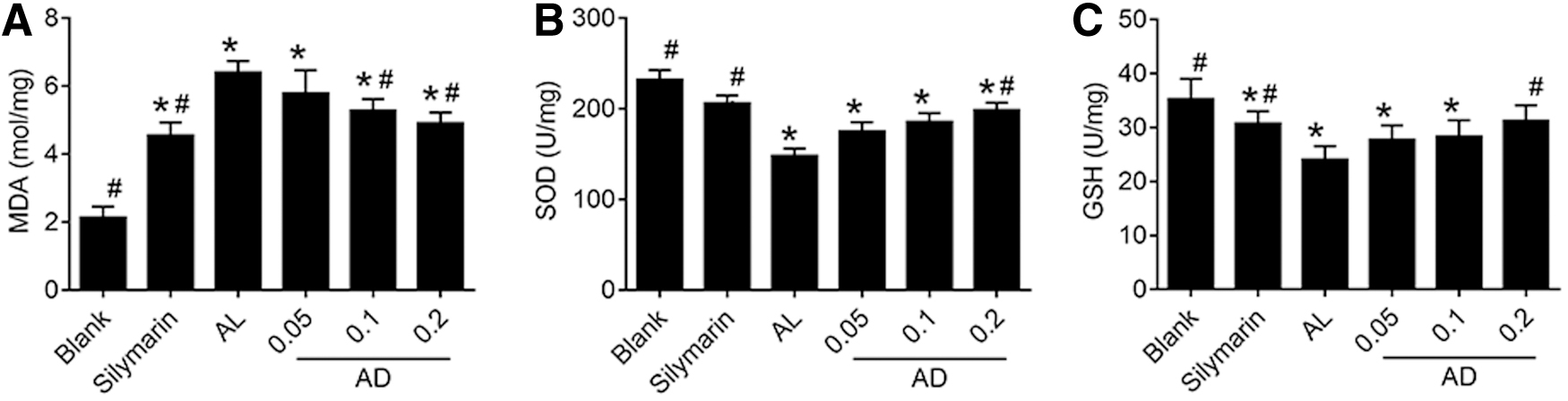
Effects of AD on lipid peroxidation and antioxidant capacities in the mice given ethanol. The levels of
Effects of AD on oxidative stress and histological findings in the liver of the ALD mice
The red fluorescence produced by DHE staining reflects the intensity of oxidative stress in tissues (Fig. 4A, B), and was significantly enhanced in the AL group mice (P < .05). We confirmed that the intensity of oxidative stress in liver was significantly decreased in AD-treated mice.
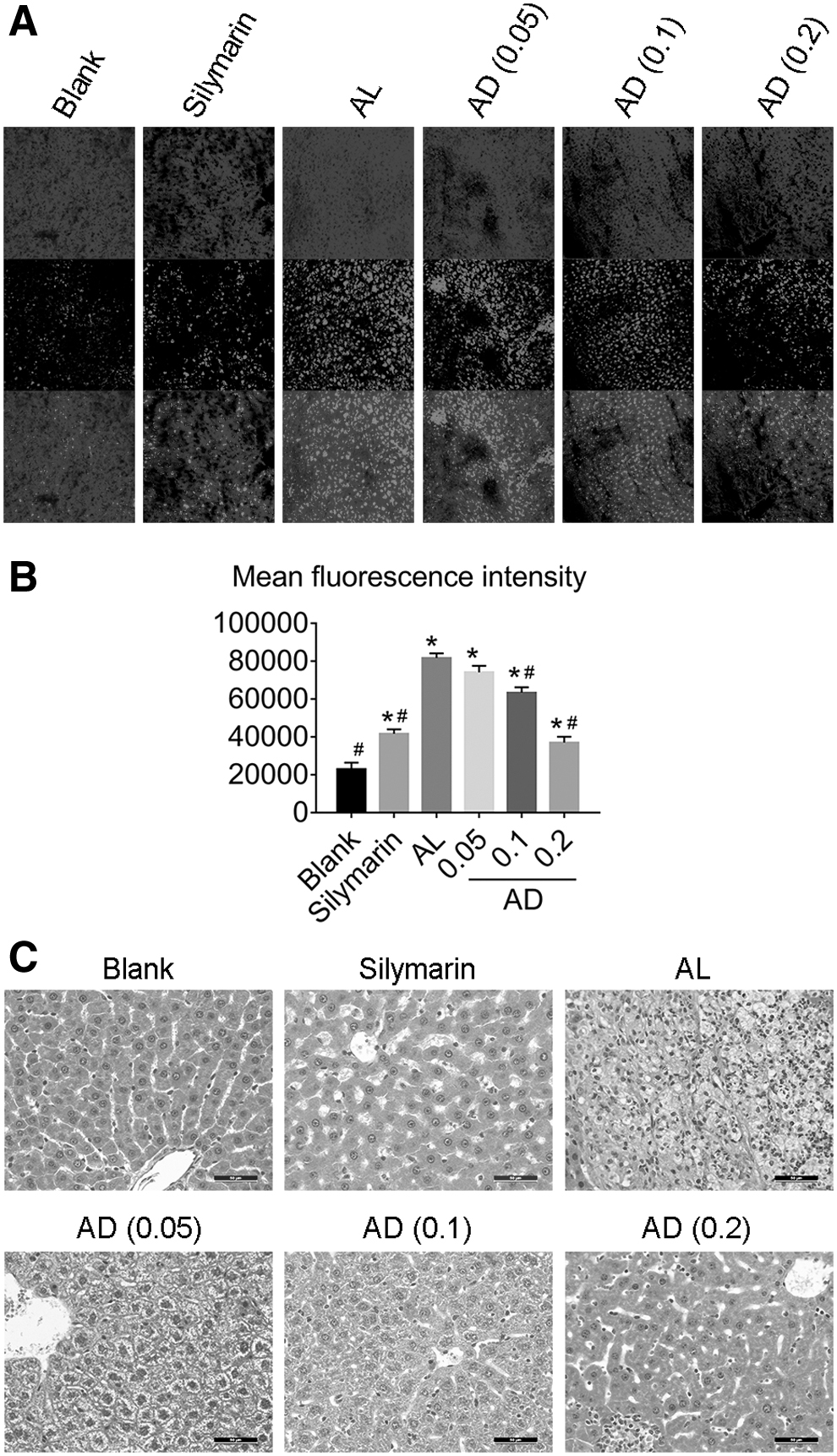
Effects of AD treatment on oxidative stress and histological findings in the liver of mice chronically fed ethanol.
In the pathological results, in particular, it can be found directly that a large number of inflammatory cells and fatty vacuoles infiltrated the AL group liver sections (Fig. 4C). In addition, fibrosis had replaced the hepatic lobules and hepatic cord structures that had been largely destroyed. The degree of damage to liver cells in the AD-administered mice (0.2 mg/g BW) was lighter than the AL group due to the presence small fat vacuoles, infiltration by a few inflammatory cells, and no fibrosis change.
Effects of AD on immunohistochemical analysis for NF-κB and TNF-α in the liver of ALD mice
The amounts of NF-κB and TNF-α (Fig. 5A, B) were increased in ethanol-administered mice liver, whereas the NF-κB and TNF-α contents in the ALD mice treated with 0.2 mg/g of AD were less.

Effects of AD on the expression of inflammation-associated proteins induced by chronic exposure to ethanol. Immunohistochemical staining of
Effects of AD on the expression of NF-κB and TNF-α proteins induced by chronic exposure to ethanol
After 12 weeks of alcohol administration, significantly higher hepatic NF-κB and TNF-α protein expression levels were detected in the AL group than those of the blank group (Fig. 5C–F; P < .05). At the same time, mice treated with AD had significantly lower expression levels of NF-κB and TNF-α compared with AL group (P < .05).
Discussion
In this study, ethanol administered by oral gavage was used to induce an animal model of ALD. The results showed that the biochemical indexes of liver function and lipid accumulation were significantly changed in the model group after alcohol treatment, and the liver tissue histological findings showed pathological changes, all of which illustrated the biochemical and pathological features of ALD, suggesting the preliminary establishment of the model. 11 In addition, inflammation and a high oxidative stress level in ALD mice were observed. Compared with the blank group, hepatic contents of MDA and reactive oxygen species (ROS) in AL group were significantly enhanced, whereas SOD and GSH were decreased. On the contrary, we found that AD could decrease serum transaminase (AST, ALT, and γ-GT) and liver lipid accumulation (TC, TG), reverse the increase of MDA content, ROS accumulation, and elevate the levels of antioxidants (SOD, GSH) in ALD mice. We observed that 0.2 mg/g AD could significantly attenuate ALD in the ethanol-induced mouse model. The histopathological image also confirmed this benefit. The release of ALT, AST, and γ-GT is widely used as a biomarker to evaluate hepatocellular damage. And lipid accumulation in hepatic cells could be aggravated by inflammatory stress induced by ALD. As aforementioned, inflammatory cytokine production is regulated by activation of NF-κB, which subsequently promotes the transcription of TNF-α, and inflammasomes, which, in turn, promote the maturation of proinflammatory cytokines. 19 Studies have shown that alcohol can induce the infiltration of neutrophils in liver tissues. 20 As a result, neutrophils release large amounts of ROS, which lead to an increase in oxidative stress that is partly responsible for alcohol-induced liver dysfunction and decreased antioxidant (SOD, GSH). 21 Oxidative stress and lipid peroxidation induced by ethanol are considered to be important factors that contribute to hepatocyte injury. 22,23 Together, these data suggest that AD may protect hepatocytes from alcohol-induced injury and exert antioxidant effects through attenuating oxidative stress. To explore the potential mechanism of AD in alleviating oxidative stress caused by alcohol, we observed the effects of AD on the expression levels of NF-κB and TNF-α.
The view that long-term consumption of alcohol can activate NF-κB in liver tissues has been demonstrated by previous research. 24,25 The promoter sequences of various inflammatory factors can bind to the activated NF-κB after it enters the nucleus. Then they promote the transcription of pro-inflammatory cytokines (TNF-α, IL-1, IL-2, IL-6, etc.), adhesion molecules, and NO synthase, resulting in inflammation, and even cell necrosis and apoptosis. 19,26 Moreover, proinflammatory cytokines can produce numerous free radicals in the process of alcohol metabolism. 27,28 When free radicals are generated beyond the body's scavenging capacity, they can inhibit fatty acid oxidation, trigger liver lipid accumulation, and lead to lipid peroxidation of the liver cell membrane, causing hepatocyte damage. At the same time, hepatic injury increases the expression of inflammatory cytokines, and excessive oxidative stress can further stimulate the release of endotoxin and inflammatory mediators. 27 These substances, in turn, react with cells, increase the nuclear translocation of NF-κB, and then cause an inflammatory cascade amplification, aggravating hepatic damage. TNF-α is the most critical inflammatory mediator and plays a positive feedback role in the activation of NF-κB. TNF-α is not only an activator of NF-κB, but also is a product of its activation. 29 TNF-α, on the one hand, has a direct cytotoxic effect, causing hepatocyte necrosis, on the other hand, it can cause microcirculation disorders leading to hepatocyte necrosis. At the same time, TNF-α and other inflammatory factors such as IL-6 stimulate each other, and participate in the cascade amplification reaction.
Studies have shown that AD can inhibit the combination of NF-κB and promoter region of downstream genes through covalent binding with the 62nd cysteine of the subunit of NF-κB p50 and thus restrain inflammatory reactions. 30 Western blot results further confirmed that the expression levels of NF-κB and TNF-α protein in the AL group were significantly increased, which was consistent with the previous views. 24,25 Whereas the levels of NF-κB and TNF-α in liver tissue of each dose group of AD were significantly decreased after administration and showed a dose–response relationship. To further confirm these results, immunohistochemistry was performed on liver tissues. Consistent with the Western blotting, results of immumohistochemical staining showed that NF-κB and TNF-α expression levels were increased in livers from model mice compared with those in normal mice, and were markedly decreased by AD treatment. Taken together, these results suggested that AD may exert hepatoprotective effects through inhibiting NF-κB and TNF-α. There are two types of NF-κB signaling pathway: canonical and noncanonical. Both require activation through posttranslational modifications of NF-κB. The canonical pathway activation predominantly occurs through B cell receptors, T cell receptors, toll-like receptor 4, IL-1, or receptors for TNF, whereas the noncanonical pathway involves receptor activation of cluster of differentiation 40 ligand, B cell activation factor, or lymphotoxin-B. 31 Each signal pathway mentioned does not exist independently. They can interact with each other and jointly regulate the self-protection function of the body. Therefore, the possible mechanisms of AD inhibiting the increase of NF-κB induced by ethanol need to be further studied in detail.
At present, the disease burden of ALD is increasing fast, especially in Asian countries such as China, Korea, and India. 1,32 Because the mechanism of ALD is complex and alcohol can induce many complications, there are limited ideal treatments for alcohol-related diseases. 33 –35 As a major active constituent isolated from A. paniculate, AD has proven pharmacokinetic properties and safety, has good antioxidative stress and anti-inflammatory activities, and has been used in many Asian countries to alleviate various inflammatory symptoms. 14,15 Whether AD can inhibit ethanol-induced oxidative stress and relieve liver damage in mice had not been reported. Therefore, we made a model of ALD to observe and use AD intervention, hoping to find effective drugs for treating ALD. The results of this study showed that AD can relieve ALD injury by its strong anti-inflammatory activation. Furthermore, we speculate that AD can be used in the clinical treatment of ALD in a relatively short time, which can avoid the limited clinical success faced by the development of new drugs.
In summary, this study revealed some molecular mechanisms of AD in the treatment of ALD, which can alleviate liver pathological injury and oxidative stress by decreasing the expression of NF-κB and TNF-α. Therefore, AD has potential as an alternative intervention that can ameliorate ALD.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This article was supported by the National Natural Science Foundation of China (Grant No. 81803218) and Jilin Province Health Science and Technology Capacity Improvement Project (Grant No. 2019Q036). The sources of funding did not participate in the following aspects: the design and implementation of this article; data collection, processing, analysis, and interpretation; the writing, examination, or approval of the article; and the decision to submit the article for publication.