
Abstract
Temporal lobe epilepsy (TLE) is the most common form of localization-related epilepsy, with the highest prevalence rate in adulthood. Recently, we reported the beneficial effects of the individual treatment with flavonoids such as silibinin and morin in kainic acid (KA)-treated mouse model for TLE. In this study, we investigated whether there is a synergistic effect of co-treatment with silibinin and morin on the susceptibility to seizure, the frequency of spontaneous recurrent seizures (SRSs), and granule cell dispersion in the dentate gyrus, which could be partially controlled by treatment with each flavonoid in the animal model for TLE. Unfortunately, we did not observe any synergistic effect against the susceptibility of seizure and SRS induced by KA treatment. However, the combination of these flavonoids showed similar antiepileptic effects compared with treatment with each one individually. Therefore, although silibinin and morin are not suitable for combination therapy, our results still suggest that these flavonoids can be used as potent therapeutic compounds for preventing epileptic seizures.
Temporal lobe epilepsy (TLE), the most common type of epilepsy, is a chronic neurological disorder characterized by recurrent seizures 1 and is frequently accompanied by neuropathological features, such as hippocampal sclerosis and granule cell dispersion (GCD). GCD is described as an abnormal extension of the granule cell layer (GCL) and was found in ∼50% of cases of hippocampal sclerosis in patients with TLE. 2 In addition, GCD-related morphological abnormalities are known to be closely related to spontaneous seizures and status epilepticus. 3
Although many antiepileptic drugs have been developed for the treatment of epilepsy, 30% of patients still experience uncontrolled epilepsy, associated comorbidities, and various side effects, such as drowsiness, unsteadiness, dizziness, and greater risk of infection and bleeding. Thus, there is a critical need for effective and safe alternative drugs for the treatment of epilepsy. 2,4 Flavonoids, important secondary plant metabolites with various biological activities, have recently attracted attention as an alternative to synthetic compounds. 2,5 There is considerable evidence that flavonoids and their metabolites are able to cross the blood–brain barrier and enter the brain, resulting in neuroprotection against neuronal death. 6 –8 For instance, silibinin has been reported to improve the abnormal locomotor activity, learning, and spatial memory through its neuroprotective effects inhibiting the inflammatory responses, oxidative stress, and neuronal death in Parkinson's disease and Alzheimer's disease. 9 –11 Morin protects cells from oxidative damages associated with neurodegenerative diseases. 12 In addition, we recently reported that each treatment with silibinin and morin against epileptic seizures can induce the attenuation of GCD in the dentate gyrus (DG) and exert neuroprotective effects through decreasing the release of proinflammatory molecules in the kainic acid (KA)-exposed hippocampus, resulting in the suppression of epileptic seizures. 2,4
The combined application of bioactive flavonoids could result in more beneficial therapeutic outcomes against various diseases. Advantageous outcomes by the synergistic effects include the increased therapeutic efficacy, decreased adverse effects, and reduced development of drug resistance. 13 Treatment with morin combined with etoposide synergistically increased the area under the curve of etoposide in blood plasma, and enhanced the distribution of etoposide into tissues by inhibiting the gut and hepatic cytochrome P4503A enzyme. 14 Similarly, treatment with a combination of curcumin, vorinostat, and silibinin yielded synergistic neuroprotective effects against amyloid beta-induced toxicity in vitro. 15 Based on these results, this study examined whether combined treatment with 200 mg/kg silibinin and 80 mg/kg morin, which showed the beneficial effects such as inhibition of GCD and neurotoxic events by each treatment in the KA-treated hippocampus in vivo, 2,4 can produce synergistic effects in seizure susceptibility, and frequency of spontaneous recurrent seizures (SRSs) and GCD in a KA-treated mouse model for TLE.
For the animal model of TLE, seizures were triggered by a unilateral injection of KA (0.2 μg/2 μL) into the hippocampus, at the following anteroposterior (AP), mediolateral (ML), and dorsoventral (DV) coordinates (AP: −2.0 mm; ML: −1.2 mm; DV: −1.5 mm, relative to bregma) using a stereotaxic apparatus (David Kopf Instruments, Tujunga, CA, USA). 4 No mortality was observed in KA-treated animals. All experiments involving animals were performed in accordance with the guidelines approved by the Animal Care Committee of Kyungpook National University (no. KNU 2016-42). All values are expressed as mean ± standard error of the mean. Differences between two independent groups were analyzed with Student's t-tests. Multiple comparisons among the groups were performed using one-way analysis of variance (one-way ANOVA) followed by Tukey's post hoc test. All statistical analyses were performed using SigmaPlot 12.0 software (Systat Software, San Leandro, CA, USA).
To evaluate whether the combination of morin and silibinin can act synergistically against KA-induced seizures, 80 mg/kg morin and 200 mg/kg silibinin were coadministrated orally and intraperitoneally, respectively, 1 h and 1 day before KA injection. As shown in Figure 1, all mice in the KA-injected group experienced status epilepticus within 100 min after KA injection. Compared with KA treatment alone, each treatment with silibinin or morin significantly delayed seizure onset, which is similar to the results of our previous studies (Fig. 1A; ***P < .001 and **P < .01). 2,4 Compared with KA alone, cotreatment with silibinin and morin also delayed the onset of seizures induced by KA injection (Fig. 1A; ***P < .001).

Effect of co-treatment with silibinin and morin on seizures induced by KA treatment.
Furthermore, we examined whether the combined treatment with silibinin and morin was synergistic or not on the suppression of chronic spontaneous seizures induced by KA exposure. To assess SRSs, co-treatment with these flavonoids was initiated 1 day before KA injection and was continued once daily for 35 days. Our results showed that co-treatment with silibinin and morin as well as an individual treatment with these flavonoids effectively decreased the frequency of chronic spontaneous seizures that were induced by KA treatment (Fig. 1B; ***P < .001, **P < .01 and ***P < .001). However, the combination of silibinin and morin showed no synergistic activity on seizure onset and chronic spontaneous seizures induced by KA compared with treatment with each individual flavonoid.
Recent studies have presented evidence that GCD, which is a structural alteration of the GCL, develops in ∼50% of patients with TLE 16,17 and it has been suggested that the restriction of GCD could be useful for preventing and inhibiting the development of chronic epilepsy. To investigate the inhibitory effect of co-treatment with silibinin and morin on the progression of GCD induced by KA exposure, we examined morphological changes in granule cells using Nissl staining 7 days after KA injection (the early stages of epileptogenesis). GCD was expressed as a percentage of GCL width in comparison with the control group after measuring the average width of GCL in the mid and medial quarter portions of the upper blade of the DG. 4 As reported previously, 2,4 KA induced a mild widening of the GCL in the DG. However, both co-treatment and single treatment with silibinin and morin resulted in a significant reduction of GCD (Fig. 2A). Quantitative analysis of GCD showed that KA treatment significantly increased GCD by 140% compared with the control group (Fig. 2B; ***P < .001). Conversely, no morphological changes were observed in the GCL of mice treated with phosphate-buffered saline and silibinin+morin compared with the control group. In particular, compared with KA treatment alone, single treatment with silibinin or morin dramatically inhibited KA-induced GCD (Fig. 2B; ### P < .001 and ## P < .01, respectively).
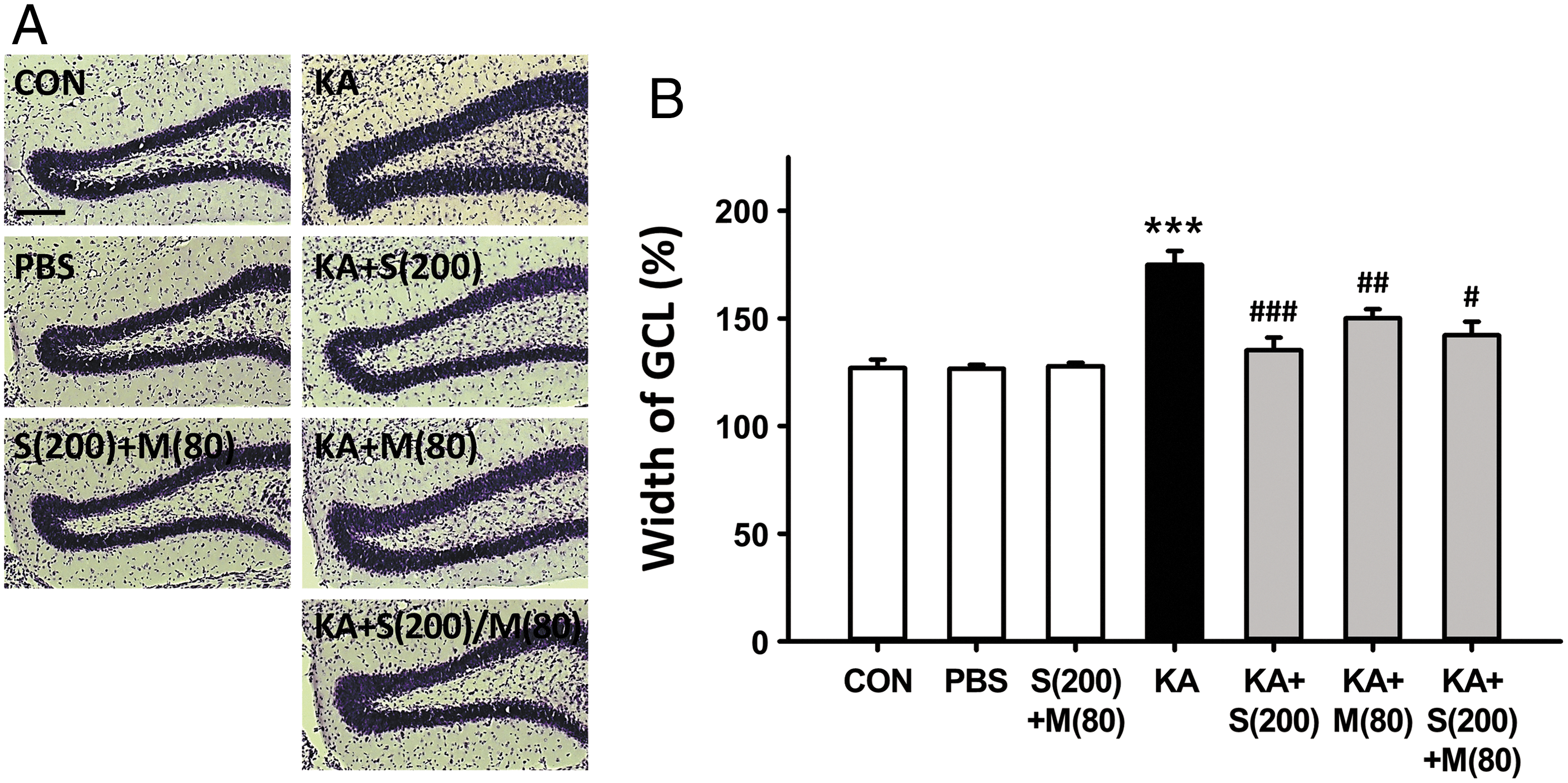
Inhibitory effect of co-treatment with silibinin and morin on the progression of GCD induced by KA in the early stages of epileptogenesis.
Next, cotreatment with these flavonoids led to a significant decrease in GCD compared with KA treatment alone (Fig. 2B; # P < .05). In addition, we investigated the effect of cotreatment with silibinin and morin against KA-induced GCD 35 days after KA injection (the later stages of epileptogenesis). GCL widening was more pronounced with the increase in time in the KA-injected group than in the control group. However, all flavonoid-treated groups showed a significant reduction in GCD compared with the KA-injected group. Quantitative analysis of GCD revealed that it was dramatically increased to 405% in the KA-injected group when compared with the control group (Fig. 3B; ***P < .001). Conversely, compared with KA treatment alone, co-treatment and single treatment with silibinin and morin significantly decreased GCL width (Fig. 3B; ### P < .001, ### P < .001 and ### P < .001, respectively), resulting in consequential development of epileptic seizures. 3 No changes in GCD were observed in any group treated with flavonoids during the process of epileptogenesis. Although our results show that the combination of silibinin and morin may not have any added antiepileptic benefit for seizures because the combined treatment did not yield superior results compared with individual treatment, our observations demonstrate that co-treatment of these flavonoids also has beneficial effects for antiepileptic seizures, such as inhibition of the susceptibility of seizure, SRSs, and GCD shown in the KA-treated animal model of TLE.
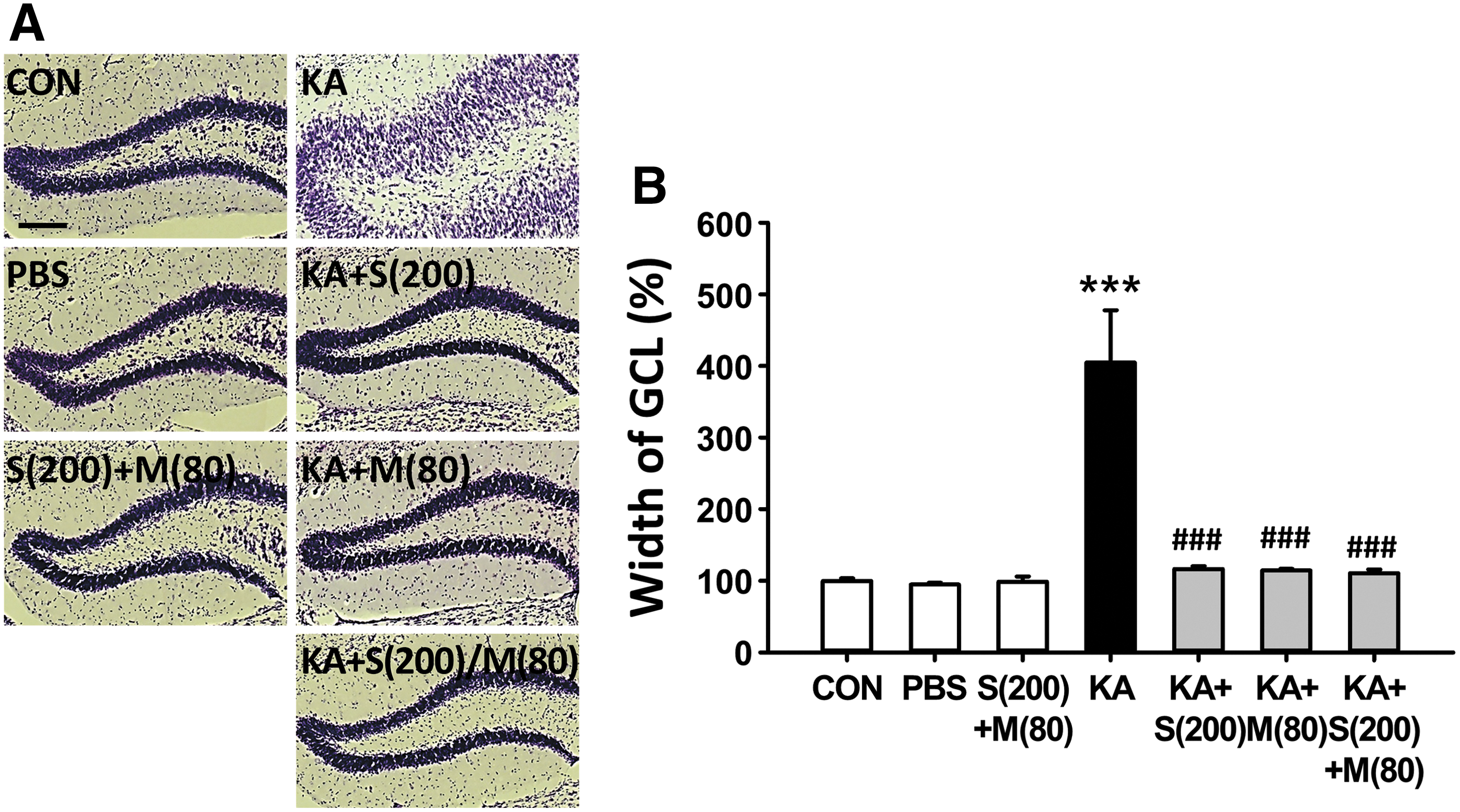
Inhibitory effect of co-treatment with silibinin and morin on the progression of GCD induced by KA in the late stages of epileptogenesis.
Since the identification of the organic anion transporter (OAT) family, there is evidence to suggest that OAT-mediated transport could affect the distribution of flavonoid glucuronides in the brain, 18,19 whereas another study has shown that flavonoids are conjugated to form glucuronidated metabolites by phase II enzymes. 20 In addition, one of the major findings of recent studies indicates that both silibinin and morin competitively inhibit the human OAT1 (hOAT1)-mediated cellular uptake of [3H]-para-aminohippuric acid in Madin–Darby canine kidney cells overexpressing hOAT1. 21 Although further research is needed to determine whether these cellular events are associated with the nonsynergistic antiepileptic effects of co-treatment with silibinin and morin in the KA-treated animal model for TLE, this report suggests that silibinin and morin can be antagonistic in an animal model for epilepsy when administered in combination.
In summary, we investigated whether co-treatment with silibinin and morin could have synergistic effects compared with treatment with alone, which previously reported that each treatment could have neuroprotective effects such as anti-apoptosis and anti-inflammation against KA-induced epileptic seizures in vivo. 2,4 Unfortunately, our results show that there was no significant synergistic effect of co-treatment with these flavonoids compared with individual treatment against the susceptibility of seizure, SRSs, and GCD induced by KA exposure. However, although silibinin and morin may not be appropriate for combination therapy against epileptic seizures, our results still suggest that these flavonoids can be individually and collectively used as potent therapeutic compounds for preventing epileptic seizures (Fig. 4).

Schematic representation of the combined effects of silibinin and morin against epileptic seizures.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
FUNDING INFORMATION
This research was supported by grants from the National Research Foundation of Korea (NRF-2017R1D1A1B03031155 and NRF-2017R1A2B4002675) by the Korean government.