
Abstract
The aim of this study was to investigate the potential protective effects of the hot water extract of Eriobotrya japonica (EJW) on EtOH- or free fatty acid (FFA)-induced fatty liver injury in vitro. HepG2/2E1 cells were exposed to EtOH and HepG2 cells were exposed to a mixture of FFAs (oleic acid:palmitic acid, 2:1) to stimulate oxidative stress and to induce lipid accumulation, respectively. Antioxidant activity was significantly increased and lipid accumulation was inhibited in cells pretreated with EJW compared to those in cells exposed to EtOH or FFA only. Also, 5′adenosine monophosphate (AMP)-activated protein kinase (AMPK) and acetyl-coenzyme A carboxylase (ACC) phosphorylations were considerably increased, indicating activation of AMPK. Furthermore, EJW reduced the messenger RNA (mRNA) expression of lipogenesis-associated factors such as ACC, sterol regulatory element binding protein-1c (SREBP-1c), and fatty acid synthase (FAS), and increased mRNA expression related to components of the fatty acid β-oxidation pathway, such as AMPK, carnitine palmitoyltransferase 1 (CPT-1), and peroxisome proliferator-activated receptor alpha (PPARα). These results suggest that EJW possessed potential preventive effects against both EtOH- and FFA-induced fatty liver disease by alleviation of oxidative stress and lipid accumulation in hepatocytes.
Introduction
Fatty liver disease (FLD), the early stage of chronic liver disease, is characterized by excess triglyceride (TG) accumulation within the liver, representing 5–10% of the organ's weight. There are two types of FLD: alcoholic FLD (AFLD), which is caused by excessive alcohol intake, and nonalcoholic FLD (NAFLD). 1,2 Studies have reported that AFLD occurs in individuals who consume >40–60 g/day pure alcohol and that the amount and duration of alcohol consumption are more relevant than the type of alcohol. 3,4 NAFLD is distinguished from AFLD in that it occurs in individuals who consume <20 g/day pure alcohol. 5,6 NAFLD appears to be closely associated with obesity, type 2 diabetes, hyperlipidemia, and metabolic syndrome. The dietary pattern and lifestyle are important pathogenic factors of these metabolic diseases. 7 NAFLD occurs in individuals with excessive caloric intake and reduced caloric combustion, which increase the levels of circulating free fatty acids (FFAs). 8 These overloading FFAs are transported to the liver where they accumulate as TGs. 9 If this phenomenon is not prevented in the early stages, simple fatty liver can progress to steatohepatitis, fibrosis, cirrhosis, and even hepatocellular carcinoma, thereby increasing the mortality rate. 10
In one way, alcohol is metabolized by the cytochrome P450 2E1 (CYP2E1) pathways in hepatocytes and an excess of alcohol promotes CYP2E1 activity. 11 A published study suggests that the upregulation of CYP2E1 activity and the elevation of circulating FFA increase reactive oxygen species (ROS) production. 12 Increased ROS enhances oxidative stress and lipid peroxidation, which may lead to the formation of secondary products such as malondialdehyde (MDA). 13 Its accumulation in hepatocytes may further promote oxidative stress during NAFLD development. Furthermore, excessive oxidative stress activates de novo lipogenesis factors, including sterol regulatory element binding protein-1c (SREBP-1c). There are some studies demonstrating that SREBP-1c is significantly activated by H2O2 and ethanol treatment in both HepG2 cells and C57BL/6J mice. 14,15 Furthermore, ROS stimulation upregulates other lipogenesis factors, such as fatty acid synthase (FAS) and acetyl-coenzyme A carboxylase (ACC), thus inducing lipid accumulation in hepatocytes. 16 Excessive oxidative stress promotes an imbalance between FFA synthesis and oxidation. In several studies, the inhibition of peroxisome proliferator-activated receptors (PPARs) and carnitine palmitoyltransferase 1 (CPT-1), key factors of β-oxidation, was observed in fatty liver models. 17,18 Hepatic antioxidants are required to prevent excessive oxidative stress. Natural materials derived from plants possess various secondary metabolites that display antioxidant activities and are traditionally used for the prevention of various diseases. 19
Eriobotrya japonica, also known as Loquat, is a subtropical evergreen shrub that contains large amounts of phenolic compounds. It exerts well-described antioxidant effects in vitro and in vivo. 20 Several studies have suggested that loquat extract increases antioxidant enzyme activities, exerts hepatoprotective effects against ethanol-induced liver disease, decreases lipid peroxidation, and suppresses nonalcoholic steatohepatitis-associated fatty liver. 21,22
In this study, the potential duel effects of loquat-water extract (hot water extract of E. japonica [EJW]) against AFLD and NAFLD were investigated, using a cell model. Also, its underlying mechanism relevant to lipid accumulation in hepatocytes was assessed by examining the expression of hepatic lipid metabolic genes, and proteins were determined.
Materials and Methods
Sample and chemicals
Loquat leaves harvested in Korea were purchased from Hwasun traditional medicine market. Minimum essential medium (MEM), fetal bovine serum (FBS), antibiotics, trypsin-EDTA, and Hanks balanced salt solution (HBSS) were from Gibco BRL (Grand Island, NY, USA). XTT sodium salt, 4-nitrocatechol, 2′,7′-dichlorodihydrofluorescein diacetate, and Oil Red O were from Sigma-Aldrich (St. Louis, MO, USA). All other chemicals used were of the highest purity from commercial supplies.
Cell culture
HepG2 cells were obtained from the American Type Culture Collection (Manassas, VA, USA) and grown in MEM containing 10% FBS and 1% penicillin/streptomycin in a humidified incubator with 5% CO2 at 37°C. To produce CYP2E1-overexpressing HepG2 cells (HepG2/2E1), a vector containing human CYP2E1 complementary DNA (cDNA) was transfected into the cells using Lipofectamine 2000™ transfection reagent (Invitrogen, Carlsbad, CA, USA).
Extraction
To prepare EJW, loquat leaves (50 g) were extracted with 20 volumes of hot water at 100°C for 3 h. The extracted solution was filtered and concentrated using a vacuum evaporator. The extract was stored at −20°C until further use.
EtOH-induced and FFA-induced lipid overloading
In the AFLD cell model, HepG2/2E1 cells (5 × 104 cells/well) seeded in a 24-well plate were pretreated with EJW for 30 min and 200 mM ethanol was added into each well according to general protocols. 23,24 The medium was exchanged for fresh medium containing ethanol every day for 3 days, maintaining minimal evaporation. 25 In the NAFLD cell model, HepG2 cells (1 × 105 cells/well) seeded in a 24-well plate were pretreated with EJW solution for 2 h, and then, 1 mM FFA mixture was added into each well. To prepare the FFA mixture, sodium oleate (Sigma-Aldrich) and sodium palmitate (Sigma-Aldrich) were conjugated with culture medium containing 1% FFA-free bovine serum albumin (BSA; Sigma-Aldrich). The FFA mixture (2:1 ratio of oleate:palmitate) was diluted in culture medium to obtain the desired final concentration.
Assay for hepatoprotection against ethanol-induced cytotoxicity
For the assay of hepatoprotective activity in EtOH-induced model, cytotoxicity of cells was measured according to the XTT assay. Each well was treated with the freshly prepared XTT-PMS solution (composed of 1 mg XTT and 10 mg PMS/mL of phenol red-free MEM). After incubation for 2 h, the cell viability was measured with a spectrophotometer at a wavelength of 450 nm.
Determination of intracellular ROS formation
Measurements of generated intracellular ROS were assayed by a fluorescence assay. After treatment, the cells were exposed to 30 μM DCF-DA for 30 min at 37°C. The intracellular ROS generation was detected with an excitation wavelength of 485 nm and an emission wavelength of 530 nm.
Determination of lipid accumulation (Oil Red O staining)
After treatment, the cells were fixed with 10% formalin for 30 min and stained with Oil Red O solution for 40 min. The stained cells were washed to remove the remnants of the Oil Red O reagent and observed under a microscope (Leica DMi1; Leica Microsystems, Wetzlar, Germany). The lipid droplets were extracted using 60% isopropanol and assessed colorimetrically at 510 nm.
Antioxidant enzyme activity
Cells treated with EJW solution and ethanol or FFA mixture were washed twice with 1 × phosphate-buffered saline (PBS) and lysed using RIPA lysis and extraction buffer (Thermo Fisher Scientific, Waltham, MA, USA) containing mixed protease inhibitor cocktails. Superoxide dismutase (SOD), catalase (CAT), glutathione (GSH), glutathione S-transferase (GST), GSH peroxidase (GPx), and GSH reductase (GR) activities were measured according to published methods. 26 –28 To assess lipid peroxidation, MDA concentrations were measured using thiobarbituric acid (TBA) following the well-established method. 29
Total RNA isolation and real-time polymerase chain reaction
Total RNA was extracted using an easy-BLUE™ total RNA extraction kit (Intron Biotechnology, Sungnam, Korea). After isolation of total RNA, cDNA was synthesized using an iScript™ cDNA Synthesis Kit (Bio-Rad Laboratories, Hercules, CA, USA). Custom-designed primers (Table 1) were used for performing the real-time polymerase chain reaction (PCR) assay with the CFX96 Touch™ real-time PCR detection system (Bio-Rad Laboratories). PCR program used were as follows: 40 cycles of denaturation (95°C for 30 sec), annealing (58°C for 30 sec), and extension (72°C for 45 sec).
Real-Time Polymerase Chain Reaction Primer Sequences
ACC, acetyl-coenzyme A carboxylase; AMPK, 5′adenosine monophosphate (AMP)-activated protein kinase; CD36, cluster of differentiation 36; CPT-1, carnitine palmitoyltransferase 1; CYP2E1, cytochrome P450 2E1; FATP2, fatty acid transport protein 2; PPAR, peroxisome proliferator-activated receptor; SREBP-1c, sterol regulatory element binding protein-1c.
Western blotting
Equal amounts of proteins (80 μg/lane) in cell lysates were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to PVDF membranes (Bio-Rad Laboratories). Using blocking buffer (10% blocker BSA in PBS; Thermo Fisher Scientific), the membranes were blocked for 30 min. The blotted membranes were washed with PBS with Tween 20 and the membranes incubated with the first antibody purchased from Cell Signaling Technology (Danvers, MA, USA) at 4°C overnight. Thereafter, the membranes were incubated with a secondary antibody (anti-rabbit immunoglobulin G [IgG]) for 2 h. All protein bands were developed using chemiluminescence (ECL) Western blotting detection reagents (Bio-Rad Laboratories).
Statistical analysis
Data are presented as mean ± standard deviation. The data were statistically evaluated by one-way analysis of variance using SPSS statistical procedures for Windows (SPSS PASW Statistics 22.0; SPSS, Inc., Chicago, IL, USA), and Duncan's multiple range test was used to compare significant differences between groups (P < .05).
Results
Hepatoprotective activity of EJW in EtOH-treated cells
We investigated the hepatoprotective effect of EJW against EtOH-induced toxicity. The viability of EtOH-treated cells was remarkably lower compared with control cells. However, when the cells were pretreated with EJW (50 and 100 μg/mL, the concentrations selected to minimize cytotoxicity, data not shown), cell viability increased compared to that in the EtOH-treated group (P < .05, Fig. 1). Also, the hepatoprotective activity was increased in a dose-dependent model.

Hepatoprotective activity of the EJW in EtOH-treated HepG2/2E1 cells. CON, normal control group; EtOH, EtOH-treated group; EJW50 and EJW100, pretreatment with EJW 50 and 100 μg/mL groups, respectively. Data are expressed as mean ± SD (n = 3), and different letters indicate significant differences (P < .05, a > b), as determined by Duncan's multiple range test. EJW, hot water extract of Eriobotrya japonica; SD, standard deviation.
Effect of EJW on lipid accumulation on EtOH-treated and FFA-treated cells
Lipid accumulation in the EtOH- and FFA-treated groups was significantly higher than that in the control group. Furthermore, we found that cells in the EtOH-treated and FFA-treated groups showed deeper red color of Oil Red O staining than those in other groups (Fig. 2), indicating the presence of lipid droplets in the cells. However, lipid accumulation and droplet formation were significantly lower in the EJW-treated groups than in the EtOH-treated and FFA-treated groups (P < .05). In both models, the lipid accumulation was dose dependently decreased.
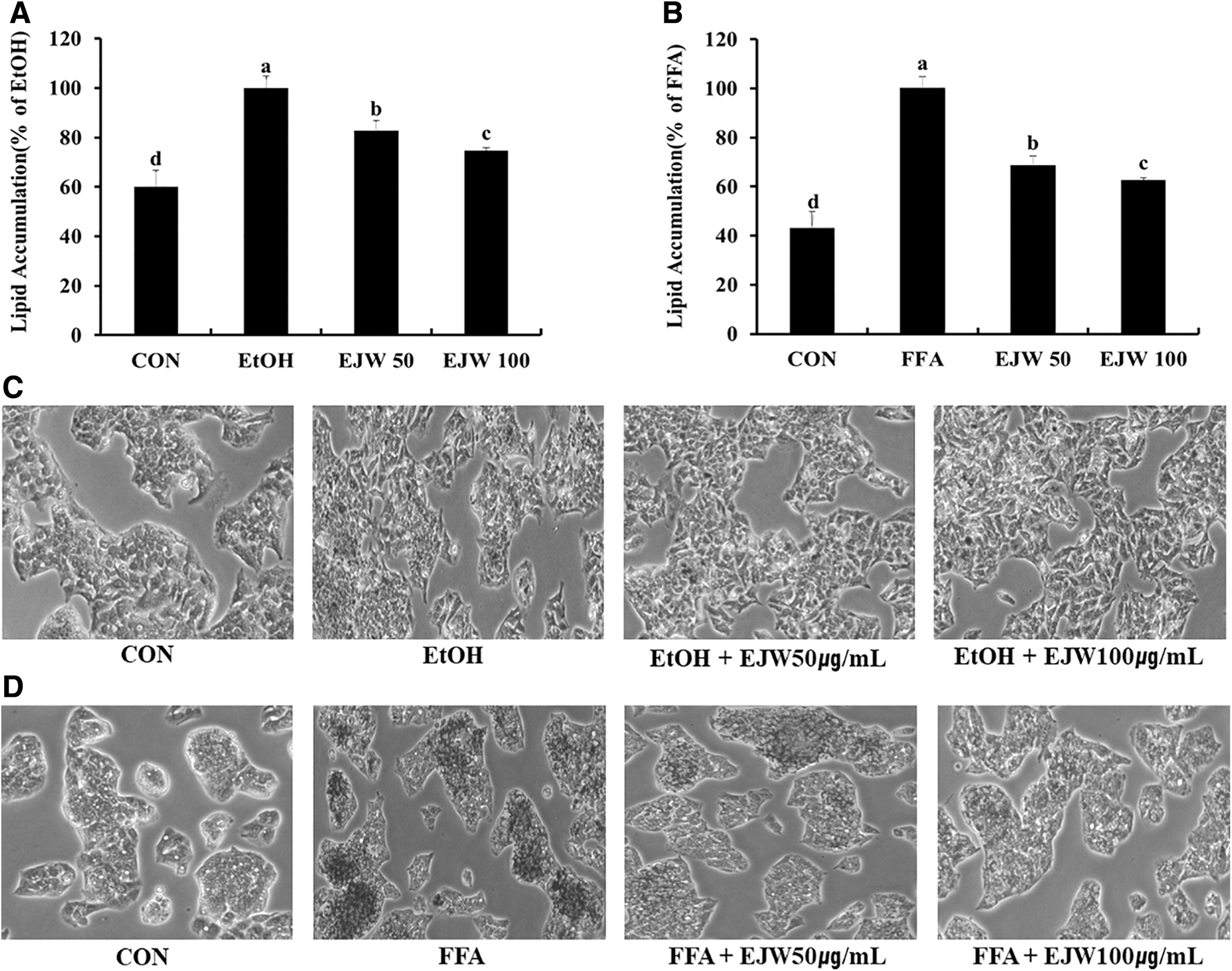
Effect of Eriobotrya japonica (EJW) on lipid accumulation in EtOH-treated cells
Antioxidant activities of EJW in EtOH-treated and FFA-treated cells
As shown in Figure 3, EtOH-treated and FFA-treated groups showed significantly higher intracellular ROS levels than the control group. Intracellular ROS formation was noticeably decreased when the cells were pretreated with 50 or 100 μg/mL EJW compared to those in EtOH-treated and FFA-treated groups (P < .05). As presented in Table 2, treatment with 200 mM ethanol for 3 days or 1 mM FFA mixture for 24 h significantly decreased the activities of antioxidant enzymes, including CAT, SOD, GR, GST, GPx, and GSH, compared to those in the control group. However, pretreatment with 50 or 100 μg/mL EJW inhibited the decreased antioxidant enzyme activities. Also, levels of MDA, a secondary product of lipid peroxidation, after pretreatment with 50 or 100 μg/mL EJW were lower than those in EtOH-treated and FFA-treated groups (P < .05).
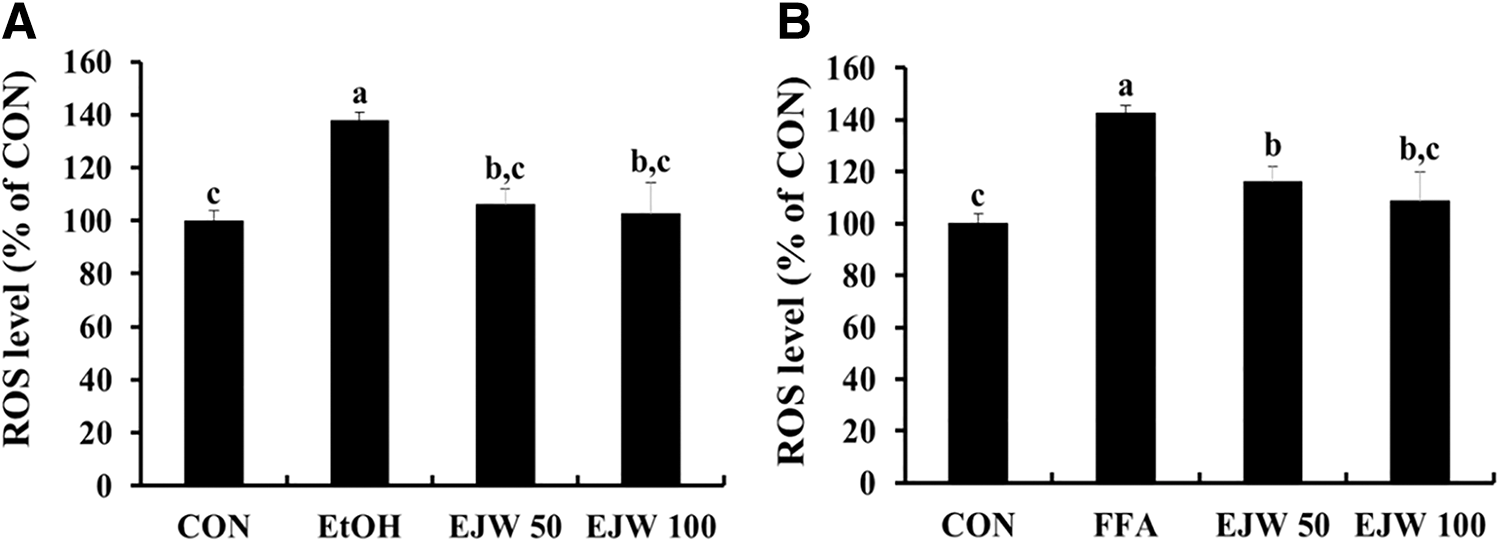
Effect of Eriobotrya japonica (EJW) on intracellular ROS levels in EtOH-treated cells
Effect of the Hot Water Extract of Eriobotrya japonica on Antioxidant Enzyme Activities in EtOH-Treated Cells and Free Fatty Acid-Treated Cells
Data are expressed as mean ± SD (n = 3), and different letters indicate (a > b > c) a significant difference (P < .05), as determined by Duncan's multiple range test. CON, normal control group; EJW50 and EJW100, pretreatment with EJW 50 and 100 μg/mL groups; EtOH, EtOH-treated group; FFA, FFA-treated group.
CAT, catalase; EJW, hot water extract of E. japonica; FFA, free fatty acid; GPx, GSH peroxidase; GR, GSH reductase; GSH, glutathione; GST, glutathione S-transferase; MDA, malondialdehyde; SD, standard deviation; SOD, superoxide dismutase.
Effect of EJW on lipogenesis-related messenger RNA expression in hepatocytes
We investigated the messenger RNA (mRNA) expression of CYP2E1, which is mainly involved in alcohol and FFA metabolism, through real-time PCR. There was a 3.5-fold increase in CYP2E1 expression in the EtOH-treated group and a 2.2-fold increase in the FFA-treated group compared to that in the control group. Furthermore, EtOH treatment in HepG2/2E1 cells and FFA treatment in HepG2 cells markedly increased the expression of de novo lipogenesis factors, while the upregulated expression was decreased by pretreatment with 50 or 100 μg/mL EJW (Fig. 4A, D). 5′Adenosine monophosphate (AMP)-activated protein kinase (AMPK) mRNA expression related to fatty acid β-oxidation was lower in both EtOH-treated and FFA-treated groups than in the control group. However, downregulated expression was increased by pretreatment with 50 or 100 μg/mL EJW compared to that in the EtOH-treated and FFA-treated groups (Fig. 4B, E). In addition, we evaluated mRNA expression levels involved in fatty acid uptake in hepatocytes. There were no significant differences in the mRNA expression of cluster of differentiation 36 (CD36), fatty acid transport protein 2 (FATP2), and FATP5 among the four groups in the AFLD cell model (Fig. 4C). On the other hand, the difference was observed in the NAFLD cell model. The FFA-treated group showed significantly higher CD36, FATP2, and FATP5 mRNA expression levels than those in the control group. Pretreatment with 50 or 100 μg/mL EJW lowered the upregulated expression levels in the FFA-treated group (P < .05, Fig. 4F).
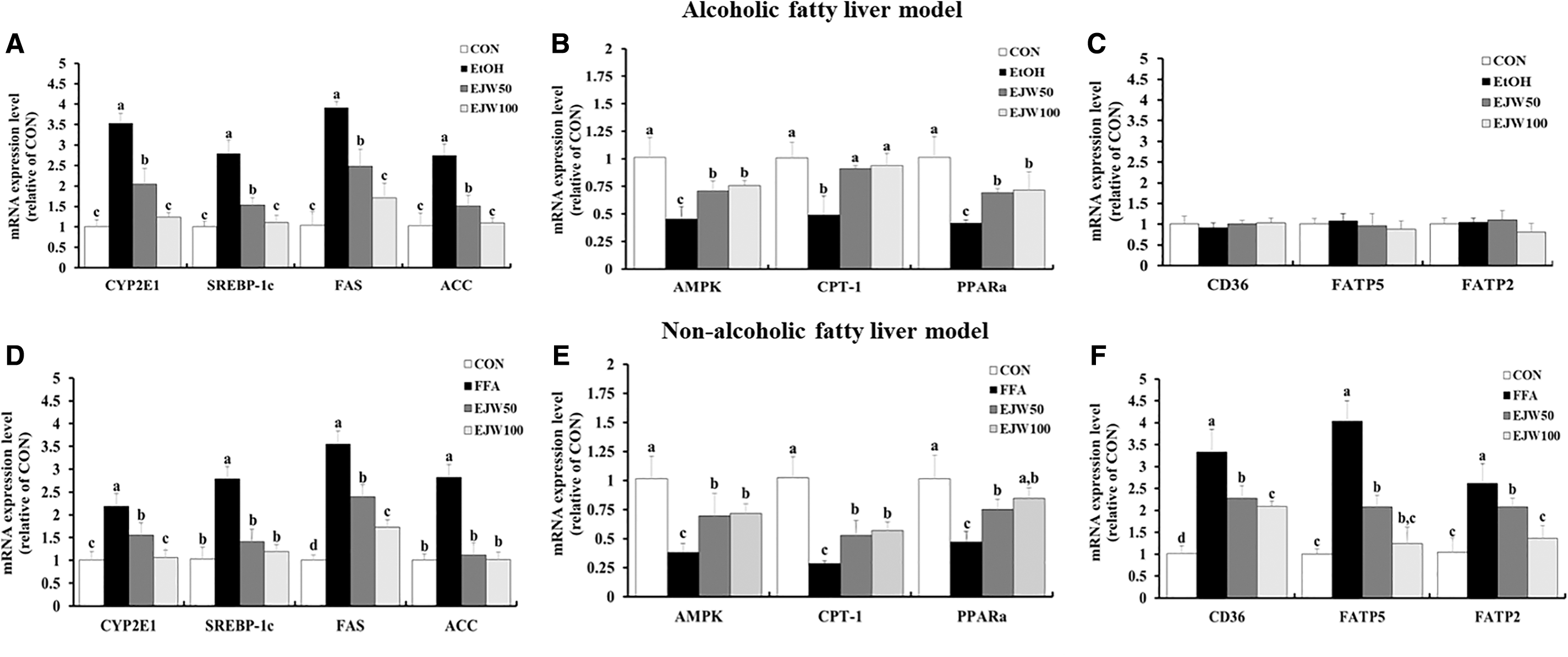
Effects of Eriobotrya japonica (EJW) on fatty acid synthesis-related mRNA expression, including CYP2E1, SREBP-1c, FAS, and ACC
Effect of EJW on AMPK and ACC activation in hepatocytes
To investigate ACC and AMPK activation related to lipogenesis in hepatocytes, we evaluated ACC and AMPK phosphorylation by Western blot assay. The ratios of phosphorylated (p)-ACC to ACC, and p-AMPK to AMPK were significantly decreased in the EtOH-treated and FFA-treated groups. As shown in Figure 5A–C, pretreatment with 50 or 100 μg/mL EJW in HepG2/2E1 cells noticeably increased the p-AMPK:AMPK and p-ACC:ACC ratios compared to those in the control group. Also, the reduced p-AMPK:AMPK and p-ACC:ACC ratios in FFA-treated HepG2 cells were greatly increased by pretreatment with 50 or 100 μg/mL EJW (P < .05, Fig. 5D–F).

Effect of Eriobotrya japonica (EJW) on the phosphorylation ACC and AMPK in ethanol-treated
Discussion
Alcohol can be metabolized by CYP2E1, an enzyme whose expression appears to be 5- to 10-times higher in chronic alcoholism than under normal conditions. 30,31 Our study also showed that CYP2E1 mRNA expression was remarkably higher in excessive EtOH-treated HepG2/2E1 cells than in the control group cells. CYP2E1 activation leads to the overproduction of ROS and increased oxidative stress in several tissues, especially in the liver. 32 In addition, FFAs taken up by liver can stimulate oxidative stress. 33 Therefore, alcohol as well as FFAs have been considered to be substantial risk factors that induce oxidative stress. Oxidative stress is able to regulate various physiological functions when it is present in adequate amounts in the body. 34 However, if production is increased as a result of excessive alcohol or FFA intake, it can cause various metabolic disorders and play an important role in the pathogenesis of AFLD and NAFLD. 35 –38
According to methods described in several studies regarding lipid overloading in vitro, we treated CYP2E1-overexpressing HepG2 cells with EtOH for 3 days to induce AFLD in a cell model. 39,40 Through CYP2E1 transfection, alcohol sensitivity was increased and CYP2E1 expression was stimulated. In the NAFLD cell model, an FFA was used to treat HepG2 cells. In another study, lipid accumulation was efficiently induced through the use of a 2:1 ratio of oleic acid and palmitic acid, and minor cytotoxicity was observed in HepG2 cells. 41 EtOH and FFA treatments produced the largest lipid droplets in cells, as determined by Oil Red O staining. Antioxidants may be valuable in protecting the cell from damage through lipid accumulation and oxidative stress. Several studies have proved that fatty acid synthesis repression in the liver was due to the inhibitory effects of polyphenols, so we hypothesized that EJW would exert preventive effects against AFLD and NAFLD. 42,43 EJW is well known for possessing several phenolic compounds such as catechin and hyperin that display high antioxidant activity. 43
Our results demonstrate that oxidative stress was alleviated by EJW pretreatment during lipid accumulation. Furthermore, EJW pretreatment in HepG2/2E1 and HepG2 cells significantly decreased CYP2E1 mRNA expression compared to that in the excessive EtOH- or FFA-exposed cells. Our study concerned not only EtOH- but also FFA-triggered CYP2E1 mRNA expression. In a recent study, FFA was also metabolized by CYP2E1, and increased CYP2E1 activity was observed in high-fat diet-fed rats and patients with NAFLD compared to that in the appropriate controls. 44 –46 The association between FFA and CYP2E1 is thought to be a factor in the development of NAFLD. 12
EJW pretreatment significantly increased AMPK activation and suppressed ACC activation, which was increased by stimulation with EtOH or FFA. As an energy homeostasis sensor that regulates lipid metabolism, AMPK activity controls ACC activity by phosphorylation and SREBP-1c expression associated with the inhibition of lipogenesis. 47 The principal role of ACC is to provide malonyl-CoA for lipogenesis and inhibit CPT-1 expression, which is a key factor for fatty acid β-oxidation. 48 Also, PPARα regulates fatty acid levels in the liver as one of the essential factors of fatty acid β–oxidation. 49 Our results revealed that CPT-1 and PPARα levels were significantly downregulated compared to those in the control, and SREBP-1c and FAS, which lead to lipogenesis, were markedly upregulated by EtOH or FFA exposure. These observations are consistent with a previous study that indicated ROS caused lipid accumulation through AMPK inhibition and SREBP-1c activation. 28 However, EJW restored the abnormal expression of these factors associated with the development of FLD to normal levels. Therefore, our results provide evidence that EJW may exert potential protective effects against lipid accumulation in hepatocytes.
FATP2 and FATP5 are principal members of the FATP family, and CD36 predominantly regulates fatty acid uptake in the liver. Several studies have demonstrated that elevated expressions of FATPs and CD36 contribute to an increased fatty acid uptake and an amplified lipid accumulation in hepatocytes, closely related to NAFLD, 50,51 which in turn fatty acid uptake factors may represent potential targets for the lipid accumulation. 52,53 Our study showed that FATPs and CD36 expression levels were significantly increased in the FFA treatment model and these increased levels were suppressed by EJW pretreatment. However, EtOH treatment had no significant effect on expressions of these fatty acid uptake factors. In earlier studies, CD36 did not affect the fatty acid uptake in hepatocytes under normal conditions, but this was actively functioned when excessive fatty acids were presented by high-fat intake or obesity condition. 54,55 In addition, CD36-deficient mice fed with alcohol diet were protected against AFLD, although this result was independent of altered hepatic fatty acid uptake. 56 Our observations suggest that direct exposure to excess fatty acids increased fatty acid uptake in hepatocytes, inducing fat accumulation, but not affecting alcohol exposure.
In conclusion, our results indicate that EJW significantly inhibited oxidative stress by decreasing intracellular ROS formation and increasing the activities of antioxidant enzymes. Also, it prevented lipid accumulation by restoring lipid metabolism by balancing FFA synthesis and β-oxidation. Furthermore, fatty acid uptake was significantly modulated by EJW pretreatment. Therefore, EJW has a potential function in suppressing the development of FLD in both EtOH-induced and FFA-induced models.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2017R1D1A3B03035177).