
Abstract
The major aim of this study was to investigate the effect of rice protein (RP) on the activation of endogenous antioxidant defense in growing and adult rats. After 2 weeks, RP activated nuclear factor erythroid 2 (NF-E2)-related factor 2 (Nrf2) by suppressing Kelch-like ECH-associated protein 1 (Keap1) and Cullin 3 (Cul3) in growing and adult rats. Compared with casein, the upregulation of antioxidant responsive element (ARE)-driven antioxidant expression levels (glutamate cysteine ligase catalytic subunit, glutamate cysteine ligase modulatory subunit, glutathione synthase, glutathione reductase, glutathione S-transferase, glutathione peroxidase, catalase, superoxide dismutase, heme oxygenase 1, NAD(P)H:quinone oxidoreductase 1) were found in RP groups. Also, RP upregulated methionine sulfoxide reductase (MsrA, MsrB2, and MsrB3) expression levels in growing and adult rats. As a result, RP enhanced endogenous antioxidative capacities to reduce hepatic accumulations of malondialdehyde, protein carbonyl, and reactive oxygen species. This study demonstrates that RP exerts the endogenous antioxidant capacity in growing and adult rats, which is due to stimulating Msr antioxidant expression and activating Nrf2-ARE pathway. Results suggest that the antioxidant activity induced by RP is independent of age.
Introduction
Dietary antioxidants are suggested to exert a free radical-scavenging capacity to reduce oxidative stress. 1 –5 Some studies suggest that the antioxidant effect of protein is attributed to its amino acid profiles, in which sulfur-containing amino acids (SAAs) appear to play a major role in inducing antioxidant response. 6,7 Cysteine (Cys) and methionine (Met) are major constituents of SAAs. 8 –10 Cys is a limiting factor for glutathione (GSH) synthesis. 11,12 GSH is an endogenous antioxidant for scavenging reactive oxygen species (ROS). 11 –13 Met is another major source for GSH synthesis through its conversion into Cys. 14,15 Recently, our study provided direct evidence to determine that Met could suppress ROS accumulation through increasing GSH synthesis and stimulating methionine sulfoxide reductase (e.g., MsrA, MsrB) expression. 16 Thus, SAAs can regulate two major endogenous antioxidant systems, which are GSH and methionine sulfoxide reductase. 17
Rice protein (RP), which is rich in SAAs, 18 –20 can exert antioxidant activity through regulating GSH metabolism and scavenging free radicals. 6,7,19 –21 Recently, Li et al. have reported that RP could increase nuclear factor erythroid 2 (NF-E2)-related factor 2 (Nrf2) expression in adult rats, suggesting RP might have a potential for activating Nrf2. 22 Nrf2 is a key transcription factor for endogenous antioxidant mechanism. 22 –26 However, some questions are left and unanswered. Mostly, the convincing evidence on whether RP can stimulate Msr expression through activating the Nrf2 pathway is lacking.
Age is a major factor leading to accelerated oxidative stress. 27 Thus, growing and adult rats were used in this study to investigate: (1) whether and how does RP exert an endogenous antioxidant capacity and (2) whether age affects the antioxidant activity of RP?
Materials and Methods
Protein sources
In this study, consistent with our previous studies, 22,28 RP and casein (CAS) were also used as dietary protein sources. RP from Oryza sativa L. cv. Longjing 20 was provided by Rice Research Institute of Heilongjiang Academy of Agricultural Sciences (Jiamusi, China). CAS was purchased from Gansu Hualing Industrial Group (Gansu, China).
Animals and diets
As with our previous studies, 16,22,23,28 this study was performed in conformity with the Guidelines of the Committee for the Experimental Animals of Harbin Institute of Technology (SCKK2012-0001, Harbin, China). In brief, 7-week-old male Wistar rats (body weight 200–220 g, designated growing rats) and 20-week-old male Wistar rats (body weight 380–400 g, designated adult rats) were purchased from the Vital River Laboratories (Beijing Vital River Laboratory Animal Technology Co. Ltd., Beijing, China) and individually housed in metabolic cages in a room maintained at 22°C ± 2°C under a 12 h light–dark cycle (07:00–19:00 for light).
In this study, four groups, consisting of six animals per group, were used for investigation. The CAS fed group (a major animal protein) was treated as a control. CAS-G (fed to growing rats) and CAS-A (fed to adult rats), respectively, contained 228.9 g CAS/kg diet and 160.2 g CAS/kg diet, according to the formula recommended by AIN-93. 29 Also, RP-G (fed to growing rats) and RP-A (fed to adult rats), respectively, contained 220.4 g RP/kg diet and 154.3 g RP/kg diet, according to the formula recommended by AIN-93. 29
Hepatic oxidative status
The measurements of ROS, malondialdehyde (MDA), and protein carbonyl (PCO) in the livers were performed as previously described. 16,22,23,28
Antioxidant capacity
The analyses of total antioxidative capacities (T-AOC), GSH contents, the activities of glutamate cysteine ligase (GCL), glutathione peroxidase (GPx), glutathione reductase (GR), glutathione S-transferase (GST), catalase (CAT), and total superoxide dismutase (T-SOD) were also performed as previously described, 16,22,23,28 and according to the methods described in the commercial kits from Nanjing Jiancheng Bioengineering Institute (Nanjing, China).
Quantitative real-time PCR
The quantitative real-time PCR was measured in this study as previously described. 16,22,23,28 In brief, using the TRIzol reagent kit (Invitrogen, Carlsbad, CA, USA) and the PrimeScript™ 1st strand cDNA Synthesis Kit (Takara Bio, Inc., Otsu, Shiga, Japan), total RNA was extracted from the livers and cDNA was reverse transcribed and normalized to the mRNA level of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The primer sequences used are shown in Table 1. In this study, the relative mRNA levels in groups CAS-G and CAS-A were, respectively, set as 1.00.
Primers for Quantitative Real-Time PCR
Protein extraction and Western blotting analysis
The total, cytoplasmic, and nuclear proteins were used for Western blot analysis, which were prepared as described in our previous studies. 16,22,23,28 The primary antibodies of Nrf2, Kelch-like ECH-associated protein 1 (Keap1), Cullin 3 (Cul3), MsrA, MsrB2, MsrB3, glutamate cysteine ligase catalytic subunit (GCLC), glutamate cysteine ligase modulatory subunit (GCLM), glutathione synthase (GS), GR, GSTA1, SOD, CAT, heme oxygenase 1 (HO-1), NAD(P)H:quinone oxidoreductase 1 (NQO1), H1.2, and GAPDH were purchased from Proteintech (Wuhan, China), and methionine adenosyltransferase Iα (MAT1A), S-adenosylhomocysteine hydrolase (SAHH), cystathionine β-synthase (CBS), cystathionine γ-lyse (CTH), GPx1/2, and second antibody were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). In this study, the relative protein expression in groups CAS-G and CAS-A was, respectively, set as 1.00.
Statistical analysis
Data are expressed as the mean ± SEM. Differences between groups were examined for statistical significance using one-way analysis of variance (ANOVA) followed by the least significant difference test. The criterion for significance was P < .05.
Results
Oxidative damage
As given in Table 2, compared with CAS-A (18.60 ± 0.63) and CAS-G (15.35 ± 0.55), hepatic accumulations of ROS were suppressed by RP-A to 14.00 ± 0.46, i.e., 24.73% (P < .05) and by RP-G to 12.89 ± 0.31, i.e., 16.03% (P < .05). RP-A and RP-G significantly decreased hepatic contents of MDA to 3.95 ± 0.17, i.e., 29.59% in adult rats (P < .05) and to 3.07 ± 0.10, i.e., 18.57% in growing rats (P < .05). PCO was also decreased by RP-A to 4.38 ± 0.22, i.e., 34.23% in adult rats (P < .05) and by RP-G to 3.17 ± 0.13, i.e., 12.19% in growing rats (P < .05).
Hepatic Antioxidant Capacity and Hepatic Contents of Reactive Oxygen Species, Malondialdehyde, and Protein Carbonyl
Values are means ± SEM (n = 6).
P < .05, in comparison with CAS-A.
P < .05, in comparison with CAS-G.
CAS, casein; CAT, catalase; MDA, malondialdehyde; PCO, protein carbonyl; RP, rice protein; ROS, reactive oxygen species; T-AOC, total antioxidative capacities; T-SOD, total superoxide dismutase.
Antioxidant capacity
The activities of CAT and T-SOD were significantly higher in RP-A and RP-G than in CAS-A and CAS-G (P < .05, Table 2). Similarly, pronounced increases of T-AOC were observed in RP-A and RP-G as compared with CAS-A and CAS-G (P < .05).
Hepatic GSH
As shown in Figure 1A, GSH concentrations were increased in RP-A to 5.57 μg/mg ±0.18, i.e., increasing 35.85% in adult rats (P < .05), and in RP-G to 3.94 μg/mg ±0.15, i.e., increasing 20.86% in growing rats (P < .05), as compared with CAS-A (4.10 ± 0.15) and CAS-G (3.26 ± 0.11).

Effects of RP on hepatic contents of GSH and activities of GCL, GST, GR, and GPx in growing and adult rats.
GSH metabolism-related enzyme activity
As shown in Figure 1B, the activity of GCL was significantly increased by RP-A and RP-G (P < .05). Hepatic activities of GST (Fig. 1C) and GR (Fig. 1D) were significantly higher in RP-A and RP-G than in CAS-A and CAS-G (P < .05). GPx activity (Fig. 1E) was not significantly different between CAS-A and RP-A (P > .05) or between CAS-G and RP-G (P > .05).
Msr expression
Compared with CAS-A and CAS-G, protein and mRNA levels of total MsrA was, respectively, stimulated by RP-A and RP-G (P < .05, Fig. 2A). Significantly,

Effects of RP on hepatic expression levels of MsrA, MsrB2, and MsrB3 in growing and adult rats.
The protein and mRNA levels of total MsrB2 (Fig. 2C) and total MsrB3 (Fig. 2D) were higher in RP-A and RP-G than in CAS-A and CAS-G (P < .05). However, neither MsrB2 (Fig. 2E) nor MsrB3 (Fig. 2F) induced a nuclear translocation in RP-A and RP-G rats after 2 weeks feeding, although cytosolic MsrB2 (Fig. 2G) and cytosolic MsrB3 (Fig. 2H) were higher in RP-A and RP-G rats.
Nrf2 activation
Compared with CAS-A and CAS-G, RP-A and RP-G significantly decreased the mRNA level and protein expression level of Keap1 (Fig. 3A) and Cul3 (Fig. 3B) (P < .05). RP-A and RP-G significantly increased the gene and protein expression of total Nrf2 (Fig. 3C, P < .05). Also, RP-A and RP-G significantly increased protein expression levels of nuclear Nrf2 and cytosolic Nrf2 (Fig. 3D, P < .05).
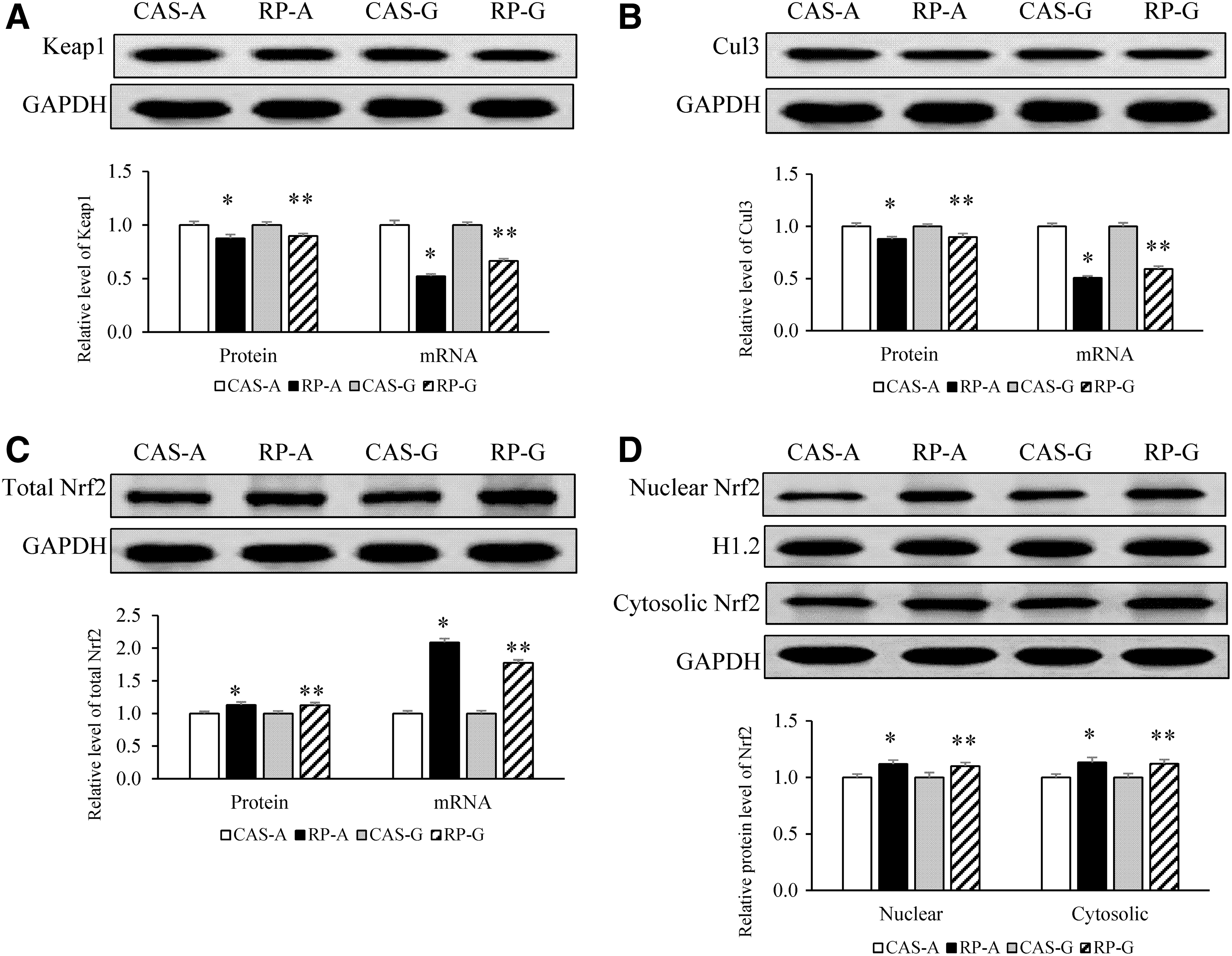
Effects of RP on hepatic expression levels of Keap1, Cul3, and Nrf2 in growing and adult rats.
Expression levels of GCLC, GCLM, and GS
Compared with CAS-A and CAS-G, RP-A and RP-G significantly upregulated the gene and protein expression levels of GCLC (Fig. 4A, P < .05) and GCLM (Fig. 4B, P < .05). RP-A and RP-G markedly increased the RNA levels of GS (Fig. 4C, P < .05). RP-A significantly upregulated protein expression of GS (P < .05, Fig. 4C). However, RP-G did not produce a significant increase in GS protein level as compared with CAS-G (P > .05, Fig. 4C).

Effects of RP on hepatic expression levels of GCLC, GCLM, GS, GST, GR, and GPx in growing and adult rats.
Expression levels of GST, GR, and GPx
As shown in Figure 4D, the gene and protein expression levels of GST were significantly stimulated by RP-A and RP-G (P < .05). RP-A and RP-G significantly enhanced the mRNA levels of GR (P < .05, Fig. 4E). A significant increase in protein expression of GR was found in RP-A (P < .05, Fig. 4E) rather than in RP-G (P > .05, Fig. 4E). In this study, although RP-A and RP-G significantly upregulated gene expression of GPx (P < .05, Fig. 4F), the marked increase in protein expression of GPx1/2 was not produced by RP-A and RP-G (P > .05, Fig. 4F).
Expression levels of CAT, SOD, HO-1, and NQO1
Upon activation of Nrf2, the mRNA levels of CAT (Fig. 5A), SOD (Fig. 5B), HO-1 (Fig. 5C), and NQO1 (Fig. 5D) were significantly augmented by RP-A and RP-G, respectively (P < .05). RP-A significantly upregulated protein expression levels of CAT, SOD, and HO-1 (P < .05), whereas RP-G did not (P > .05, Fig. 5A–C). There were no significant differences of NQO1 protein levels (Fig. 5D) among all groups (P > .05, Fig. 5D).

Effects of RP on hepatic expression levels of CAT, SOD, HO-1, and NQO1 in growing and adult rats.
Expression levels of MAT1A, SAHH, CBS, and CTH
RP-A and RP-G significantly increased gene expression levels of MAT1A (Fig. 6A), SAHH (Fig. 6B), CBS (Fig. 6C), and CTH (Fig. 6D), respectively (P < .05). RP-A upregulated protein expression levels of MAT1A (Fig. 6A), SAHH (Fig. 6B), and CTH (Fig. 6D) (P < .05), whereas RP-G did not (P > .05). Significantly, as illustrated in Figure 6C, hepatic protein levels of CBS were stimulated by both RP-A and RP-G (P < .05).

Effects of RP on hepatic expression levels of MAT1A, SAHH, CBS, and CTH in growing and adult rats.
Discussion
Msr is a ubiquitous enzyme involved in endogenous antioxidant defense and acts as an ROS scavenger. 16,17 This study supports the observation RP upregulates the expression of MsrA, MsrB2, and MsrB3 to potentiate ROS-scavenging action. These results were supported by previous studies and our recent findings, 16,17,30 –32 suggesting that RP could exert an endogenous antioxidant activity through stimulating the Msr endogenous antioxidant system.
Accordingly, these results lead to the question of how RP can stimulate the Msr antioxidant system after 2 weeks feeding. The fact that RP is rich in SAAs may provide a clue. 18 –20 It has been demonstrated that SAAs can exert antioxidant effects, 16,17,33 and there were the markedly positive correlations between the SAAs consumption and the expression levels of MsrA (r = 0.8890, P < .05), of MsrB2 (r = 0.8314, P < .05), and of MsrB3 (r = 0.8181, P < .05) in this study. Thus, it is not surprising that higher level of SAAs in RP (RP, 44.42 μg/mg; CAS, 32.64 μg/mg) could stimulate the Msr antioxidant system, highlighting a role of SAAs in inducing endogenous antioxidant capacity of RP.
Another question requiring explanation is how could RP induce this endogenous antioxidant activity? One explanation involves the activation of Nrf2-antioxidant responsive element (ARE) antioxidant system by RP. This study clearly indicated the markedly positive correlations between the nuclear Nrf2 and protein expression levels of MsrA (r = 0.9109, P < .05), of MsrB2 (r = 0.7779, P < .05), and of MsrB3 (r = 0.7824, P < .05). Therefore, this study suggests that the activation of Nrf2 might be linked to the stimulation of Msr by RPs.
Upon activation of Nrf2, in addition to the upregulations of CAT, SOD, HO-1, NQO1, GR, GST, and GPx, which are involved in the endogenous antioxidant system, the marked stimulations of GSH synthetic enzymes were also induced by RP-G and RP-A. There was a markedly positive correlation between nuclear Nrf2 and the expression levels of GCLC (r = 0.8136, P < .05), of GCLM (r = 0.8313, P < .05), and of GS (r = 0.7808, P < .05), respectively. These findings, combined with the facts that RP stimulated Msr expression, suggest that the endogenous antioxidant capacity exerted by RP through Nrf2-ARE pathway might be a synergistic action rather than a solely antioxidant response.
In this study, it must be emphasized that age had an effect on the antioxidant capacity of RP. It is known that age is associated with increased oxidative stress, causing an accumulation of oxidative damage and a decrease in activation of Nrf2. 27,34 However, worthy of note, in this study, the increased age did not reduce antioxidant capacity in adult rats fed with RP-A in comparison with RP-G. Instead, stronger antioxidant action was found in RP-A than in RP-G. These results were consistent with our other recent findings. 28 Thus, although we cannot fully explain these findings, but observations that Msr expression can extend life span are especially relevant to these results. 35,36 Thus, it is especially interesting that Msr expression levels were stronger in adult rats fed with RP-A than in growing rats fed RP-G. Furthermore, another view that the native amino acid composition of RP may be a major regulator in oxidative/antioxidative status further supports our present findings. 6,7 Accordingly, the fact that the increased age could not attenuate the RP-A-induced endogenous antioxidant response in adult rats is especially interesting.
To further elucidate the antioxidant mechanism exerted by RP, the role of SAAs in the antioxidant system was also investigated in this study. MAT, SAHH, CBS, and CTH play important roles in the conversion of Met into Cys for GSH synthesis. 8,9,14,15 To support this view, in addition to the marked increases in the expression levels of GCLC, GCLM, and GS, the upregulated expression levels of MAT, SAHH, CBS, and CTH further confirmed that GSH synthesis could be enhanced by RP feeding, in which higher level of SAAs might be a contributor. More significantly, the present results showed the significantly positive correlations between consumption of SAAs and nuclear contents of Nrf2 (r = 0.7852, P < .05), as well of nuclear MsrA (r = 0.8952, P < .05), suggesting that SAAs may be a modulator responsible for the stimulation of endogenous antioxidant mechanisms through the Nrf2 pathway. These results are consistent with the study by Wang et al., which demonstrated that Met can exert endogenous antioxidant capacity to improve oxidative stress through the Nrf2-ARE antioxidant system. 16 Moreover, some studies have also highlighted an important role of transsulfuration pathway for GSH synthesis through the Nrf2 pathway. 37 However, it must be noted that the endogenous antioxidant capacity exerted by RP cannot be solely attributed to the effect of SAAs, other amino acids may also contribute to the antioxidant activity of RP. Our recent findings indicate that arginine can produce antioxidant activity through increasing GSH synthesis and stimulating Nrf2 antioxidant system. 23 In light of this view, higher level of arginine in RP (RP, 88.17 μg/mg; CAS, 33.33 μg/mg) might be another contributor to the endogenous antioxidant response induced by RP. In addition, besides SAAs and arginine, another factor of RP, which might be involved in antioxidant response (e.g., another amino acid profile, special peptide, etc), should also be investigated in further studies.
In conclusion, this study is the first to demonstrate that RP exerts the endogenous antioxidant capacity in growing and adult rats through stimulating the Msr antioxidant system and activating the Nrf2-ARE antioxidant pathway (Fig. 7). The novel findings are that the endogenous antioxidant capacity exerted by RP is independent of age. Instead, it might be dependent on the amino acids profile of native RP. Clearly, more studies are needed to elucidate the precise antioxidant mechanisms provided by RP.
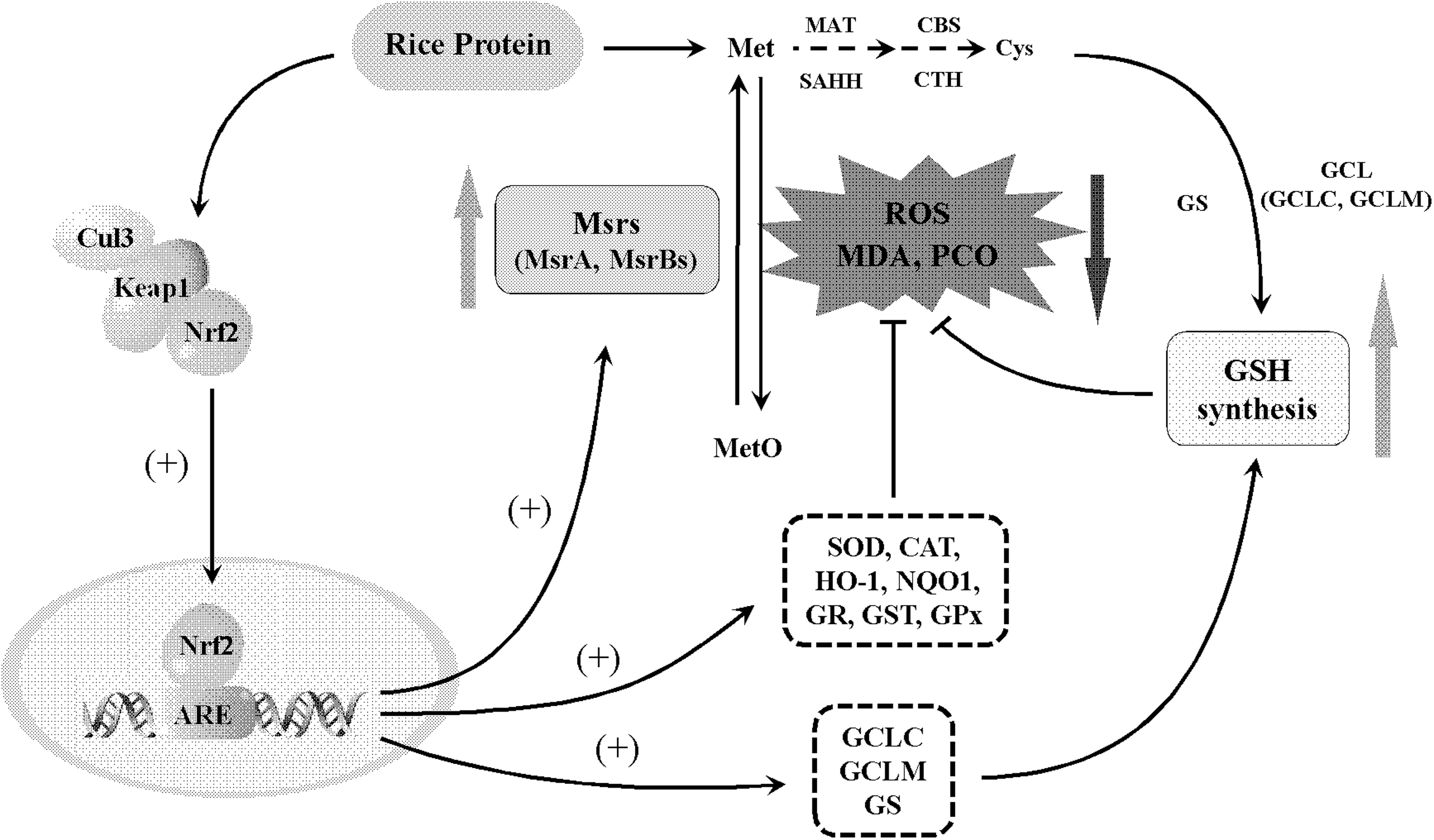
RP exerts endogenous antioxidant capacity through stimulating methionine sulfoxide reductase antioxidant system and activating Nrf2-ARE pathway. ARE, antioxidant responsive element; Met, methionine; MetO, methionine sulfoxide.
Footnotes
Acknowledgment
We thank Rice Research Institute of Heilongjiang Academy of Agricultural Science for providing rice grains.
Author Disclosure Statement
The authors declare that there is no conflict of interest.
Funding Information
We are grateful to the National Natural Science Foundation of China for supporting this study (Grant No. 31371755).