
Abstract
Honey bee larva powder (HLP) has traditionally been used as a daily supplement and tonic for health promotion with an uncertain scientific basis. In this study, B16-F10 tumor-bearing mice were established to evaluate the immunomodulatory activity of HLP. The proliferation and apoptosis assays were performed to evaluate the anti-inflammatory activity of honey bee larva extract (HLE) in RAW 264.7 macrophage. The in vivo experimental results demonstrated that the oral administration of freeze-dried HLP (4 and 6 g/kg) significantly enhanced the spleen index, the percentage of CD4+cells, and the ratio of CD4+ and CD8+ T lymphocytes (CD4+/CD8+) in the peripheral blood compared with those in the tumor control mice. The in vitro studies demonstrated the potent immunomodulatory activities of HLE through the induction of RAW 264.7 macrophage proliferation and the mitigation of doxorubicin (DOX)-induced toxicity. HLE also exhibits anti-inflammatory activity by decreasing the production of nitric oxide (NO) and the cytokine level of interleukin-6 (IL-6) in lipopolysaccharide (LPS)-stimulated RAW 264.7 macrophage. The present study provides important scientific evidence for the immunomodulatory and anti-inflammatory activities of HLP and HLE.
Introduction
Color images are available online
Malignant cancer is a leading cause of death worldwide. 1 Currently, advances in antitumor strategies and novel drug screening have been performed worldwide to improve longer survival in cancer patients. Among them, the therapeutic power of many useful natural products with a long history and affordable prices has attracted more and more research. In contrast, substantial evidence indicates that the immune system plays a critical role in controlling the development and progression of cancer, 2,3 thereby making it more and more important to find novel anticancer drugs that can modulate the host immune response and inflammatory reaction.
Honey bee larva was first recorded in the earliest extant classic on pharmacology for traditional Chinese medicine, “Shen nong ben cao jing.” 4 In China, honey bee larva has been used to treat and prevent various diseases for more than 1000 years. With the exception of one previous study, which reported that lyophilized powder of honey bee larvae could alleviate tinnitus-associated depression, 5 few studies on the bioactivities of honey bee larva have been reported. Previously, we found that the oral administration of lyophilized honey bee larva powder (HLP) could inhibit atopic dermatitis-like skin lesion development in NC/Nga mice by suppressing the responses of both T-helper (Th)1 and Th2 cells. 6
Our latest work indicated that the oral application of the freeze-dried HLP suppressed tumor growth and metastasis in tumor-bearing BALB/c mice. 7 In vivo results also suggested that HLP was safe under the tested dosage, and in vitro assay demonstrated that its extract, namely honey bee larva extract (HLE), showed no toxic effect neither on tumor cells nor on normal cells. It is well known that macrophages play an important role both in immune and inflammation processes, 8,9 and RAW 264.7 macrophage is widely used in the laboratory for the identification of the immunomodulatory and anti-inflammatory effects of numerous natural products. 10 –12 Therefore, in the present study, we evaluated the immunomodulatory activity of HLP in B16-F10 tumor-bearing mice and identified the immune-enhancement and anti-inflammatory activities of HLE in RAW 264.7 macrophages.
Materials and Methods
Preparation of HLP and HLE
The freeze-dried HLP (∼21 days old) was purchased from Nakahara Co., Ltd. (Saitama, Japan).
Sixty grams of HLP was extracted with 600 mL of 70% ethanol at room temperature for 24 h, and the extraction was repeated once. The two extractions were then combined, concentrated using a rotary vacuum evaporation, lyophilized, and labeled as HLE. The obtained HLE with yield of 39.45% (w/w) was stored at −20°C for further use.
Immunomodulatory activity of HLP in B16-F10 tumor-bearing mice
Cell culture
The murine melanoma cell line B16-F10 was purchased from the Riken Cell Bank (Tsukuba, Japan). The cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco BRL, Grand Island, NY, USA), supplemented with 10% (v/v) fetal bovine serum (FBS), and 1% (v/v) penicillin/streptomycin at 37°C in a humidified atmosphere with 5% CO2.
Animals
Six-week-old BALB/c mice (female) were purchased from Beijing HFK Bioscience Co., Ltd. (Beijing, China). Mice received human care and were acclimatized to the laboratory condition at a temperature of 23°C ± 1°C, relative humidity of 55% ± 5%, and an alternating 12-h light/12-h dark cycle, for 1 week before being used for the experiments. The mice were fed on standard food pellet and water ad libitum. This study was approved by Animal Experiments Committee of Chinese Academy of Medical Sciences with license number ILAS-GR16002.
Design of mouse experiment
A total of 20 mice were used for this study. B16-F10 cells (5 × 105/mouse) were injected into 16 mice through the tail vein. After 3 days, the mice were randomly allocated to four groups. These were the normal control (not inoculated with tumor cells, administrated with distilled water, n = 5) group, tumor control (inoculated with tumor cells, administrated with distilled water, n = 5) group, 4 g/kg/day HLP (inoculated with tumor cells, administrated with 4 g/kg/day of HLP, n = 5) group, and 6 g/kg/day HLP (inoculated with tumor cells, administrated with 6 g/kg/day of HLP, n = 5) group. The HLP was prepared with distilled water before use. The distilled water and HLP were administrated orally every day for 22 days, and the body weights of the mice were monitored on alternating days.
Before mice were sacrificed, the body weights of each mouse were measured, and the blood of each mouse was collected retro-orbitally for further analysis; afterward, their spleen weights were measured and then sampled for histopathology staining. The spleen index was calculated according to the following equation: spleen index = weight of the spleen (mg)/body weight (g). 13
Determination of the lymphocyte phenotype in the peripheral blood
Peripheral blood was collected retro-orbitally from each mouse and collected using ethylene diamine tetraacetic acid (EDTA K3) tubes for lymphocyte phenotype determination. The blood samples were lysed in RBC Lysis Buffer (eBioscience) and then stained with the following fluorescein-labeled monoclonal antibodies: FITC-conjugated anti-CD4 (Cat. No. 561835; BD Pharmingen), PE-conjugated anti-CD8 (Cat. No. 561095; BD Pharmingen), PerCP/Cy5.5-conjugated anti-CD45R/B220 (Cat. No. 103236; BioLegend), Alexa Fluor 647-conjugated anti-F4/80 (Cat. No. 123122; BioLegend), and PE/Cy7-conjugated anti-NK1.1 (Cat. No. 108713; BioLegend). Isotype-matched antibodies were used as negative controls.
Cells were analyzed using a Gallios flow cytometer (Beckman Coulter), and more than 50,000 events were analyzed using FlowJo software. The absolute specific cell populations were calculated by multiplying the respective percentages by the whole number of lymphocytes. The ratio of CD4+ and CD8+ T lymphocytes, an indicator of measurement to investigate the effect of HLP on T lymphocyte subpopulations, was determined as CD4+/CD8+ according to Bie et al. 14
Histopathology staining
The spleen organs were fixed with formaldehyde, embedded in paraffin, and cut into 3-mm thick sections. Representative sections were stained with hematoxylin and eosin (H&E) and examined by light microscopy as described previously. 13
Immunomodulation activity assessment in vitro
Cell line and proliferation assay
The murine macrophage cell line RAW 264.7 was used for in vitro assays. RAW 264.7 macrophage was purchased from the Riken Cell Bank and maintained in DMEM supplemented with 10% FBS and 1% (v/v) penicillin/streptomycin at 37°C in an atmosphere of 5% CO2 and 95% air in a humidified incubator. The proliferation effect of HLE treatment on RAW 264.7 macrophages was determined after 24/48 h of treatment by Cell Counting Kit-8 (CCK-8; Dojindo, Kumamoto, Japan) assay as described previously. 15
For morphologic observation, RAW 264.7 macrophages with a density of 2 × 105 cells/well were plated in a six-well plate and allowed adherence for 24 h. The culture medium was then replaced with DMEM supplemented with 0, 62.5, 125, 250, 500, or 1000 μg/mL of HLE. After 24 h of incubation at 37°C, the cell morphologies were observed using a phase-contrast inverted microscope fitted with a digital camera (Axio Vert A1; Carl Zeiss, Germany).
Protection activity of HLE against doxorubicin-induced toxicity
RAW 264.7 macrophages with a density of 3 × 103 cells/well were plated in a 96-well plate. After 24 h of incubation at 37°C, cells were treated with DMEM supplemented with 0.05 or 0.1 μmol/L doxorubicin (DOX) (final concentration) in the presence of HLE (0, 62.5, 125, 250, 500, and 1000 μg/mL). After 24 h of incubation at 37°C, the cells were subjected to viability measurement using the CCK-8 method as described above. Morphological observation following the DOX (0.1 μmol/L) treatment was also carried out accordingly.
Protective activity of HLE against DOX-induced apoptosis
To measure apoptosis, RAW 264.7 macrophages with a density of 2 × 105 cells/well were seeded in six-well plates; after 24 h of culture, the cells were treated with 0, 500, and 1000 μg/mL of HLE. Apoptosis analysis was conducted using Guava Nexin Reagents (Millipore, Billerica, MA, USA) and analyzed by a Guava PCA flow cytometer (Millipore) as previously described. 16 The data were further analyzed using FlowJo Software (version 7.6; FlowJo, LLC, USA).
The protein expression level of anti-pro caspase 3 (Cell Signaling Technology) was determined by western blotting 17 with anti-beta actin (Abcam, Cambridge, MA, USA) being used as an internal loading control.
Effect of HLE in alleviating H2O2-induced oxidative stress
RAW 264.7 macrophages with a density of 3 × 103 cells/well were plated in a 96-well plate and allowed to adhere at 37°C for 24 h, followed by 300 or 600 μmol/L H2O2 (final concentration) treatment in the presence of HLE (0, 62.5, 250, 500, and 1000 μg/L) for 24 h. Cell proliferation assay was then performed according to the CCK-8 method as described above.
Anti-inflammatory activities of HLE against lipopolysaccharide-induced inflammation
Cell viability assay
RAW 264.7 macrophages with a density of 3 × 103 cells/well were plated in a 96-well plate. After 24 h of incubation at 37°C, the RAW 264.7 macrophages were treated with 500 ng/mL of lipopolysaccharide (LPS) (final concentration) in the presence of HLE (0, 15.6, 31.3, 62.5, 125 μg/mL) for 24 h. Cell proliferation assay was then performed according to the CCK-8 method as described above.
Measurement of interleukin-6 and nitric oxide production
To assess anti-inflammatory activity, 2 × 105 macrophage cells/well were seeded in six-well plates, cultured at 37°C for 24 h, and then treated with 500 ng/mL of LPS (final concentration) in the presence of HLE (0, 15.6, 15.6, 31.3, 62.5, and 125 μg/mL). The interleukin-6 (IL-6) level in the supernatant was determined using the Mouse IL-6 DuoSet ELISA Kit (R&D systems, USA).
Nitrite accumulation was identified using the Griess reagent, which was used as an indicator of nitric oxide (NO) production in the medium. 15 NaNO2 was used as the standard to calculate the nitrite concentration.
Statistical analyses
All experiments were carried out more than thrice, and data are expressed as the mean ± standard deviation. One-way analysis of variance followed by a Duncan's test was carried out on the experimental data. Statistical significance was defined as significant (*P value <.05), very significant (**P value <.01), and very very significant (***P value <.001). All the statistical analyses were performed using SPSS 13.0 software (SPSS, Inc., Chicago, USA).
Results
HLP enhances CD4+ cells in the peripheral blood and increases the spleen index in vivo
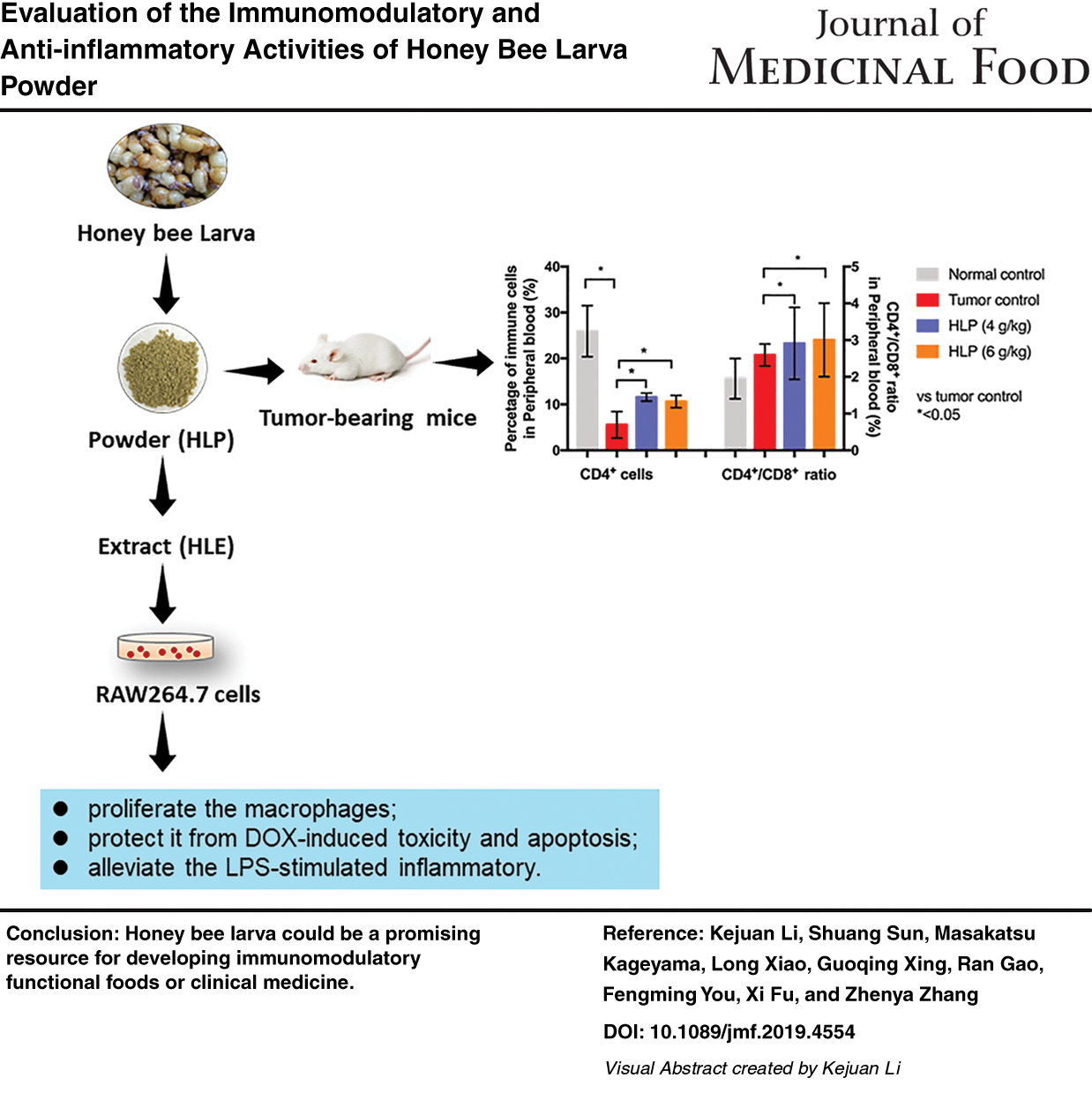
As shown in Figure 1, among all the peripheral blood immune cells, the percentage of CD4+ was sharply decreased in mice with lung metastasis compared with normal mice. It is well known that CD4+ are the T helper (Th) lymphocytes and CD8+ are the T cytotoxic (Tc) lymphocytes. In this study, the percentage of CD4+ cells was 25.94% ± 5.57% in the peripheral blood of normal control mice, and it decreased to 5.55% ± 2.88% in the tumor control group. The oral application of HLP with 4 and 6 g/kg dosage significantly (P < .05) increased the percentage of CD4+ cells to 11.58% ± 0.86% and 10.61% ± 1.34%, respectively.
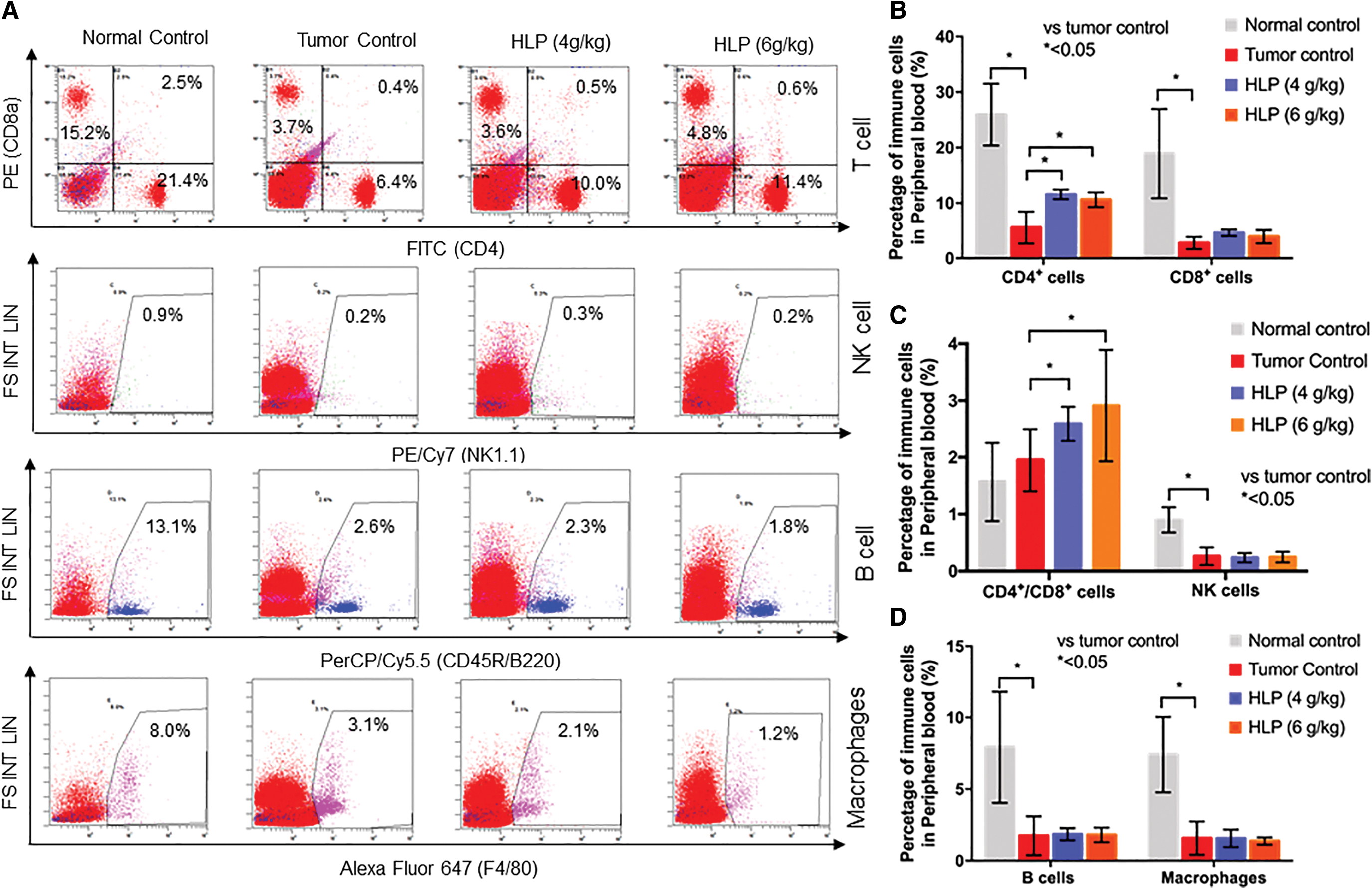
Flow cytometric analysis of cell markers for T cells, NK cells, B cells, and macrophages
Moreover, the CD4+/CD8+ ratio in the peripheral blood of HLP-treated mice was significantly (P < .05) higher than that in the tumor control group. The other immune cells, including CD8+ cells, natural killer (NK) cells, B cells, and macrophages, exhibited few significant differences between the HLP treatment groups and the tumor control group.
As shown in Figure 2A, after consecutive treatment for 22 days, with a higher spleen index of 4.67 ± 0.57 mg/g and 4.47 ± 0.53 mg/g in 4 and 6 g/kg HLP treated groups, HLP was found to significantly (P < .05) elevate the spleen organ growth in B16-F10 tumor bearing mice (Tumor control group, 3.89 ± 0.17 mg/g). Meanwhile, the H&E stained histopathological images of the spleen illustrated that HLP had no damage on the spleen (Fig. 2B).

Effects of HLP on spleen index and histological analysis on the spleen tissues. Effects of HLP on the spleen index
HLE affects macrophage proliferation
To verify the immunomodulatory activities of honey bee larva, RAW 264.7 macrophages were used to evaluate the proliferative effect of HLE on immune cells in vitro. As shown in Figure 3A, the cell viability assay indicated that exposure to HLE stimulated macrophage proliferation after 24 h of culture. At a concentration of 125 μg/mL, HLE increased the macrophage viability rate to 122.68% ± 10.87%, and the cell viability rate reached 167.38% ± 9.46% at the HLE concentration of 1000 μg/mL. HLE treatment for an additional 24 h did not result in a significant increase in RAW 264.7 proliferation (Fig. 3B). The morphological features shown in Figure 3C were in accordance with the above results. The proliferative effect of HLE on RAW 264.7 macrophages indicated the potential of immunomodulatory activity of HLE in vitro.

Proliferation effect of HLE in RAW 264.7 cells. The proliferation effect of HLE (62.5 ∼ 1000 μg/mL) on RAW 264.7 cells for 24 h
HLE protects against DOX-induced toxicity and apoptosis in RAW 264.7 macrophages
According to the results of Figure 4A and B, the viability of macrophage cells treated with 0.05 and 0.1 μmol/L DOX was found decreased to 57.12% ± 8.80% and 38.81% ± 14.16%, respectively. However, after being cocultured with HLE, the viability of RAW 264.7 macrophages was noticeably increased in a dose-dependent manner, and the cell viabilities reached 115.83% ± 6.82% and 106.85% ± 8.36% at the HLE concentration of 1000 μg/mL, indicating that HLE has potential protective activity in RAW 264.7 macrophage. In addition, the above proliferative activities of HLE were reflected in the morphological changes (Fig. 4C).

Protection activity of HLE on DOX-induced toxicity. RAW 264.7 cells were treated with 0.05 μmol/L of DOX
The protective effect of HLE in DOX-treated macrophages was confirmed by the flow cytometric analysis results. As shown in Figure 5A and B, the apoptotic population of RAW 264.7 macrophages was 57.9% after 0.1 μmol/L of DOX treatment for 24 h. The apoptotic population of cells treated with 500 and 1000 μg/mL HLP was reduced to 39.9% and 24.64%, respectively.
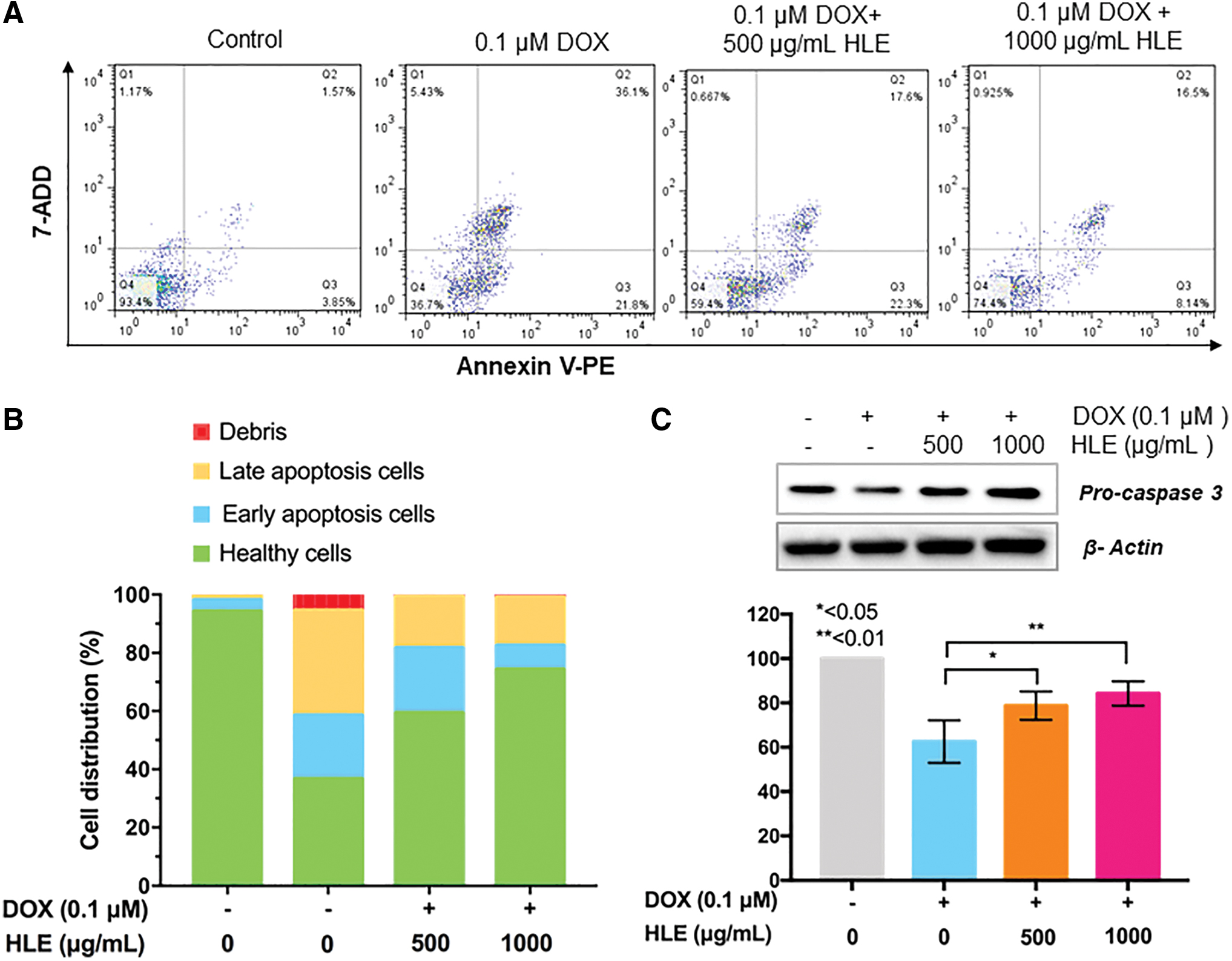
Protection activity of HLE on DOX-induced apoptosis. Control, 0.1 μmol/L DOX, and 0.1 μmol/L DOX + HLE treated cell population were examined for Annexin-V expression by cytometric analysis
As shown in Figure 5C, western blotting also confirmed the protective effect of HLE against DOX-induced apoptosis. The results above indicated that HLE exhibits promising protective effects against DOX-induced toxicity and apoptosis in macrophages, implying that it may serve as a potent complementary reagent during the use of chemotherapeutics (i.e., DOX).
HLE alleviates H2O2-induced oxidative stress in RAW 264.7 macrophages
To identify the protective effect of HLE against H2O2-induced oxidative stress in RAW 264.7 macrophages, the viability of the macrophages treated with HLE at different doses (0, 62.5, 125, 250, 500, and 1000 μg/mL) was evaluated. As shown in Figure 6A and B, treatment with 300 and 600 μmol/L H2O2 induced cytotoxicity in RAW 264.7 macrophages as indicated by decreased viability to 32.72% ± 4.34% and 24.95% ± 3.33%, respectively. In addition, compared to the model group, 250, 500, and 1000 μg/mL HLE significantly attenuated H2O2-induced cytotoxicity, which represented a dose-dependent manner (P < .05). Thus, these data suggested that HLE could decrease the oxidative stress in RAW 264.7 macrophages induced by H2O2.

Protection activity of HLE on H2O2-induced stress. RAW 264.7 cells were treated with 300 μmol/L of H2O2
Anti-inflammatory activities of HLE
To explore the anti-inflammatory activities of HLE in vitro, we investigated the proliferative effect of HLE (0–1000 μg/mL) on LPS (500 ng/mL)-stimulated RAW 264.7 macrophage. As shown in Figure 7A, LPS treatment showed no significant difference in cell viability compared with nontreated cells, and HLE did not have proliferative effect on RAW 264.7 macrophages at concentrations up to 125 μg/mL.

Anti-inflammatory activities of HLE in LPS-induced inflammation. HLE did not show any proliferation effect on macrophages RAW264.7 within the concentration of 125 μg/mL
We next examined the NO production and IL-6 expression in LPS-stimulated RAW 264.7 macrophages after 15.6, 31.3, 62.5, and 125 μg/mL of HLE treatment. As shown in Figure 7B, a three-fold increase in NO production was observed in the cells treated with 500 ng/mL LPS compared with that observed in the blank control. However, treatment with 62.5 and 125 μg/mL HLE efficiently reduced the NO production with an inhibition rate of 14.44% and 18.47%, respectively. Moreover, HLE treatment (62.5 and 125 μg/mL) reduced the cytokine IL-6 expression with inhibition rates of 20.7% and 36.29%, respectively (Fig. 7C). In this study, HLE significantly alleviated the production of NO and the expression of cytokine IL-6 in LPS-induced macrophages, suggesting that HLE has potent anti-inflammatory activities.
Discussion
Our previous research used tumor xenograft assays in BALB/c mice in vivo, and the results revealed that application of freeze-dried HLP orally suppresses both tumor growth and lung metastasis. However, our previous result also suggested that HLE shows nontoxic effect in cancer cells at concentration up to 1000 μg/mL in vitro. 7 Many chemotherapeutic agents exert their antitumor effects through antiproliferative and cytotoxic actions, and these anticancer drugs may also modulate the host immune response and inflammatory reaction. 18 –20 The significant tumor suppression effect of HLP promoted us to examine whether the administration of HLP changed the composition of immune cells in the host.
In the present work, we compared the composition of four immune cells in the peripheral blood, including T cells, B cells, NK cells, and macrophages. The flow cytometric results suggested that the oral administration of HLP (4 and 6 g/kg) significantly elevates peripheral blood CD4+ cells and enhances the CD4+/CD8+ ratio compared with those in the peripheral blood from mice in the tumor control group (P < .05). CD4+ T cells play significant role in modulating immune responses to pathogens and tumor cells. 21 The dominant effector cells mediating tumor killing are CD8+ cytotoxic T lymphocyte (CTL) cells; CD4+ T cells are not directly involved in antitumor immunity; rather, CD4+ T cells function to prime CD8+ CTL cells by activating antigen-presenting cells (APCs), increasing antigen presentation on MHC class I and secreting excitatory cytokines, such as IL-2. 22 –24 Moreover, a previous study also reported that a high CD4+/CD8+ ratio favors tumor suppression. 25
According to Figure 2A, the spleen index in the HLP treated mice increased significantly compared to that in the tumor control group, indicating an elevated immune response and spleen organ growth. Meanwhile, the H&E staining results indicated that HLP had no toxic effect to the spleen (Fig. 2B). Previously, we evaluated the direct anti-inflammatory effects of the freeze-dried HLP on an atopic dermatitis model.l6 Results found that the NC/Nga mice administrated with the freeze-dried HLP-supplemented diet showed histological alleviation effect on the ear swelling and the infiltration of inflammatory cells in the skin. The present study supports the assertion that the tumor suppression effect of HLP on tumor-bearing mice may be attributable, at least partially, to the immune-enhancement activity of HLP in vivo.
It is well known that the human host defense is complex and contains multiple levels, involving many cell types with distinct but overlapping roles. 26 We next examined the immunomodulatory activities of HLE using RAW 264.7 macrophages in vitro. Figure 3A and C shows that 250, 500, and 1000 μg/mL HLE had a dose-dependent stimulatory effect on macrophage cell viability after 24 h of culture. The proliferative effect of HLE on RAW 264.7 macrophages indicated the potential of immunomodulatory activity of HLE in vitro.
As a broad-spectrum antitumor anthracycline antibiotic, DOX is widely used to treat various types of cancers. Figure 4B and C shows obvious cytotoxic effects of DOX at concentrations of 0.1 μmol/L DOX with a viability of 38.81% ± 14.16%. However, after coculturing with HLE, the viability of RAW 264.7 macrophages increased with the increasing of HLE concentrations. The macrophage viability was then reached to 106.85% ± 8.36% at the HLE concentration of 1000 μg/mL.
To confirm the protective effect of HLE on DOX-induced macrophages, flow cytometric analysis was investigated to evaluate apoptotic populations in untreated, 0.1 μmol/L DOX treated, and 0.1 μmol/L DOX+HLE-treated cells. Figure 5A and B shows that after treatment with 0.1 μmol/L DOX for 24 h, the apoptotic population of RAW 264.7 macrophages was 57.9%. In addition, the apoptosis population in the presence of 500 and 1000 μg/mL HLE was reduced to 39.9% and 24.64%, respectively. Caspase 3 is activated in apoptotic cells both by extrinsic and intrinsic pathways. 27,28 In this study, the expression of pro-caspase 3 in DOX-treated macrophages was increased in the presence of 500 and 1000 μg/mL HLE (Fig. 5C).
Thus, these results indicated that HLE exhibits promising protective effects against DOX-induced toxicity and apoptosis in macrophages, implying that HLE may serve as a potent complementary reagent during the use of chemotherapeutics (i.e., DOX).
Oxidative stress is a major effect of DOX-induced cytotoxicity, and it plays a critical role in various diseases, including cancer. 29,30 To determine the protective effect of HLE against H2O2-induced oxidative stress, we examined the viability of RAW 264.7 macrophage after different concentrations (62.5, 125, 250, 500, and 1000 μg/mL) of HLE treatment. Figure 6 shows a decreased viability of 32.72% ± 4.34% and 24.95% ± 3.33% after treatment with 300 and 600 μmol/L H2O2. However, treatment with 250, 500, and 1000 μg/mL of HLE significantly attenuated the damage in a dose-dependent manner (P < .05). Thus, these findings suggested that HLE shows protective effects against H2O2-induced oxidative stress.
The immunomodulatory effect of HLP and HLE provides the therapeutic basis for the treatment of autoimmune diseases. The inflammatory response is a complex physiological response of living organisms against harmful stimuli, such as toxic molecules. 31 Immune cells travel to the sites of injury or infection and cause inflammation. It is well known that macrophage cells perform a significant role in the inflammatory process, and LPS-stimulated RAW 264.7 macrophages are commonly used for anti-inflammatory activity investigation of natural products. 8,10
To identify the anti-inflammatory activities of HLE in vitro, the effect of HLE (0, 15.6, 31.3, 62.5, and 125 μg/mL) on LPS (500 ng/mL)-stimulated RAW 264.7 macrophage proliferation was evaluated. Figure 7A shows that HLE did not affect RAW 264.7 macrophage proliferation at a dose up to 125 μg/mL. Thus, we examined the NO production and cytokine IL-6 expression in LPS-induced RAW 264.7 macrophage after treatment with HLE at a dose up to 125 μg/mL. Figure 7B shows a three-fold increase in NO production after treatment with 500 ng/mL LPS compared with that in control cells. However, treatment with 62.5 and 125 μg/mL HLE significantly reduced NO production with inhibition rates of 14.44% and 18.47%, respectively. Moreover, treatment with 62.5 and 125 μg/mL HLE significantly reduced IL-6 production with inhibition rates of 20.7% and 36.29%, respectively (Fig. 7C).
It has been reported that LPS could activate the mitogen-activated protein kinase (MAPK), phosphatidylinositol 3-kinase (PI3K)/AKT, and NF-κB signal pathways, upregulate the expression of cytokines TNF-α, IL-1β, and IL-6, and increase the production of inducible NO synthase and NO. 32,33 In this study, HLE significantly alleviated the NO and IL-6 production in LPS-stimulated macrophages, suggesting that HLE has potent anti-inflammatory activities.
In conclusion, the present work suggests that the oral application of freeze-dried HLP significantly enhances the spleen index, the CD4+ cells, and the CD4+/CD8+ ratio in peripheral blood compared to the tumor control mice. The in vitro studies also indicated that the ethanol extract of HLP exhibits potent immunomodulatory activities by enhancing the cell viability of RAW 264.7 macrophages and protecting them from DOX-induced toxicity and apoptosis. Furthermore, HLE significantly alleviates the LPS-stimulated inflammation in RAW 264.7 macrophages. The experimental data indicated that honey bee larva could be a promising resource for developing immunomodulatory functional foods or clinical medicine.
Footnotes
Authors' Contributions
K.L., S.S., M.K., L.X., and X.F. conducted the experiments and analyzed the data. G.X., R.G., F.Y., and Z.Z. contributed reagents/materials/analysis tools. K.L. and Z.Z. conceived the study and wrote the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by National Natural Science Foundation of China (81904081), Sichuan Science and Technology Department (2019IDRC0124), Chengdu University of Traditional Chinese Medicine (BSH2018005), and Chinese Academy of Medical Sciences (2016-I2M-3-019).