
Abstract
1′-Acetoxychavicol acetate (ACA) is found in the rhizomes or seeds of Alpinia galanga and Alpinia conchigera, which are used as traditional spices in cooking and traditional medicines in Southeast Asia. ACA possesses numerous medicinal properties. Those include anticancer, antiobesity, antiallergy, antimicrobial, antidiabetic, gastroprotective, and anti-inflammatory activities. ACA is also observed to exhibit antidementia activity. Recent studies have demonstrated that combining ACA with other substances results in synergistic anticancer effects. The structural factors that regulate the activity of ACA include (1) the acetyl group at position 1′, (2) the acetyl group at position 4, and (3) the unsaturated double bond between positions 2′ and 3′. ACA induces the activation of AMP-activated protein kinase (AMPK), which regulates the signal transduction pathways, and has an important role in the prevention of diseases, including cancer, obesity, hyperlipidemia, diabetes, and neurodegenerative disorders. Such findings suggest that AMPK has a central role in different pharmacological functions of ACA, and ACA is useful for the prevention of life-threatening diseases. However, more studies should be performed to evaluate the clinical effects of ACA and to better understand its potential.
Introduction
1′-Acetoxychavicol acetate (ACA; Fig. 1) is found in the rhizomes or seeds of Alpinia galanga and Alpinia conchigera, which are plants in the ginger family. A. galanga and A. conchigera are commonly called “greater galangal” and “lesser Alpinia,” respectively. These are used as traditional spices in cooking, especially the former is used in Thai and Indonesian cuisines, while the latter grows in eastern Bengal, southern peninsular Malaysia, and Sumatra. These rhizomes have often been used as traditional medicines for gastrointestinal disorders in Thailand and fungal infections in Malaysia.
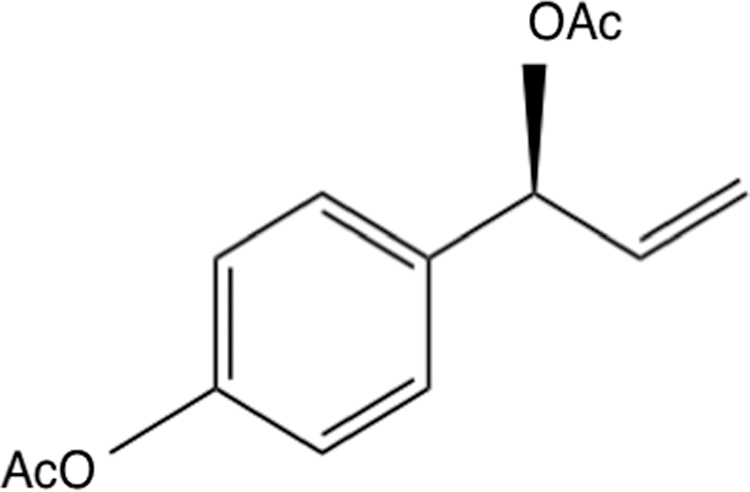
The structure of ACA. ACA, 1′-acetoxychavicol acetate.
To date, ACA, the main component of the crude ethanolic extract of rhizomes, has exhibited diverse medicinal properties like anticancer effects, gastroprotective effects, xenobiotic protection, antiallergic activity, antimicrobial activity, and antidementia effects. In this review, we aimed to summarize the diverse bioactivities of ACA, which are presented in Tables 1 and 2, the efficacy of combined treatment with ACA and other substances (Table 3), and discuss the relationships between structure and pharmacological activities of ACA and the involvement of AMP-activated protein kinase (AMPK) activation in its different pharmacological functions.
Antitumor Activity of 1′-Acetoxychavicol Acetate
ACA, 1′-acetoxychavicol acetate; GSH, glutathione; GST, glutathione-S-transferases; HUVECs, human umbilical vascular endothelial cells; miRNA, microRNA; NF-κB, nuclear factor κB; ROS, reactive oxygen species; TRAIL/Apo2L, tumor necrosis factor-related apoptosis-inducing ligand/Apo2 ligand.
Physiological Effects of 1′-Acetoxychavicol Acetate, Except Its Antitumor Activity
Il-4, interleukin-4; MIC, minimum inhibitory concentration; NQO, NAD(P)H:quinone oxidoreductase; TNF-α, tumor necrosis factor-α; TRPA1, transient receptor potential cation channel subfamily V, member 1.
Effects of Combination Treatment with 1′-Acetoxychavicol Acetate and Other Substances
AMPK, AMP-activated protein kinase; NRF2, nuclear factor, erythroid 2 (NF-E2)-related factor-2.
Pharmacological Effects of ACA
Anticancer effects of ACA
Cancer is the second leading cause of death in the world. Therefore, numerous efforts have been dedicated to establishing preventive methods and discovering new treatments.
ACA is one such preventive method and has been reported to exhibit potent anticancer effects against various tumor cells, such as colorectal adenocarcinoma cells, cervical cancer cells, oral squamous carcinoma cells, prostate cancer cells, breast cancer cells, glioblastoma cells, myeloma cells, hepatocellular carcinoma cells, and Ehrlich ascites tumor cells (EATCs).
After examining the antitumor activity of ACA with a convenient short-term in vitro assay that involved tumor promoter-induced Epstein-Barr virus (EBV) activation in 1993, Kondo et al. became the first to report that ACA has anticancer effects. Based on their results, the rhizome extract and its main component, ACA, potently inhibited EBV activation induced by a tumor promoter, 12-O-tetradecanoylphorbol-13-acetate. 1,2 Thereafter, many researchers identified the influences of ACA in the initiation step of carcinogenesis via in vivo experiments. Among them, Ohnishi et al. found that ACA ingestion during the initiation phase effectively inhibited 4-nitroquinoline 1-oxide-induced oral carcinogenesis in male F344 rats. 3 ACA also exhibited preventive effects on chemical carcinogenesis in the skin. Murakami et al. also examined the effect of ACA in a two-stage carcinogenesis experiment in ICR mouse skin and found that topical application of ACA markedly reduced the average number of tumors. 4 Furthermore, Tanaka et al. found that dietary ACA effectively suppressed the development of colonic crypt foci induced by azoxymethane in rats when ingested during or after the initiation phase. 5 Kobayashi et al. demonstrated that ACA prevents endogenous rat liver carcinogenesis in male Fisher 344 rats by preventing oxidative DNA damage. 6 Feeding ACA in diet at the initiation step inhibited the cholangiocarcinogenesis induced by N-nitrosobis(2-oxoprooyl)amine in hamsters 7 and esophageal tumorigenesis induced by N-nitrosomethylbenzylamine in male F344 rats. 8
In 2001, Mori et al. found that ACA causes apoptosis and suppresses cell proliferation.
9
Many reports exist on the mechanism of apoptosis induced by ACA. In fact, Ito et al. indicated that ACA induced apoptosis, and was independently mediated via the mitochondrial oxidative stress pathway and Fas-induced apoptotic pathway in myeloid leukemia.
10
They also reported that ACA suppresses the proliferation of human myeloma cells by inhibiting the translocation of nuclear factor κB (NF-κB) to nuclei, and then induces the apoptosis of myeloma cells.
11
Ichikawa et al. reported that ACA enhances apoptosis and inhibits invasion by blocking NF-κB activation using different agents and numerous human cell lines.
12
Batra et al. indicated that ACA inhibits skin tumorigenesis in constitutive Stat3 expressed transgenic mice (
The efficacy of the antitumor activity of ACA, which is reported as a suppressor of carcinogenesis, on polyamine (putrescine, spermidine and spermine) metabolism was examined in EATCs. Polyamines are necessary for the proliferation and differentiation of cells. In our laboratory, Moffatt et al. found that the antitumor effects of ACA were derived from the perturbation of the late limiting enzymes in the polyamine metabolic pathway and the enhancement of caspase-3-like activity. 16 They also found that ACA decreased in phosphorylated retinoblastoma protein, and induced nuclear localization of p27kip1, followed the accumulation of G1 phase of EATC. 17,18 Furthermore, these events were demonstrated to be cellular thiol-dependent. 19 –21 Baradwaj et al. confirmed that the mode of Dukes' type B colorectal adenocarcinoma (SW480) cell death with ACA was due to apoptosis and the G0/G1 arrest. 22 In addition, they observed an increase in p21 expression and a concomitant decrease in cyclin D in SW480 cells treated with ACA.
ACA caused the suppression of angiogenesis-mediated prostate tumor growth. Angiogenesis plays an important role in cancer metastasis. By examining the effect of ACA on antiangiogenesis activity, Pang et al. found that ACA inhibited the vascular endothelial growth factor-induced migration, proliferation, adhesion, and tubulogenesis of human umbilical vascular endothelial cells. 23
Recently, several researchers have published intriguing findings that ACA suppresses the microRNA (miRNA) expression in several cancer cells. miRNAs are highly conserved small noncoding RNAs that post-transcriptionally regulate gene expression.
miRNAs bind to target sequences in the noncoding regions of target messenger RNAs and induce messenger RNA degradation or suppress translation. 24 An miRNA is able to regulate many genes; a gene can be targeted by different miRNAs; and each miRNA can regulate human tumorigenesis, cell proliferation, metabolism, or apoptosis. 25 Phuah et al. examined the combined effects of ACA and cisplatin, a platinum-based anticancer drug, on HPV-positive human cervical carcinoma cell lines, and identified the miRNAs regulated in response to ACA and/or cisplatin. They reported that ACA synergistically enhances the cytotoxic effects of cisplatin by dysregulating specific miRNAs in cervical carcinoma cells. These researchers also emphasized that alterations in miRNA expression affect many important signaling pathways, such as the ERK pathway. 26 Wang et al. demonstrated that ACA inhibits the proliferation of HN4, a human head and neck squamous cell carcinoma cell (SSC) line, and downregulates miRNA-23a expression. They confirmed that phosphatase and tensin homologs deleted on chromosome 10 (PTEN) are the targets of miRNA-23a 27 and proceeded to demonstrate that the downregulation of miRNA-210 conferred sensitivity toward ACA in cervical cancer cells by binding to sequences in the 3′ noncoding region of the Smad4 mRNA. 28 Recently, Phuah et al. reported that sensitivity to ACA was increased by the suppression of miRNA-629 with reducing cell growth and inducing apoptosis in cervical cancer cells. 29 These results suggest that combining miRNAs and natural compounds like ACA, could become new strategies to treat cervical cancer. These and other reports on anticancer effects are shown in Table 1.
Antiobesity effects of ACA
Obesity is one of the main risk factors for type 2 diabetes, cardiovascular disease, and chronic kidney disease. Several drugs that decrease appetite, fat absorption, and fat oxidation have been developed to treat obesity. However, many have disappeared or been removed from the market due to low efficacy and side effects. Natural compounds cause minimal side effects, consequently they are excellent sources for developing new drugs. Therefore, use of natural compounds may be a promising antiobesity strategy. 30
Narukawa et al. found that ACA acts as an agonist to activate transient receptor potential cation channel subfamily V, member 1 (TRPA1). 31 Furthermore, cinnamaldehyde, the TRAP1 agonist, is involved in the reduction of visceral body fat and exhibits antiobesity effects. 32 These results indicate that ACA may have antiobesity effects.
By investigating the antiobesity effect of ACA, Ohnishi et al. found that ACA inhibits cellular lipid accumulation by downregulating transcription factors, such as PPARγ and C/EBPα in 3T3-L1. ACA also caused the activation of AMPK. In an in vivo animal obesity model, rats fed a high fat diet (HFD) with 0.05% ACA gained significantly less body weight than rats fed an HFD alone. Additionally, the visceral fat mass in rats fed an HFD with 0.05% ACA was lower than that in rats fed HFD alone. 33 Recently, many substances with antiobesity effects that are regulated by the activation of the AMPK signaling pathway have been reported. 34 –36 These findings suggest that AMPK mediates the antiobesity effect of ACA.
Gastroprotective effects of ACA
In 1976, Mitsui et al. found that ACA had strong inhibitory effect against Shay ulcer in rats 37 ; Shay ulcer was established through pylorus ligation according to Shay's method.
Owing to the absence of studies on the gastroprotective effect of ACA using other ulcer models, Matsuda et al. opted to examine its effect on other ulcer models. Based on their results, ACA significantly protected against ethanol-induced gastric mucosal lesions. 38 Furthermore, ACA prevented 0.6 M HCl-and aspirin-induced gastric mucosal lesions. However, the gastroprotection of ACA was inhibited by pretreatment with indomethacin and N-ethylmaleimide, an SH-blocker. Such findings indicate that the protective effect of ACA may be associated with endogenous prostaglandins and intracellular glutathione (GSH) levels. By performing a further examination of the structure-activity relationship of ACA to elucidate its gastroprotective effect using various related compounds these researchers also reported that the 1′-acetoxyl group of ACA is essential for this effect. 38
Free radicals by oxygen and lipid peroxidation are considered to be factors that cause gastrointestinal lesions. Therefore, these results indicate that the GSH levels increased by ACA in gastric mucosa may prevent the increase in free radicals and lipid peroxidation and then prevent lesions caused by different ulcers.
Xenobiotic protective activities of ACA
Xenobiotic metabolism that alters the structure of exogenous or endogenous chemicals is an important metabolism for maintaining homeostasis and cell conservation. The xenobiotic metabolism pathway consists of three steps: phase 1, phase 2, and phase 3. In phase 1, xenobiotics are modified by hydrolysis, oxidation, and reduction and ultimately become the active metabolites. In phase 2, the phase 1 product is converted to hydrophilic products by transferase enzymes. In phase 3, the final modified product is excreted from the cells via different transporters. The detoxification process is catalyzed by the cytochrome P450 (CYPs) family, UDP-glucuronosyltransferases (UGT) family, glutathione-S-transferases (GST) family, and aldehyde dehydrogenase family. In particular, UGT and GST families in phase 2 play important roles to determine the rate of xenobiotic metabolism.
Phytochemicals, such as flavonoids, sulforaphane, and curcumin, induce the phase 2 enzyme activities. When examining the effect of ACA on colon tumorigenesis induced by azoxymethan, Tanaka et al. found that dietary ACA inhibited tumorigenesis by suppressing cell growth in the colonic mucosa and inducing phase 2 detoxification enzymes, GST and NAD(P)H:quinone oxidoreductase (NQO), in the liver and colon. 5
In our research group, Yaku et al. revealed that ACA induces GST and NQO, in normal cells and rat intestine epithelial cells (IEC-6). They also found that the induction of phase 2 enzymes with ACA was caused by the upregulated levels of intracellular nuclear factor, erythroid 2-related factor-2, and cytosolic p21. 39
Antiallergy effects of ACA
Allergies are hypersensitivity disorders of the immune systems. Matsuda et al. found that ACA could suppress the secretion of β-hexosaminidase, a marker of antigen- immunoglobin E-mediated degranulation, in rat basophilic leukemia cells. 40 β-Hexosaminidase accumulates in the secretory granules of mast cells and is concomitantly released with histamine when mast cells are activated immunologically. 41 Matsuda et al. also reported that ACA inhibits the release of tumor necrosis factor-α and interleukin-4, which are involved in the late phase of type I allergic reaction. 40 Furthermore, ACA suppressed ear passive cutaneous anaphylactic reactions in mice.
Asthma is an immune-mediated disease that is associated with an imbalance between T helper subsets. This imbalance causes the upregulation of cytokines and then promotes chronic inflammation in the respiratory system.
By examining the effect of ACA in an ovalbumin-induced asthma mouse model, Seo et al. found that ACA dose-dependently reduced white blood cell infiltration in the lungs of the model mouse. 42 This increase in white blood cell infiltration is characteristic of asthma.
Antimicrobial effect of ACA
The antimicrobial activity of A. galanga extract was previously reported. 30 In Thailand, Phongpaichit et al. extracted many medicinal plants with chloroform, methanol, and water and evaluated their antimycobacterial activity against Mycobacterium tuberculosis H37 Ra. 43 As a result, they found that the chloroform extract of A. galanga was most active. The minimum inhibitory concentration (MIC) was 0.12 μg/mL. This MIC was almost the same as that of isoniazid, an antituberculosis drug (0.1 μg/mL). Furthermore, after isolating ACA from a chloroform extract of A. galanga and evaluating its antimycobacterial activity, they found that ACA has very strong antimycobacterial activity with an MIC of 0.024 μg/mL.
Recently, Warit et al. examined the effects of the S-enantiomer of ACA (S-ACA), isolated from A. galanga, and the synthetic racemic 1′-R, S-ACA (rac-ACA) on the cytocidal activity of M. tuberculosis H37Ra and H37Rv. Based on their findings, S-ACA and rac-ACA had MICs of 0.5 and 2.7 μg/mL, respectively. 44 These findings suggest that S-ACA exhibits potent bactericidal activity.
Latha et al. reported that ACA has an antiplasmid activity against multidrug-resistant bacteria. 45 Furthermore, ACA showed antibacterial activity against the anaerobic bacterium, Propionibacterium acnes. 46
Effect of ACA on proteasome activity
The ubiquitin-proteasome-system, one of the protein degradation systems, is an important pathway for the degradation of misfolded, old-, and short-lived proteins. However, proteasome activity decreases in an age-dependent manner and causes age-related neurodegenerative processes.
Based on the relationship between proteasome activity and ACA in the neurodegenerative process, Yaku et al. examined the effect of ACA on proteasome activity and cell protection against neurotoxicity using differentiated PC12 cells. Based on their findings, ACA enhanced proteasome activity during the early stages of ACA treatment in PC12 cells. 47 Furthermore, they reported that the increased proteasome activity was inhibited by H-89, a cAMP-dependent protein kinase inhibitor. ACA also protected against amyloid β-protein fragment induced-neurotoxicity in differentiated PC12 cells. On the other hand, the treatment with MG132, a proteasome inhibitor, reduced the protective effect of ACA. Altogether, these findings demonstrate that ACA induces protease activity by activating the cAMP/PKA signaling pathway in neuronally differentiated PC12 cells.
Antidementia effect of ACA
Alzheimer's disease (AD) is an irreversible progressive neurodegenerative disease with age-dependent and impaired cognitive function and thinking ability. In our laboratory, the effect of ACA on learning and memory with ageing was examined using senescence-accelerated mice prone 8 (SAMP8), a suitable AD model. 48 The learning and memory abilities of SAMP8 fed the 0.02% ACA diet using the Morris water maze test were significantly improved compared to those of SAMP8 mice fed the control diet.
We also examined short-term memory by observing the spontaneous alternations of mice using the Y-maze test. The spontaneous alterations in SAMP8-control mice were significantly decreased compared to the senescence-accelerated resistant/1 (SAMR1) mice. On the other hand, the cerebral tissue in SAMR1 mice was observed to be normal. However, in SAMP8-control mice, neuronal damage in the hippocampal CA1 region was observed. Nonetheless, the hippocampal CA1 region in SAMP8-ACA mice were almost normal, similar to that of SAMR1. The study also showed that ACA increased the serum concentration of β-hydroxybutyric acid, and suggested that this may be involved in the amelioration of cognitive function in SAMP8-ACA mice.
β-Hydroxybutyric acid is presumed to assume roles such as an alternative energy to glucose and an inhibitor of histone deacetylase; however, its uptake into neurons is reduced by aging or an epigenetic action. 49 Further research is thus needed to draw definitive conclusions.
These and other reports on the physiological effects of ACA, except its antitumor activity, are summarized in Table 2.
Effects of Combination Treatment with ACA and Other Substances
Many studies have reported that combination treatments with natural products and other substances, such as chemotherapy agents, may be more efficacious than higher or maximal doses of a single agent. 50 Recently, the effects of combination treatment with ACA and other substances were examined in several experimental systems.
In et al. examined the effect of a combination of ACA and cisplatin in oral SSC and in vivo human oral tumor xenografts using Nu/Nu mice. 51 In SSC, combined treatment with ACA and cisplatin was found to enhance the cytotoxic effects in a synergistic manner. Furthermore, ACA was found to potentiate the effect of cisplatin and thereby caused a reduction in tumor volume.
Phuah et al. also investigated the effect of ACA combined with cisplatin on HPV-positive human cervical carcinoma cells. 26 They found that combined treatment induced cytotoxity dose- and time-dependently and enhanced the expression of 25 miRNAs, including has-miR-138, has-miR-210, and has-miR-744, with predicted gene targets involved in the signaling pathways that regulate apoptosis and cell cycle progression. Such findings suggest that miRNAs have an important role in the response of anticancer drug to ACA, and synergistically potentiate the effect of cisplatin as a chemosensitizer.
Previously, we investigated the effect of the combination of ACA and sodium butyrate. Moreover, Kato et al. evaluated the combined effect of ACA and sodium butyrate on the proliferation of HepG2 human hepatocellular carcinoma cells in our laboratory. 52 In combination treated cells, cell numbers were synergistically decreased via apoptosis; however, this decrease was suppressed in the cells pretreated with catalase, which also caused a significant increase in NADPH oxidase activities and intracellular reactive oxygen species (ROS) levels. Such findings suggest that an increase in intracellular ROS levels relates with cancer cell death. AMPK phosphorylation was also significantly induced in cells treated with the combination; however, this induction was suppressed in the cells pretreated with catalase, suggesting that an increase in ROS is involved in the enhancement of phosphorylation of AMPK. These results indicate that combination with ACA and sodium butyrate synergistically induces apoptosis by increasing intracellular ROS and AMPK phosphorylation.
Yaku et al. also examined the effect of the combination of ACA and sodium butyrate on phase 2 enzyme activities in intestinal epithelial cells in our laboratory. 53 ACA and sodium butyrate synergistically increased phase 2 enzyme activities and promoted p53 acetylation. Furthermore, the inhibition of AMPK activity was found to reduce the phase 2 enzymes. Such findings suggest that the combined treatment of ACA and sodium butyrate synergistically upregulates phase 2 enzyme activities through activation of AMPK and acetylation of p53.
Arshad et al. examined the effect of ACA and recombinant human alpha fetoprotein (rhAFP) on human cancer xenografts in athymic nude (Nu/Nu) mice. 54 According to their findings, mice administered the combined treatment of rhAFP and ACA had a synergistic reduction in tumor volume compared to those administered each agent alone. The combined treatment could not only inhibit the translocation of NF-κB to the nuclei but also decreased the expression of NF-κB regulated genes that code for cyclooxygenase-2, 5-lipoxygenase, histone deacetylase, and vascular endothelial growth factor.
These and other reports on the effects of combined treatments of ACA and other substances are summarized in Table 3.
These findings may provide new insights for the use of novel combination therapies against diseases, especially carcinogenesis.
Structure-Activity Relationships of ACA
The relationships between structure and activity of ACA have been investigated in several experimental systems. Murakami et al. investigated the structure-activity relationship of ACA in a test for inhibition of tumor promoter teleocidin B-4-induced EBV activation in Raji cells by using 16 derivatives of ACA. As a result, they demonstrated that the structural factors that regulate its activity were (1) the two acetyl groups at positions 1′ and 4. They also confirmed that (2) the unsaturated double bond between positions 2′ and 3′ of ACA are essential for ACA activity; and (3) the configuration at position 1′ of ACA does not affect its activity. 55 Matsuda et al. examined the relationships in an inhibitory test to elucidate the secretion of β-hexosaminidase, a marker of antigen-IgE-mediated degranulation in RBL-2H3 cells, 40 and its inhibitory effect on nitric oxide (NO) production in lipopolysaccharide-activated mouse peritoneal macrophages. 56 Based on the effects of different compounds in the β-hexosaminidase release pathway, the 1′- and 4-acetoxyl groups of ACA and the 2′–3′ double bond enhanced its activity.
For the inhibitory effects of ACA on NO production, Matsuda et al. clarified the following structure-activity relationships: (1) the para or ortho substitution of the acetoxyl and 1-acetoxypropenyl groups at the benzene ring is essential; (2) the S configuration of the 1′-acetoxyl group is preferable; (3) the presence of the 3-methoxyl group and the loss of the 2′–3′ double bond by hydrogeneration reduce ACA activity; (4) the substitution of the acetyl groups with propionyl or methyl groups reduces ACA activity; and (5) the lengthening of the carbon chain between the positions 1′ and 2′ reduces ACA activity.
Azuma et al. studied the structure-activity relationship to elucidate the apoptotic activity of ACA in HL-60 cells human leukemia. As a result, they demonstrated that (1) the two acetyl groups at positions 1′ and 4′ of ACA and (2) the unsaturated double bond between positions 2′ and 3′ of ACA are essential for ACA activity, but (3) the configuration at position 1′ of ACA does not affect its activity. 57
Liu et al. examined the structure-activity relationships of ACA using fission yeast expressing nuclear export signal of Rev, an HIV-1 viral regulatory protein. Based on their findings, (1) para substitution of the acetoxyl and 1′-acetoxypropenyl groups at the benzene ring is essential, (2) linear ethyl and propyl chain carbonates are more active than branching chain carbonates, and (3) substitution of the acetoxyl groups with alkylcarbomate groups prevents or reduces ACA activation. 58
These results demonstrate that the structural factors regulating the activity of ACA are: (1) the acetyl group at position 1′; (2) the acetyl group at position 4; and (3) the unsaturated double bond between positions 2′ and 3′ (Fig. 2).

Structural factors regulating the activity of ACA. Color images are available online.
Summary
ACA is found in A. galanga and A. conchigera, which are often used as traditional spices in cooking, especially in Thailand and Indonesia.
A quarter-century ago, Kondo et al. discovered the anticancer effects of ACA when they inhibited tumor promoter-induced EBV activation, which was employed as an in vitro assay system. Since then, many researchers have investigated the anticancer effects of ACA in many experimental systems. Recent studies have shown that the combination with ACA and other substances exhibits synergistical effects on anticancer activities. These findings may give new insights for using novel combination therapies against cancer.
Many in vitro and in vivo researches have shown that ACA has several biological and pharmacological effects, including antimicrobial, antiallergy, anti-inflammatory, antiobesity, and antiasthmatic activities and gastroprotective effects. Recently, we demonstrated that ACA has protective effects against dementia. 48 Based on prior findings, these pharmacological effects of ACA might be mediated by the activation of AMPK.
Altogether, these findings indicate that ACA may be beneficial for preventing lifestyle-related diseases. However, more studies should be carried out to examine the clinical effect of ACA to obtain a better understanding of its potential.
Footnotes
Author Disclosure Statement
No competing financial interest exists.
Funding Information
This work was supported by JSPS KAKENHI (Grant Nos. JP 24500987 and JP15K00832).