
Abstract
Siegesbeckia orientalis has been reported to exhibit anti-allergic, anti-infertility, anti-inflammatory, anti-rheumatic, and immunosuppressive activities. However, there are very few studies describing its stimulatory effects on exercise capacity. This study elucidated whether S. orientalis extract (SOE) standardized to kirenol content can enhance exercise endurance by increasing mitochondrial biogenesis. SOE significantly improved the running distance and time in mice fed normal diet (ND) and high-fat diet (HFD). SOE also enhanced mitochondrial biogenesis by stimulating the mitochondrial regulatory genes including peroxisome proliferator-activated receptor gamma co-activator 1 alpha (PGC-1α), estrogen-related receptor α (ERRα), nuclear respiratory factor 1 (NRF-1), and mitochondrial transcription factor A (TFAM) in the skeletal muscles of ND and HFD mice. Furthermore, SOE upregulated the AMP-activated protein kinase (AMPK)/sirtuin 1 (SIRT1)/PGC-1α/peroxisome proliferator-activated receptor delta (PPARδ) signaling pathway in the skeletal muscles of ND and HFD mice. Kirenol markedly increased adenosine triphosphate production and mitochondrial activity by stimulating the expression of markers of mitochondrial biogenesis and upregulating the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway in L6 myotubes. These results show that SOE has the potential to be used to develop an exercise supplement capable of stimulating mitochondrial biogenesis through the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway.
Introduction
Regular exercise or physical activity is essential for maintaining a healthy lifestyle, and contributes to reducing the risk of developing metabolic diseases such as coronary heart disease, type 2 diabetes, hypertension, hyperlipidemia, and atherosclerosis as well as some cancers. 1 –3 It also increases functional exercise capacity, and hence, is needed to sustain a good quality of life. 4,5
Exercise endurance affects the metabolic characteristics of skeletal muscles by increasing the number of mitochondria, thus, enhancing oxidative metabolism and respiratory capacity. 6 –8 Exercise endurance also dominates the physiological induction of mitochondrial biogenesis in skeletal muscles. 9 Mitochondrial biogenesis is the process of mitochondria synthesis in the cell, characterized by an increase in relative mitochondrial DNA (mtDNA) content and mitochondrial function, owing to decreased oxygen consumption and increased adenosine triphosphate (ATP) production. 10
The peroxisome proliferator-activated receptor gamma co-activator 1α (PGC-1α) is a key regulator of mitochondrial biogenesis, and it controls processes involved in energy metabolism, such as muscle fiber type switching, energy substrate transport, and fatty acid oxidation in skeletal muscles. 1,9 Two major proteins, AMP-activated protein kinase (AMPK) and sirtuin 1 (SIRT1), stimulate PGC-1α through phosphorylation and deacetylation, resulting in increased mitochondrial biogenesis and improved exercise endurance. 4 Furthermore, peroxisome proliferator-activated receptor delta (PPARδ) activity is controlled by AMPK and SIRT1, which are key exercise-related markers, essential for exercise-induced adaptive responses. 8,11 Thus, exercise capacity can be regulated by activating mitochondrial biogenesis and upregulating the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway.
Siegesbeckia orientalis L. is widely distributed in the tropics and subtropics and has been used as a traditional Chinese medicine against rheumatic arthritis, hypertension, malaria, and inflammation. It has also been known to possess anti-allergic, anti-infertility, anti-inflammatory, anti-rheumatic, and immunosuppressive properties. 12 –14 However, there are very few studies on the effects of its extract on exercise capacity. S. orientalis contains bioactive compounds such as ent-kaurane, ent-pimarane diterpenoids, and sesquiterpenoids. 15 Kirenol is the main ent-pimarane diterpenoid in S. orientalis, possessing anti-adipogenic, osteoblastogenic, and anti-photoaging activities. 15 –17 In this study, we examined the stimulatory effects of the standardized S. orientalis extract (SOE) and kirenol on exercise capacity, by observing the enhancement of mitochondrial biogenesis and the activation of the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway.
Materials and Methods
Preparation of the standardized SOE
The aerial parts of S. orientalis were purchased from Radian Inc. (Chuncheon, Korea), dried, ground (100 g), and extracted with 95% ethanol for 2 days at room temperature. Then, the extract was filtered and concentrated using a rotary evaporator (Laborata 4000 Efficient; Heidolph Instruments GmbH & Co. KG., Schwabach, Germany) to obtain the SOE with a yield of 10.81% (w/w). The identification and quantification of kirenol (molecular weight: 338.48 g/mol) (Fig. 1) in the standardized SOE was performed using high-performance liquid chromatography and calculated based on the standard curve using pure kirenol (Institute for Korea Traditional Medical Industry, Daegu, Korea). The SOE contained 1.05% (w/w) kirenol as a bioactive compound.
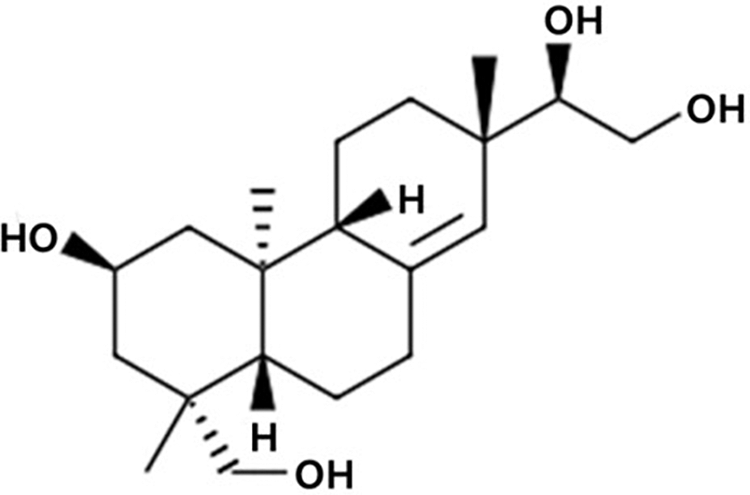
Chemical structure of kirenol.
Chemical reagents
Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), and horse serum were purchased from Hyclone Laboratories, Inc. (Logan, UT, USA). Protease inhibitor cocktail and 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) were supplied by Sigma-Aldrich (St. Louis, MO, USA). Primary antibodies against phosphorylated AMPK (p-AMPK), total AMPK (t-AMPK), SIRT1, and α-tubulin were supplied by Cell Signaling Technology (Beverly, MA, USA). Primary antibodies against PGC-1α and PPARδ were supplied by Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
Animal experiments
Four-week-old male C57BL/6J mice (Central Lab Animals, Inc., Seoul, Korea) were maintained at 25°C ± 2°C and 55% ± 5% relative humidity with a 12 h light–dark cycle and were provided food and tap water ad libitum during the course of the experiment at the Yonsei Laboratory Animal Research Center (Seoul, Korea). After acclimatizing for 1 week, the mice were separately fed either normal diet (ND; 10% kcal fat, D12450B; Research Diets, Inc., New Brunswick, NJ, USA) or high-fat diet (HFD; 60% kcal fat, D12492; Research Diets, Inc.) for 6 weeks. Then, they were randomly assigned to the following treatment groups: (1) ND; (2) ND with SOE: 250 mg/kg/day (ND+SOE); (3) HFD; and (4) HFD with SOE: 250 mg/kg/day (HFD+SOE). SOE was administered using oral gavage for 6 weeks.
Throughout the experimental period, food intake and body weight were monitored twice per week. After killing the mice, their hindlimb skeletal muscles, including soleus and gastrocnemius muscles, were separated and stored at −70°C until further use. This study adhered to the guidelines for animal experiments that were approved by the Institutional Animal Care and Use Committee (IACUC-201401-120-01) of Yonsei University.
Exercise endurance test
Before the killing, exercise performance of each mouse was tested by assessing its running capacity, as previously described. 18 A variable speed belt treadmill (Columbus Instruments, Columbus, OH, USA), enclosed in a plexiglass chamber, with a stimulus device, consisting of a shock grid attached to the rear of the belt (Panlab, Barcelona, Spain) was used to obtain measurements.
Reverse transcription–polymerase chain reaction
Total RNA from homogenized skeletal muscle tissues and L6 cells were analyzed by reverse transcription–polymerase chain reaction (RT-PCR), as previously described. 18 The sequences of primer pairs used (Bioneer, Daejeon, Korea) are listed in Table 1. PCR was carried out in a Gene Amp PCR System 2700 (Applied Biosystems, Foster City, CA, USA). Gene expression was visualized and analyzed using a G:BOX EF imaging system (Syngene, Cambridge, United Kingdom) and the Gene Snap program.
Primer Sequences Used for Reverse Transcription–Polymerase Chain Reaction Analysis
ERRα, estrogen-related receptor α; mtDNA, mitochondrial DNA; NRF-1, nuclear respiratory factor 1; PGC-1α, peroxisome proliferator-activated receptor gamma co-activator 1 alpha; TFAM, mitochondrial transcription factor A.
Western blot analysis
The molecular mechanism of the exercise-related signaling pathway was confirmed by Western blotting analysis, as previously described. 18 Primary antibodies (1:1000 dilution) against p-AMPK, t-AMPK, SIRT1, PGC-1α, PPARδ, and α-tubulin were used. Protein expression levels were visualized and analyzed using a G:BOX EF imaging system (Syngene) and the Gene Snap program.
Cell culture and differentiation
COS-7 monkey kidney cells and L6 rat skeletal muscle cells (American Type Culture Collection, Manassas, VA, USA) were maintained in DMEM with 1% antibiotics (100 U/mL penicillin A and 100 μg/mL streptomycin) and 10% FBS in a humidified atmosphere of 5% CO2 at 37°C. To stimulate differentiation of L6 cells, serum was changed to 2% horse serum. Differentiation medium containing kirenol was replaced every 2 days till day 6. Kirenol was used at various nontoxic concentrations (10, 20, and 40 μM).
Intracellular ATP content
The ATP analysis kit (Molecular Probes, Eugene, OR, USA) was used to quantify intracellular ATP in L6 cells, following the manufacturer's instructions. Luminescence intensity was detected using a MicroLumat Plus LB 96V luminometer (Berthold, Wildbad, Germany).
Luciferase reporter gene assay
PGC-1α and SIRT1 activities were assessed using the luciferase assay, by transfecting cells with the reporter plasmids, such as pGL3-PGC-1α-Luc and pTA-Luc-SIRT1 (Addgene, Cambridge, MA, USA). COS-7 cells were seeded onto a 24-well plate (4 × 106 cells/plate) and incubated for 24 h. Transfection reagent, Lipofectamine (Invitrogen, Grand Island, NY, USA) was used to co-transfect the reporter plasmids into COS-7 cells. After treatment with kirenol, transcriptional activities were determined by using the luciferase assay substrate (Promega, Madison, WI, USA) and readings were obtained with a MicroLumat Plus LB 96 V luminometer (Berthold).
Quantification of mtDNA content
Skeletal muscle mtDNA content was measured using RT-PCR, as previously described. 18 The primer pairs (Bioneer) are listed in Table 1. Results were calculated as the relative number of mitochondrial genomes per diploid nuclei. The ratio of mtDNA:genomic DNA was estimated by detecting the relative density of the expression of DNA.
Statistical analysis
Data are represented as mean ± standard deviation of three independent experiments (in vitro) or 5–7 mice per group (in vivo). Statistical analysis was conducted using SPSS version 21.0 (SPSS, Inc., Chicago, IL, USA). Differences between groups were analyzed using unpaired Student's t-tests for in vivo experiments and one-way analysis of variance (ANOVA) followed by a Scheffe's test for in vitro experiments. Results were considered statistically significant at a value of P < .05.
Results and Discussion
SOE enhances exercise endurance in mice
Before measuring the exercise endurance, we measured the body weight and food intake. The body weight gain and food intake in the SOE-treated ND group were not significantly different, as compared with the untreated ND group. In contrast, SOE treatment markedly reduced the body weight gain in HFD-induced obese mice without affecting food intake (Fig. 2).

Effects of SOE treatment on body weight and food intake in mice. Mice were orally administered ND, ND with SOE (250 mg/kg/day) (ND+SOE), HFD, and HFD with SOE (250 mg/kg/day) (HFD+SOE) for 6 weeks. Body weight and food intake were measured twice per week. All the data are expressed as mean ± SD (n = 7). ## P < .01 versus ND control mice; **P < .01 versus HFD obese mice. HFD, high-fat diet; ND, normal diet; SOE, Siegesbeckia orientalis extract.
To determinate whether SOE could increase exercise endurance, a treadmill was used to measure the running distance and time of ND and HFD mice. The running distance and time of HFD mice were markedly decreased compared with those of ND mice. However, the running distance of SOE-treated HFD and ND mice was significantly different, as they were ∼2.3 and 1.2 times longer than that of HFD and ND mice, respectively (Fig. 3).

Effect of SOE treatment on exercise endurance in mice. Mice were orally administered ND, ND with SOE (250 mg/kg/day) (ND+SOE), HFD, and HFD with SOE (250 mg/kg/day) (HFD+SOE) for 6 weeks. Mice were subjected to a treadmill test. All the data are expressed as mean ± SD (n = 5). ## P < .01 versus ND control mice; *P < .05 versus HFD obese mice. SD, standard deviation.
High fat intake causes mitochondrial dysfunction and decreases energy metabolism in the skeletal muscle. 19 Impairment of energy metabolism in the skeletal muscle may be because of mitochondrial dysfunction, which is closely related to poor exercise endurance and obesity. 20 Mitochondrial biogenesis in the skeletal muscle is closely linked to exercise endurance and energy metabolism. In this study, SOE treatment in HFD-induced obese mice increased exercise endurance and reduced body weight, which can be attributed to its ability to stimulate mitochondrial biogenesis and improve mitochondria function in the skeletal muscle. In addition, SOE treatment enhanced exercise endurance in the ND group, suggesting that SOE can stimulate mitochondrial biogenesis under normal conditions as well.
Thus, these results suggest that SOE effectively enhances exercise endurance capacity by activating skeletal muscle mitochondrial biogenesis.
SOE upregulates mitochondrial biogenesis in the skeletal muscles of mice
The beneficial effects of exercise endurance are related to the maintenance of homeostatic energy metabolism through skeletal muscle mitochondrial biogenesis and oxidative capacity. Mitochondrial biogenesis is controlled by the activation of PGC-1α. 6 PGC-1α mediates mtDNA replication and transcription through increased expression of mitochondrial transcription factor A (TFAM) because of the co-activation of nuclear respiratory factor 1 (NRF-1) and estrogen-related receptor α (ERRα), which are major regulators of mitochondrial biogenesis. 6,9
To ensure that the increase in exercise endurance in the SOE-treated mice is because of improvement of mitochondrial biogenesis, we evaluated the molecular mechanisms and mtDNA content in skeletal muscles. The mtDNA content in skeletal muscles was markedly increased in ND and HFD mice after SOE treatment (Fig. 4A). Mitochondrial biogenesis-related transcriptional factors such as NRF-1, ERRα, TFAM, and PGC-1α were upregulated in the skeletal muscles of SOE-treated ND and HFD mice (Fig. 4B).

Effect of SOE treatment on the expression of mitochondrial biogenesis markers in the skeletal muscles of mice.
The PGC-1α gene in skeletal muscle promotes mitochondrial biogenesis in response to physical performance. It indicates several hallmarks of exercise endurance-induced changes, including increases in mitochondrial function and density, and conversion of muscle fiber type toward a more oxidative form with enhanced oxidation of fat. 6,9 SOE treatment significantly upregulated the mRNA expression of PGC-1α in the skeletal muscles of ND and HFD mice, which enhanced mitochondrial biogenesis by stimulating mitochondrial regulatory genes and mtDNA content.
The stimulation of mitochondrial biogenesis-related factors can be effective in treating and delaying obesity and age-related metabolic diseases, by alleviating mitochondrial dysfunction. 21 A previous study indicated that mtDNA content, which is a biomarker for mitochondrial density, was lower in patients with metabolic syndrome than in lean controls. 22 SOE treatment inhibited HFD-induced mitochondrial dysfunction by improving mitochondrial biogenesis, which resulted in elevated exercise endurance in the HFD+SOE mice.
These results demonstrate that SOE treatment increases mtDNA content and stimulates mitochondrial biogenesis-related factors in the skeletal muscles of ND and HFD mice, suggesting its ability to mimic exercise response and enhance exercise capacity.
SOE activates the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway in the skeletal muscles of mice
AMPK phosphorylation, SIRT1 protein induction, PGC-1α deacetylation, and PPARδ protein stimulation in skeletal muscles are important participants in exercise-induced adaptive responses and mitochondrial biogenesis. 8,23 The key factors related to exercise, such as AMPK, SIRT1, PGC-1α, and PPARδ, have been observed to enhance exercise endurance without actual exercise leading to an increase in oxidative muscle fibers. 24
To investigate whether SOE could activate these proteins, we assessed the protein expression levels of AMPK, SIRT1, PGC-1α, and PPARδ in the skeletal muscles of mice. As given in Figure 5, SOE treatment enhanced the protein levels of p-AMPK, SIRT1, PGC-1α, and PPARδ in the skeletal muscles of ND and HFD mice, suggesting that SOE can activate the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway.
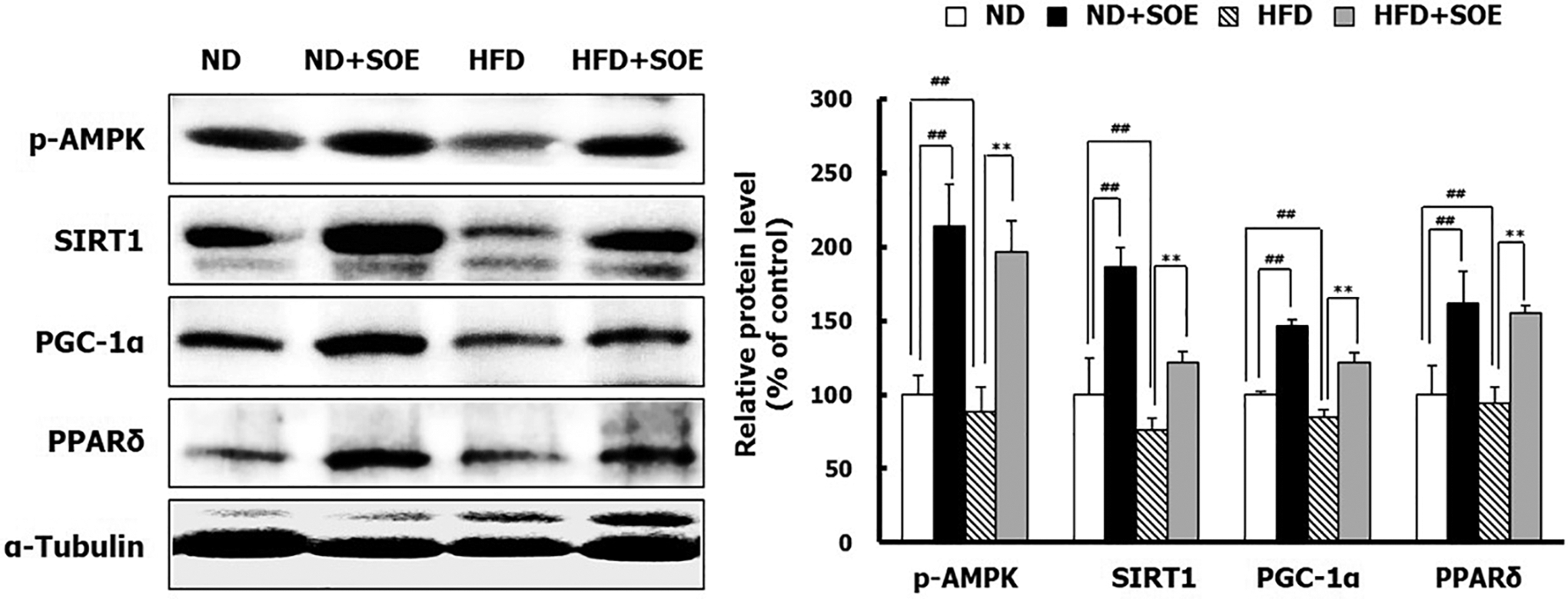
Effect of SOE treatment on the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway in skeletal muscles of mice. Total protein was isolated from skeletal muscles of mice. The protein levels of AMPK, SIRT1, PGC-1α, and PPARδ were evaluated by Western blotting. α-Tubulin was used as an internal control. All the data are expressed as mean ± SD (n = 5). ## P < .01 versus ND control mice; **P < .01 versus HFD obese mice. AMPK, AMP-activated protein kinase; SIRT1, sirtuin 1; PGC-1α, peroxisome proliferator-activated receptor gamma co-activator 1 alpha; PPARδ, peroxisome proliferator-activated receptor delta.
AMPK and SIRT1 promote the activity of important transcriptional regulators, such as PGC-1α. These factors regulate energy expenditure and mitochondrial function in response to exercise and prevent diseases related to mitochondrial dysfunction. 7,21,25 The overexpression of active PPARδ increases oxidative muscle fibers through muscle remodeling and enhances exercise performance. The interaction between AMPK and PPARδ interaction acts as a key regulator of type I fiber specification and endurance adaptations. 24,26 SOE treatment increased the activation of PGC-1α and PPARδ in the skeletal muscles of ND and HFD mice, which was mediated by AMPK and SIRT1, resulting in enhanced mitochondrial biogenesis, increased muscle fiber type switching, and upregulation of the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway. This eventually led to a boost in exercise endurance. 7,27
In addition, the SOE-treated HFD mice had upregulated expression of exercise-related markers, which are closely associated with reduction in body weight and glucose transport stimulation through the modulation of energy expenditure, lipid accumulation, thermogenesis, and metabolic homeostasis. 28,29 Mimicking exercise and activating the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway are essential for alleviating metabolic disease, and SOE activates these exercise-related signal cascades, suggesting its potential as an exercise mimetic.
Kirenol increases mitochondrial biogenesis in L6 myotubes
Exercise endurance increases mitochondrial ATP production rates that are determined by the oxidative phosphorylation in mitochondria. 22 The presence of kirenol may explain the efficacy of SOE in enhancing exercise endurance through increased mitochondrial biogenesis in skeletal muscles of mice.
To elucidate the stimulatory effect of kirenol on mitochondrial biogenesis, we measured ATP production and the underlying mechanism of mitochondrial biogenesis in L6 myotubes. Kirenol treatment elevated ATP production in a dose-dependent manner (Fig. 6A). The maximal effect was seen at 40 μM of kirenol, which increased ATP production by 76% compared with the controls. Mitochondria are crucial for the production of ATP, because they support physical activity by increasing skeletal muscle function. 30 Thus, kirenol stimulates mitochondrial activity by increasing ATP production, which may contribute to enhancing exercise endurance in the SOE-treated mice.

Effect of kirenol treatment on mitochondrial biogenesis in L6 myotubes. The cells were cultured in differentiation media with different concentrations of kirenol for 6 days.
Kirenol treatment was also effective in stimulating the mRNA expression of PGC-1α, which in turn activated mitochondrial transcription factors, such as NRF-1, ERRα, and TFAM (Fig. 6B). Thus, kirenol promotes mitochondrial biogenesis by increasing the mRNA expression of mitochondrial transcription factors that mediate mitochondrial function. These results confirm that mitochondrial biogenesis is enhanced in the skeletal muscles of SOE-treated mice.
Studies show that several natural active compounds, such as resveratrol and chitooligosaccharides, increase exercise endurance by stimulating mitochondrial biogenesis. 7,8 Likewise, kirenol can enhance exercise endurance by promoting mitochondrial biogenesis. The stimulatory effect of kirenol on mitochondrial biogenesis may contribute to the increased physical activity of SOE-treated mice.
Kirenol stimulates the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway in L6 myotubes
Exercise-related markers, such as AMPK, SIRT1, PGC-1α, and PPARδ are activated during exercise and play a critical role in exercise-induced adaptive responses. 8,11 Kirenol treatment stimulated protein expression of p-AMPK, SIRT1, PGC-1α, and PPARδ in L6 myotubes (Fig. 7A). Kirenol treatment also significantly activated SIRT1 and PGC-1α in a dose-dependent manner (Fig. 7, C). Kirenol treatment increased SIRT1 and PGC-1α activation by 50% and 79%, respectively, compared with the controls. PGC-1α activity is regulated by AMPK and SIRT1, which control energy expenditure and mitochondrial biogenesis in response to exercise. 7,25 Kirenol activated the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway, which led to improved mitochondrial biogenesis through PGC-1α activation in L6 myotubes. These results indicate that kirenol directly activates the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway by increasing SIRT1 and PGC-1α activation.

Effect of kirenol treatment on the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway in L6 myotubes. The cells were cultured in differentiation media with different concentrations of kirenol for 6 days.
To the best of our knowledge, this is the first study to report the significant effect of standardized SOE on running distance and time, by stimulating mitochondrial regulatory genes and effectively upregulating the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway in the skeletal muscles of ND and HFD mice. Furthermore, kirenol, a major bioactive constituent of SOE, increased ATP production and mitochondrial activity by upregulating the expression of biomarkers of mitochondrial biogenesis and by activating the AMPK/SIRT1/PGC-1α/PPARδ signaling pathway in L6 myotubes. These results suggest that SOE and kirenol can be potentially effective nutraceutical candidates for enhancing exercise capacity.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.