
Abstract
Endotoxic shock exhibits a considerably high mortality risk. It is defined as a systemic inflammatory response syndrome caused by a microbial infection. Radix Isatidis has anti-inflammatory, antiviral, and antipyretic effects and is used worldwide. This study investigated the antiendotoxin sepsis effects of an aqueous R. Isatidis extract (RIE) and explored the possible pharmacological molecular mechanisms. Male C57BL/6J mice were intravenously injected with 15 mg/kg lipopolysaccharide (LPS) to induce endotoxic shock. The results demonstrated that the survival rate of mice pretreated with RIE increased, and LPS-induced liver and lung damage were reduced by inhibiting inflammation. For elucidating detailed molecular mechanisms, we focused on LPS-induced transcription factors: nuclear factor-κB (NF-κB) and interferon regulatory factor 3 (IRF3). Our results demonstrated that the protective effects of RIE were strongly dependent on IRF3-induced interferon-β, not on NF-κB-induced tumor necrosis factor-α and interleukin-1β. In addition, RIE suppressed the phosphorylation of IRF3, not NF-κB. In conclusion, this study revealed the antiendotoxic properties of RIE on LPS-induced sepsis and provided mechanistic evidence for the beneficial effects of RIE.
Introduction
Sepsis is defined as a systemic inflammatory response syndrome caused by a microbial infection. 1 Endotoxic shock is the most severe form of sepsis and exhibits a considerably high mortality risk, making it the most common cause of death in intensive care units. 2 –4 The current treatment for sepsis relies on supportive treatment, with no effective treatment that has been reported. 5 –7 Uncontrolled inflammatory response is a crucial mechanism for the development of bacterial sepsis causing septic shock. Overproduction of mediators of inflammation and immune responses results in pathological outcomes of sepsis, including tissue injury, multiorgan failure, and death. 8 –10
Lipopolysaccharide (LPS), a major component of the outer membrane of Gram-negative bacteria, plays a central role in the sepsis, inducing excessive or unwanted inflammation and finally fatal endotoxic shock. 11 –13 Seminal studies have shown that LPS binds to toll-like receptor 4 (TLR4) to activate two transcriptional factors: nuclear factor-κB (NF-κB) and interferon regulatory factor 3 (IRF3). 14,15 NF-κB controls the expression of proinflammatory cytokines such as tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β). 16,17 Meanwhile, IRF3 induces type I interferon-β (IFN-β) expression. 18,19 Thus, the mRNA expressions of these genes represent the activation of transcriptional factors.
Radix Isatidis is the dry root of Isatis indigotica Fort. (Fam. Brassicaceae), which is a cruciferous plant of the genus Isatis. It is one of the most important traditional Chinese medicines with anti-inflammatory, antiviral, and antipyretic effects; it has been used to remove heat and toxins, cool the blood, and clear the throat in East Asian countries for thousands of years. 20 –25 Although aqueous R. Isatidis extract (RIE) is clinically used for sepsis treatment, its main use is against viral or bacterial infections, including viral pneumonia, mumps, flu, and common cold 21 ; thus, few studies have focused on its antisepsis effects. In addition, only a few molecular based studies on R. Isatidis have been reported, and most reports have used the chloroform or methanolic R. Isatidis extracts. 20,26 However, the main components of chloroform or methanolic extracts poorly dissolve in water; therefore, using RIE to determine its detailed molecular mechanisms is essential for understanding the therapeutic effects of R. Isatidis. In this study, we evaluated the effect of RIE in an LPS-induced bacterial sepsis mouse model and RAW 264.7 cells and studied its pharmaceutical functions and molecular mechanisms.
Materials and Methods
Chemicals and antibodies
LPS, uridine, and guanosine were purchased from Sigma-Aldrich (St. Louis, MO, USA). Adenosine was purchased from the National Institute for the Control of Pharmaceutical and Biological Products (Beijing, China). R,S-goitrin and o-aminobenzoic acid (purity >98%) were provided by the Shanghai R&D Centre for Standardization of Traditional Chinese Medicines (Shanghai, China). Syringic acid, benzoic acid, salicylic acid, and 4(3H)-quinazolone were purchased from Adamas Reagent, Ltd. (Shanghai, China).
Anti-NF-κB p65 and anti-rabbit immunoglobulin G–horseradish peroxidase (HRP) conjugates were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Anti-β-Actin, anti-LaminB1, anti-IRF3, anti-p-IRF3 (Ser-396), anti-tank-binding kinase 1 (TBK1), anti-p-TBK1 (Ser-172), anti-I-kappaB kinase ɛ (IKKɛ), anti-p-IKKɛ (Ser-172), and anti-NF-κB pp65 (Ser-536) were purchased from Cell Signaling Technology (Beverly, MA, USA).
RIE preparation
Raw R. Isatidis was purchased from the Shanghai Yang-He-Tang Traditional Chinese Medicine Company (Shanghai, China). R. Isatidis was extracted as follows: R. Isatidis (200 g) was immersed in 2000 mL of distilled water for 1 h, followed by reflux extraction at 100°C for 30 min. The resulting solution was filtered through three gauze sheets, and the residues were extracted again under the same condition for 20 min. After filtration, the extract was condensed to make the concentration equivalent to 1.67 g herb/mL, filtered through 0.22-μm sterile membranes, labeled RIE, and finally, kept at 4°C.
Quality control of RIE
To control RIE quality, a high-performance liquid chromatography (HPLC) method was performed to analyze chemical characterization. The analyses were performed using an Agilent 1100 system (Agilent Technologies, Hewlett-Packard-strasse-8, Germany) equipped with a quaternary gradient pump. The chromatographic column was Agilent USA SB-C18 (4.6 × 250 mm2, 5 μm). UV detection was set at full wave scanning. The chromatographic condition was modified according to a previous publication 27 : solvent A, methanol, and solvent B, water. A mobile-phase gradient was used with varying percentages of A in B as follows: initial concentration, 3% A, 0–3 min; 3–7% A, 3–13 min; 7–10% A, 13–15 min; 10% A, 15–20 min; 10–13% A, 20–24 min; and 13% A, 24–40 min. The mobile-phase flow rate was 0.8 mL/min, and the injection volume was 10 μL.
Uridine, guanosine, R,S-goitrin, and adenosine were dissolved in water and used as controls. Otherwise, 4(3H)-quinazolone, syringic acid, o-aminobenzoic acid, salicylic acid, and benzoic acid were dissolved in methanol and used as controls. The mobile phase consisted of 0.2% acetic acid in water (C) and acetonitrile (D). A mobile-phase gradient was used with varying percentages of D in C as follows: initial concentration, 0–20% D, 0–10 min; 20–40% D, 10–25 min; and 40% D, 25–30 min. The mobile phase was set at a flow rate of 1 mL/min, and the injection volume was 10 μL.
Mice and endotoxin model establishment
Male C57BL/6J mice (age, 6–8 weeks; weight ∼18–22 g), were obtained from the Shanghai SLAC Laboratory Animal (Shanghai, China). All efforts were made to minimize animal suffering and to reduce the number of animals used in the experiments. The animal protocol in this study was performed in accordance with the National Institute of Health Guide for the Care and use of Laboratory Animals and approved by the Animal Ethics Committees of Shanghai University of Traditional Chinese Medicine (ethics number: SZY201705008). Animals were housed at a constant room temperature (25°C) and relative humidity (70%) under a 12-h light–12-h dark cycle. Food and tap water were provided ad libitum.
Male C57BL/6J mice were randomly divided into three groups: untreated, LPS treated, RIE pretreated sepsis. In the RIE-pretreated sepsis group, mice were pretreated with 7.5 g/kg RIE through intraperitoneal injection for 1 h and then injected intravenously with 15 mg/kg LPS to induce experimental sepsis. Mice in the LPS group were administered equivalent volumes of 0.9% NaCl (vehicle) for 1 h before LPS injection. In the untreated group, mice were administered with equivalent volume of 0.9% NaCl.
Cell culture and stimulation
Murine macrophage RAW 264.7 cells were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China) and were cultured in Dulbecco's modified Eagle medium (Thermo Fisher Scientific, Waltham, MD, USA) supplemented with 10% fetal bovine serum (Thermo Fisher Scientific), 100 U/mL penicillin (Thermo Fisher Scientific), and 100 g/mL streptomycin (Thermo Fisher Scientific) at 37°C with 5% CO2.
RAW 264.7 cells were plated at a density of 5.0 × 104 cells/well in 24-well polystyrene plates (Eppendorf, Hamburg, Germany) and preincubated for 24 h. The cells were then pretreated with 30 or 60 μg/mL RIE for 30 min before adding 100 ng/mL LPS. After incubation for another 4 h, the cells were collected for reverse transcription-quantitative polymerase chain reaction (RT-qPCR) analysis.
RAW 264.7 cells were plated at 5.0 × 105 cells/dish in 6-cm polystyrene dishes (Eppendorf) and preincubated for 24 h. Then, the cells were pretreated with RIE (100 μg/mL) for 30 min before adding 1 μg/mL LPS. After incubation for another 10, 30, 60, and 120 min, the cells were collected for Western blot analysis.
Measurement of body temperature and survival analysis
For body temperature measurement, we monitored rectal temperature for 8 h. The body temperature of mice was measured once per hour by using a human electronic thermometer; the thermometer was gently inserted 1.0–1.5 cm into the rectum using petroleum jelly. For survival analysis, general conditions and mouse mortality were observed up to 7 days.
Histological analysis
The livers and lungs were collected and fixed with 4% formaldehyde and then embedded with paraffin. Serial 5-μm-thick sections were cut using a microtome (Leica, Heidelberg, Germany) and then stained with hematoxylin and eosin. Light microscopy photographs were taken using Olympus DX45 (Tokyo, Japan).
RT-qPCR analysis
Total RNAs from different experimental groups were extracted using an RNA Faster200 reagent (Fastagen, Shanghai, China), according to manufacturer's instructions. RNA was reverse transcribed to generate first-strand cDNAs by using the PrimeScript™ RT Master Mix Kit (Takara Bio, Shiga, Japan), and reaction conditions were 37°C for 15 min and 85°C for 5 s. The qPCR analysis was performed using a SYBR Premix Ex Taq™ (Tli RNaseH Plus) Kit (Takara Bio), according to manufacturer's instructions. All the reactions were performed on a 6000 Real-Time PCR System (Thermo Fisher Scientific), and the relative quantity was quantified using the 2−ΔΔCt method. The cycle threshold values of the target genes were normalized to that of glyceraldehyde 3-phosphate dehydrogenase (Gapdh) from the same sample. The primers are listed in Table 1.
Primers Used in This Study
ELISA analysis
Blood samples were centrifuged at 1,200 g for 15 min. Serum TNF-α, IL-1β, and IFN-β concentrations were measured using ELISA kits (Boster Biotech, Wuhan, China for TNF-α and IL-1β; Cusabio, Wuhan, China for IFN-β), according to manufacturers' instructions.
Western blot analysis
RAW264.7 cells were washed with cold phosphate-buffered saline (PBS), scraped off, and harvested. For obtaining total protein, cell extracts were prepared using whole cell lysate buffer (1 M HEPES [pH 7.7], 5.94 M NaCl, 0.1 M MgCl2, 0.5 M EDTA [pH 8.0], 10% TritonX-100, 10 mM pyrophosphate, and protease inhibitors) and chilled on ice for 30 min. The lysates were centrifuged at 18,000 g for 10 min at 4°C.
For obtaining nuclear protein, cell extracts were added to 200 μL buffer A (1 M HEPES [pH 7.9], 2 M KCl, 0.25 M EDTA [pH 8.0], 0.25 M EGTA [pH 8.0], 10 mM pyrophosphate, and protease inhibitors), chilled on ice for 10 min, and vortexed every 5 min. Subsequently, 25 μL of 10% NP-40 was added to the extract; it was then resuspended, chilled on ice for 20 min, and then centrifuged at 1,500 g for 5 min at 4°C. The supernatant was then removed and 100 μL buffer C (1 M HEPES [pH 7.9], 4 M NaCl, 0.25 M EDTA [pH 8.0], 0.25 M EGTA [pH 8.0], 10 mM pyrophosphate, and protease inhibitors) was added; this solution was chilled on ice for another 30 min, vortexed every 5 min, and centrifuged at 18,000 g for 10 min at 4°C.
Protein concentration was analyzed using a BCA Protein Quantification Kit (Yeasen Biotech, Shanghai, China). Cell protein was separated through 10% SDS-PAGE and transferred on to polyvinylidene difluoride membranes (Merck Millipore, Billerica, MA, USA). These membranes were blocked for 2 h at room temperature in SuperBlockTM Blocking Buffer in PBS (Thermo Fisher Scientific) on a rotary shaker, followed by washing with PBS Tween-20 (PBST). The membranes were subsequently incubated overnight at 4°C with the primary antibody. After being washed thrice with PBST, the membranes were incubated with the secondary HRP-conjugated antibody at room temperature for 1 h. The membranes were again washed thrice with PBST and subsequently developed using Immobilon Western Chemiluminescent HRP Substrate (Merck Millipore).
Statistical analysis
Statistical analysis was performed using GraphPad Prism (version 7; GraphPad, La Jolla, CA, USA). All the assay values are given as mean ± SD. Differences among multiple groups were analyzed using one-way analysis of variance, followed by Tukey post hoc test. P < .05 was considered statistically significant.
Results
HPLC analysis
To investigate the quality of RIE used in this study, the quantity of R,S-goitrin was determined through HPLC. As shown in Figure 1A (peak number 3), R,S-goitrin content in the RIE sample was 0.034% (w/w), which complies with the Chinese Pharmacopoeia (2015 edition, Volume I) standards. Uridine, guanosine, and adenosine are active compounds in RIE; therefore, their quantities were also determined. The contents of uridine (peak number 1), guanosine (peak number 2), and adenosine (peak number 4) in RIE were 0.035% (w/w), 0.054% (w/w), and 0.038% (w/w), respectively. In addition, 4(3H)-quinazolone, syringic acid, o-aminobenzoic acid, salicylic acid, and benzoic acid, which were isolated from the chloroform R. Isatidis extract, were reported to have antiendotoxin effects. 28 –32 These compounds (peak numbers 5–9) were below the detection limit in our sample (Fig. 1B).

Representative HPLC spectra of the RIE sample. The upper and lower figures show the chromatograph of the RIE sample and standard, respectively.
Protective effects of RIE in the LPS-challenged sepsis mice model
We investigated antiendotoxic effects of RIE using the LPS-challenged lethal sepsis C57BL/6J mice model. 33 –35 As shown in Figure 2A, normal mice challenged with a lethal dose of LPS were dead within 2 days. The survival rates of RIE-pretreated mice were obviously improved to 80% at the end of the observation period. We also measured the body temperature and found that RIE prevented LPS-induced hypothermia in mice (Fig. 2B).
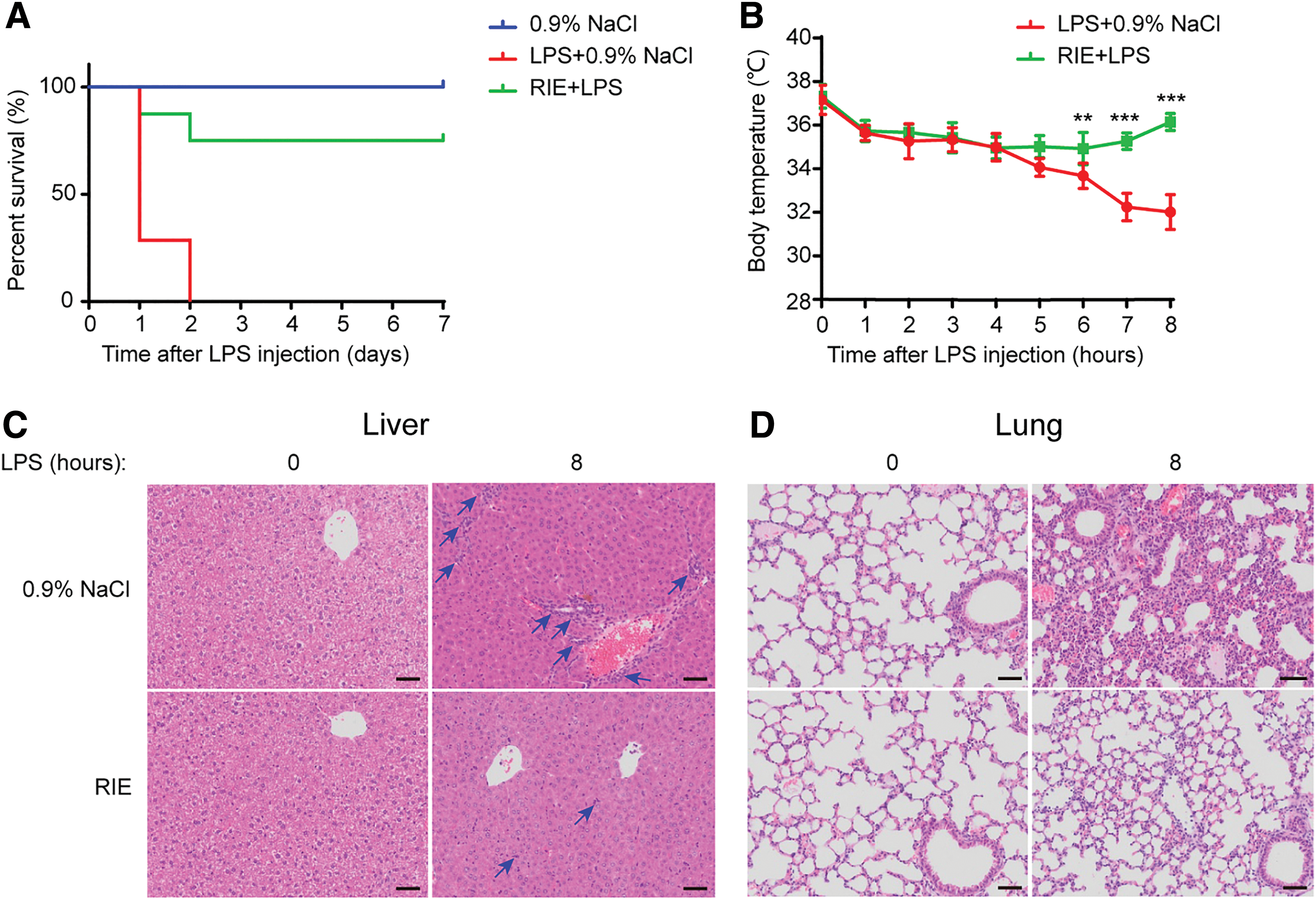
Protective effects of RIE in LPS-challenged mice. Mice were peritoneally injected with 0.9% NaCl or RIE (7.5 g/kg) 1 h before LPS (15 mg/kg) intravenous injection (n = 10).
Liver and lung damage in LPS-challenged mice were evaluated through pathological analysis (Fig. 2C, D). In the normal group, the liver structures were clear, the liver cords were orderly, and inflammatory cell infiltration was not observed in the livers. After LPS injection, the hydropic degeneration of hepatocytes and the aggregation of inflammatory cells (the blue arrow) were observed at the portal areas and in the hepatic sinusoids (Fig. 2C, upper right). In the RIE-pretreated group, the liver had a small amount of inflammatory cell infiltration, and almost no morphologic changes in hepatocytes were observed (Fig. 2C, lower right). Similarly, severe lung damage was shown in the LPS-injected group. After LPS injection, alveolar wall thickness increased and the number of pulmonary alveoli decreased (Fig. 2D, upper right). In the RIE-pretreated group, lung damage was reduced in comparison to the LPS-injected group (Fig. 2D, lower right).
Because RIE reduced the aggregation of inflammatory cells in the livers of the mice, we further investigated whether RIE could reduce the expression of inflammatory-related genes. The nitric oxide synthase gene (Nos2) encodes inducible NOS2 and is often upregulated under acute inflammation to produce nitric oxide. 36,37 Interleukin-6 (IL-6) is a pleiotropic cytokine that strongly influences the inflammatory response and acts as a major effector during acute inflammation. 38 Thus, we measured the expression of Nos2 and Il-6 in the LPS-challenged septic mice livers. Nos2 and Il-6 expression, which were increased in the LPS-challenged mice, were significantly decreased in the RIE-treated group (Fig. 3A). Similar results were obtained in the mice lungs (Fig. 3B).

Inhibition of Nos2 and Il-6 in RIE-pretreated LPS-challenged mice. Mice were peritoneally injected with 0.9% NaCl or RIE (7.5 g/kg) 1 h before LPS (15 mg/kg) intravenous injection for 8 h. Relative mRNA levels in the liver
Taken together, RIE has protective effects against mice septic death because of the inhibition of inflammatory responses.
Effects of RIE on NF-κB-related genes and proteins on the mouse model
On LPS stimulation, its receptor TLR4 is activated. Triggering of the TLR4 pathway leads to the activation of NF-κB and IRF3 and promotes the subsequent production of inflammatory and immune genes. 15 –18 RIE inhibited the infiltration of inflammatory cells and the expression of inflammatory genes; thus, we elucidated the detailed molecular mechanisms of its protective effects.
We first investigated whether RIE could inhibit the expression of NF-κB-related genes, Tnf-α and Il-1β. As shown in Figure 4A, LPS induced Tnf-α and Il-1β expression at 1, 4, and 8 h in the mouse livers. However, the preadministration of RIE inhibited their expression only in the 4-h group, not in the 1- and 8-h groups. Similar results were shown in the lungs (Fig. 4B). Although RIE reduced the expression of NF-κB-related genes, the effect is transient, and the inhibition ratio is not very high. Next, the release of TNF-α and IL-1β proteins in serum was detected. RIE had almost no detectable effects on the production of these cytokines (Fig. 4C, D). Collectively, RIE had little effects on reducing the expression of LPS-induced NF-κB regulating genes and proteins.

RIE weakly inhibited NF-κB-related genes and proteins in LPS-challenged mice. Mice were peritoneally injected with 0.9% NaCl or RIE (7.5 g/kg) 1 h before LPS (15 mg/kg) intravenous injection.
RIE significantly inhibits IRF3 signaling in the mouse model
We investigated whether RIE could block the LPS-induced activation of IRF3. The expression of IFN-β, which is the typical gene induced by IRF3, was determined. As shown in Figure 5A and B, Ifn-β was increased in the LPS-challenged group both in the liver and lung. The preadministration of RIE significantly inhibited the expression of Ifn-β at all time points. Next, we detected the IFN-β protein level in mice serum. Correlated to the mRNA level, IFN-β induced by LPS was suppressed in the RIE-pretreated group (Fig. 5C). In conclusion, RIE significantly blocked the activation of the IRF3 signal.
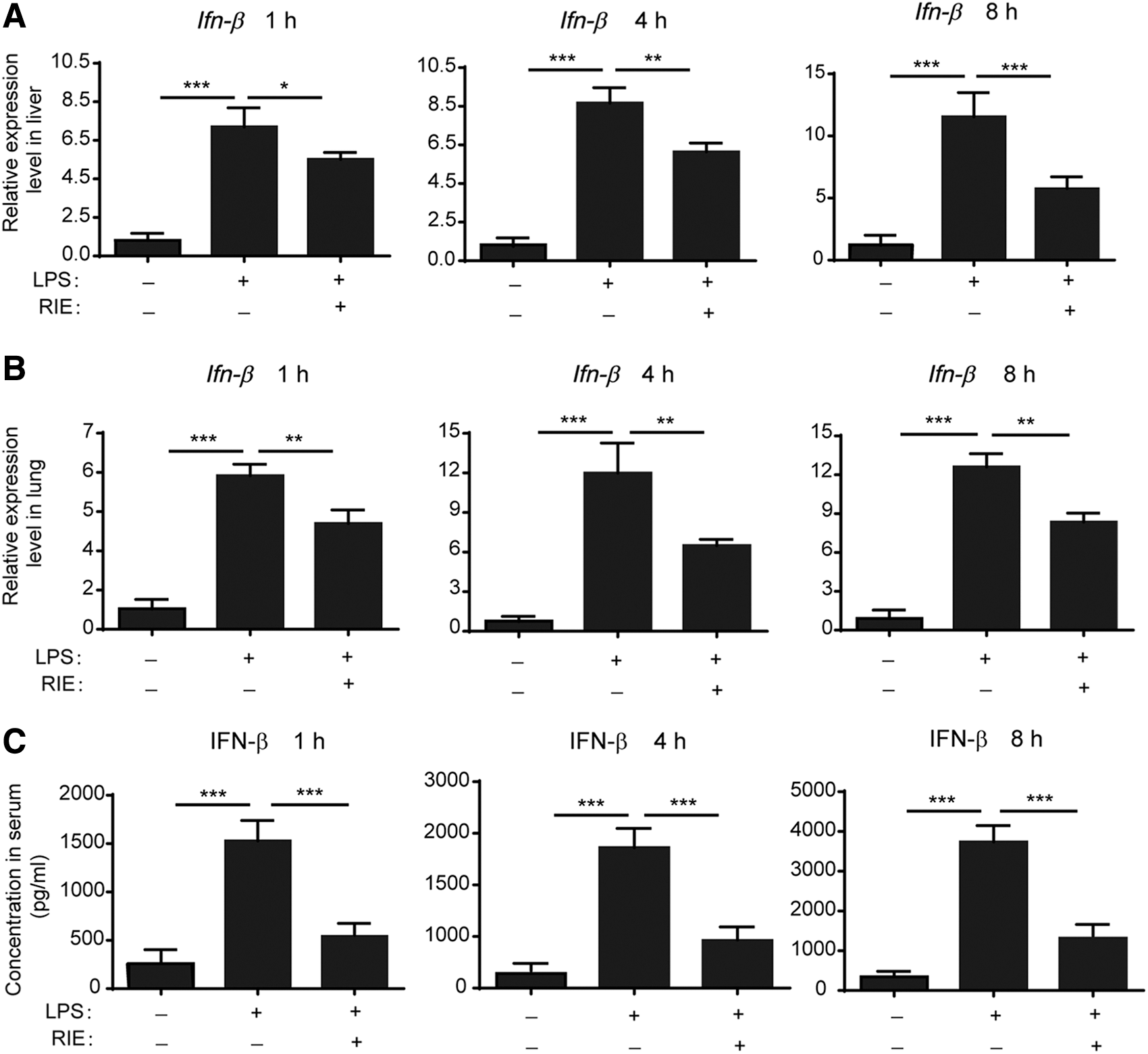
RIE significantly inhibited IFN-β in LPS-challenged mice. Mice were peritoneally injected with 0.9% NaCl or RIE (7.5 g/kg) 1 h before LPS (15 mg/kg) intravenous injection. Relative expression of Ifn-β in the liver
IRF3 inhibitory effects of RIE on LPS-stimulated RAW 264.7 cells
The above analysis was performed in vivo. To confirm the in vitro effects of RIE and elucidate the underlying molecular mechanisms, the mouse macrophage cell line RAW264.7 was used in the next experiments. RAW264.7 cells were pretreated with RIE for 30 min and then stimulated with LPS for 4 h. Consistent with the mouse model, the expression of Ifn-β, Tnf-α, and Il-1β was upregulated on LPS stimulation (Fig. 6A). The level of Ifn-β, not Tnf-α or Il-1β, was strongly suppressed in an RIE concentration-dependent manner. These results correlated with those of in vivo experiments.
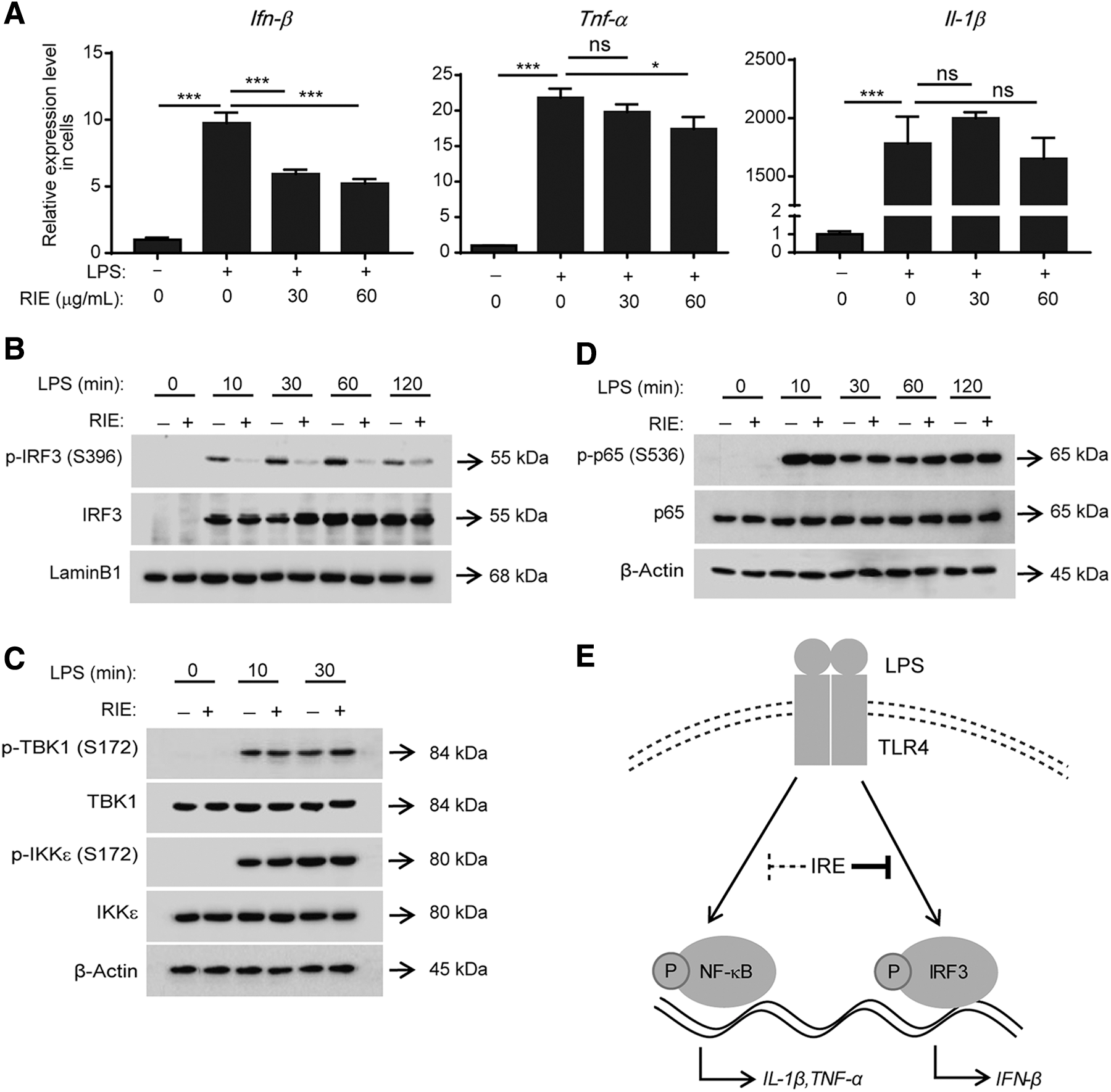
Effects of RIE on the IRF3 and NF-κB pathways in LPS-stimulated RAW 264.7 cells.
RIE mainly inhibits IRF3 signaling and has a small inhibitory effect on NF-κB signaling, both in vivo and in vitro; thus, we examined the effects of RIE on IRF3 and NF-κB activation. As shown in Figure 6B, the Western blotting analysis was performed to analyze the phosphorylation of IRF3 (Ser-396), which is the indicator for IRF3 activation, using the nucleus lysates of RAW 264.7 cells. Upon LPS stimulation, IRF3 was translocated to the nucleus and its phosphorylation was induced. 18,19 Similar to our prediction, this phosphorylation was strongly inhibited by RIE pretreatment. IRF3 phosphorylation is induced by its upstream kinases, TBK1 and IKKɛ. 18,19 RIE did not inhibit the activation of these kinases, suggesting that RIE may affect IRF3 directly (Fig. 6C).
The phosphorylation of NF-κB p65 (Ser-536) is the indicator of NF-κB p65. In contrast to IRF3, RIE did not inhibit this phosphorylation (Fig. 6D). Taken together, RIE mainly inhibited the IRF3 pathway by inhibiting IRF3 phosphorylation, not its upstream kinases.
Discussion
The overactivated immune signaling can induce severe inflammatory injury, and it is linked to poor disease outcomes and increased mortality, especially in sepsis. 39,40 Immune cytokines potentially drive local tissue damage and hypothermia, which are considered an important indicator of inflammation in rodent and human sepsis. 41 –43 In our results, LPS-induced hypothermia, inflammatory injury in tissues and mortality, and RIE pretreatment showed strong protective effects against them. RIE significantly inhibited the inflammatory response, including the expression of inflammatory cytokines Nos2 and Il-6 in the endotoxin mice, suggesting that inhibiting inflammation is the main effect of RIE in sepsis.
In our research, we focused on the NF-κB and IRF3 signaling pathways. RIE strongly reduced IRF3 signaling, including IRF3 phosphorylation and IFN-β mRNA and protein expressions at all time points. On the other hand, RIE did not inhibit NF-κB phosphorylation, however, it slightly inhibited Tnf-α and Il-1β expression at 4 h but no effect was observed at 1 or 8 h. Upon NF-κB activation, CYLD, A20, and OTULIN, which are transient negative regulators of NF-κB, are induced, and then, they inhibit NF-κB activation. 44,45 Our data demonstrated that LPS induced A20 expression from ∼2 h, and A20 expression decreased to the basal level before 8 h (data not shown). A20 expression might be induced more in RIE-pretreated cells than non-pretreated cells, therefore, RIE showed the inhibitory effects on Tnf-α and Il-1β expression at 4 h. The detailed molecular mechanisms of the transient inhibition of NF-κB genes in RIE should be elucidated by further study.
IFN-β is a main molecule that naturally regulates the innate immune signal. Upon viral infection, innate immune regulates cytokine secretion to eliminate viruses; however, when excessively produced, the cytokines may be directed against the host. 46 Pharmacological inhibition of type I IFN signaling protects mice against lethal sepsis. 47 IFN-β and its receptor IFNAR-knockout mice are protected against the lethal effects of LPS. 48 –51 Our results indicated that RIE strongly inhibited LPS-induced IRF3 phosphorylation and suppressed IFN-β expression, indicating that the suppression of IRF3-dependent IFN-β is the main effect of RIE for the protection against LPS-induced sepsis (Fig. 6). IFN-β oversecretion was shown not only in sepsis but also in many types of inflammatory diseases, 52,53 suggesting that RIE can be used for the treatment of many inflammatory diseases, which depend on the IRF3 signaling pathway.
In one study, chloroform R. Isatidis extract F022 had protective effects against LPS-induced sepsis mice. 26 By further isolation from F022, 4(3H)-quinazolone, 28 syringic acid, 29 o-aminobenzoic acid, 30 salicylic acid, 31 and benzoic acid 32 were identified as the main components providing protective effects by inhibiting LPS-induced TNF-α expression. We tried to detect the contents of these molecules in RIE using HPLC analysis; however, all the contents were below the detection limit, indicating that these components were present in very low amounts or not present in RIE (Fig. 1B) because of their hydrophobic states.
Unlike the hydrophobic constituents contained in the chloroform R. Isatidis extract, some types of water-soluble components, including uridine, guanosine, and adenosine, were detected in the RIE. 54 Uridine reduced both edema and the infiltration of leukocytes in a rat model of lung inflammation. 55 Guanosine and adenosine attenuated the nitric oxide release of macrophages. 56 In our experiments, these compounds may play a critical role in reducing the aggregation of inflammatory cells and the expression levels of Nos2. No report demonstrated that these compounds can suppress IFN-β in the sepsis mice model; therefore, further studies are required for elucidating these compounds' molecular mechanisms and understanding the whole therapeutic effects of RIE.
In addition, R. Isatidis-containing serum could downregulate IFN-β expression in respiratory syncytial virus (RSV)-infected RAW 264.7 cells. 57 Furthermore, He et al. reported that tryptanthrin B, 4(3H)-quinazolone, and R-goitrin were present for a long time in R. Isatidis-containing serum, and only 4(3H)-quinazolone inhibited both NF-κB and IRF3 activities in RSV-infected RAW 264.7 cells. 58 In our study, R,S-goitrin, but not tryptanthrin B and 4(3H)-quinazolone, was detected in our RIE sample (Fig. 1A), suggesting the presence of novel components in RIE with inhibitory effects against the IRF3-IFN-β axis. These components require further investigation.
In summary, this is the first study illustrating that RIE has protective effects on LPS-induced sepsis by inhibiting the IRF3 signaling pathway; it does not have effects on NF-κB (Fig. 6E), which is different from previous studies reporting that the chloroform R. Isatidis extract inhibited LPS-induced TNF-α expression. Although detailed studies are required, our study elucidates the effects of this traditional Chinese medicine.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was financially supported by the National Natural Science Foundation of China (Nos. 81603156 and 81573571), Excellent Academic Leaders Program of Shanghai (No. 16XD1403500), Young Eastern Scholar Program (No. QD2016038), Chenguang Program (No. 16CG49), and the programs of High Level University Innovation Team and Shanghai E-Research Institute of Bioactive Constituents in Traditional Chinese Medicine.