
Abstract
Aging can cause retinal degeneration, which leads to visual impairment among the elderly population. Age-dependent increases in amyloid beta (Aβ) inducesinflammatory cytokine overexpression in the retinal pigment epithelium (RPE), which promotes the progression of age-related retinal degeneration. However, whether dietary antioxidants are useful for the treatment of RPE degeneration remains to be clarified. This study exposited the protective activities and underlying mechanisms of grape seed extracts (GSEs) against Aβ-induced proinflammatory events in mouse retinas and ARPE-19 cells. The experimental data demonstrated that GSEs attenuated the increases in messenger RNA (mRNA) levels of interleukin 12 (IL-12), interleukin 6 (IL-6), interleukin 1β (IL-1β), and interleukin 18 (IL-18) in the retinal tissues of Aβ-treated mice. The experimental results in mice were confirmed by findings in ARPE-19 cells with or without treatment with GSEs. GSEs affected the protein expression levels of endoplasmic reticulum stress markers in ARPE-19 cells exposed to Aβ. Knockdown of Bip blocked the inhibitory activities of GSEs on mRNA levels of IL-6, IL-1β, IL-18, and IL-8. We conclude that GSEs may suppress proinflammatory cytokines partly by increasing the expression of Bip.
Introduction
Age-related retinal degeneration is a major cause of visual damage among the aged population. 1,2 Chronic inflammation is the leading cause of retinal degeneration. 3 Age-dependent increases in amyloid beta (Aβ) are present in retinal pigment epithelial (RPE) cells, 4 and the content of Aβ is correlated with proinflammatory events in the retina. 5,6 Aβ accumulation is involved in multiple age-related degenerative illnesses. Compared with other forms of Aβ, Aβ1–40 is the most prevalent type in eye diseases. 7 It has been reported that Aβ1–40 deposition stimulates the upregulation of interleukin 12 (IL-12), interleukin 6 (IL-6), interleukin 1β (IL-1β), interleukin 18 (IL-18), and interleukin 8 (IL-8) genes in the RPE cells. 8 Studies have shown a significant correlation between inflammatory factors and age-related retinal degeneration. 9
Moreover, aging may trigger retinal degeneration through oxidative stress. Previous research showed that oxidative stress has a critical role in upregulation of proinflammatory cytokines in RPE cells. 10 Therefore, dietary antioxidants may decrease age-related RPE injury and provide a new preventive or therapeutic strategy for treating RPE degeneration. Grape seed extracts (GSEs) are rich in proanthocyanidins and have strong antioxidant activity. 11 Our previous experiments have also indicated that GSEs attenuate oxidative stress and increase in proinflammatory cytokines in the retina of streptozotocin-induced diabetic rats. Thus, we hypothesized that GSEs might be used as dietary supplements to prevent RPE degeneration. However, the underlying molecular mechanisms of the activities of GSEs are not clear.
In this study, we investigated whether GSEs can suppress age-related increases of proinflammatory factors and injury in RPE cells using a mouse model and ARPE-19 cells. For this purpose, GSEs were administered to mice with intravitreal injection of Aβ1–40 and then the expression of inflammatory cytokines was detected. The ARPE-19 cells were also exposed to Aβ1–40 to determine the sheltering effects of treatment with GSEs on RPE and to explore their underlying mechanisms.
Materials and Methods
Materials
GSEs from Vitis vinifera L. were provided by JF-NATURAL Corporation (Tianjin, China). A detailed description of the extract preparation was described in our previous study. 12 ARPE-19 (ATCC CRL-2302) was obtained from the American Type Culture Collection (Manassas, VA, USA).
Amyloid oligomerization
Aβ1–40 peptides were dissolved in deionized water to a concentration of 28 μg/μL. Aβ solutions were diluted by addition of phosphate buffered saline (PBS), the final concentration was 2.8 μg/μL, and then incubated for 4 days at 37°C. The aggregation states of Aβ1–40 were examined using electron microscopy (Supplementary Fig. S1).
Animal model and treatment
Two-month-old male C57BL/6 mice were obtained from the Animal Center of the Chinese Academy of Sciences (Shanghai, China). Mice were randomized into the following four groups: control, Aβ1–40, Aβ1–40 + 50 mg/kg, and Aβ1–40 + 100 mg/kg. The “Aβ1–40 + 50 mg/kg” and “Aβ1–40 + 100 mg/kg” groups were treated with 50 and 100 mg/kg GSEs by oral gavage once daily for 4 weeks, respectively. The GSEs were dissolved in distilled water. Then, the “Aβ1–40,” “Aβ1–40 + 50 mg/kg,” and “Aβ1–40 + 100 mg/kg” groups were anesthetized with 1.5% sodium pentobarbital (100 μL/20 g intraperitoneally) and were administered an intravitreal injection of oligomeric Aβ1–40 (14 μg/5 μL) in PBS. The oligomeric Aβ1–40 peptides were delivered using a 32-gauge Hamilton needle and syringe under a dissecting microscope (SZ2-lLST; Olympus, Tokyo, Japan). The eyes of each mouse were enucleated under anesthesia at 4-day postinjection (Project permission number: A2015024).
Retinal hematoxylin and eosin assays and TdT-mediated dUTP nick-end labeling detection
Hematoxylin and eosin (H&E) staining was prepared as previously described, 13 and the images were captured using a microscope (Zeiss, Jena, Germany).
The paraffin-embedded sections were stained with TdT-mediated dUTP nick-end labeling (TUNEL) fluorescent dye according to an In Situ Cell Death Detection Kit (Roche, Shanghai, China). The nuclei were counterstained with 4′,6-diamidino-2-phenylindole. The morphology of retinal sections was detected using a Leica DM 3000 (Leica, Germany) fluorescent microscope and analyzed through Image J software.
Cell culture and treatment
ARPE-19 cells were cultured and treated as previously described. 4 In brief, cells were treated with GSEs (5 μg/mL or 10 μg/mL) or vehicle (0.0001% dimethylsulfoxide) for 24 h. Then, oligomeric Aβ1–40 peptides were added to culture media to reach a final concentration of 5 μM, and then incubated for 24 h. Cells that had neither been exposed to Aβ1–40 nor treated with GSEs were regarded as controls.
Quantitative real-time PCR
Quantitative real-time PCR (qRT-PCR) was conducted according to our former published studies. 12 The sense and antisense primer sequences were as follows: mouse GAPDH, 5′-CGGAGTCAACGGATTTGGTCGTAT-3′ and 5′-AGCCTTCTCCATGGTGGTGAAGAC-3′; mouse IL-12, 5′-ATGGAGTCATAGGCTCTGGAAA-3′ and 5′-CCGGAGTAATTTGGTGCTTCAC-3′; mouse IL-6, 5′-CTTCCATCCAGTTGCCTTCTTG-3′ and 5′-AATTAAGCCTCCGACTTGTGAAG-3′; mouse IL-1β, 5′-AGTTGACGGACCCCAAAAGAT-3′ and 5′-GTTGATGTGCTGCTGCGAGA-3′; mouse IL-18, 5′-CAGGCCTGACATCTTCTGCAA-3′ and 5′-CTGACATGGCAGCCATTGT-3′; human GAPDH, 5′-TGTAGACCATGTAGTTGAGGTCA-3′ and 5′-AGGTCGGTGTGAACGGATTTG-3′; human IL-12, 5′-ACCTGACCCACCCAAGAACT-3′ and 5′-GGACCT GAACGCAGAATGTC-3′; human IL-6, 5′-GATGGCTGAAAAAGATGGATGC-3′ and 5′-TGGTTGGGTCAGGGGTGGTT-3′; human IL-1β, 5′-TTACAGTGGCAATGAGGATGAC-3′ and 5′-TGTAGTGGTGGTCGGAGATTC-3′; human IL-18, 5′-AGTCAGCAAGGAATTGTCTCC-3′ and 5′-GAAGCGATCTGGAAGGTCTG-3′; and human IL-8, 5′-ATGACTTCCAAGCTGGCCGT-3′ and 5′-TCCTTGGCAAAACTGCACCT-3′.
Western blotting
RPE–choroid tissues or ARPE-19 cells were prepared in lysis buffer and Western blotting analysis was conducted as described in previously published studies. 8 In brief, polyvinylidene difluoride membrane was exposed to antibodies against XBP-1 (1:1000; Cell Signaling Technology [CST], Beverly, MA, USA), p-IRE1 (1:2000; Abcam, Cambridge, MA, USA), ATF6 (1:2000; CST), Bip (1:2000; Abcam), NLRP3 (1:1000; CST), Caspase-1 (1:1000; CST), IL-1β (1:1000; CST), GAPDH (1:1000; CST), or β-actin (1:1000; CST) overnight. The membranes were then incubated with horseradish peroxidase-conjugated secondary antibodies (1:2000; Proteintech, Chicago, IL, USA). Binding was visualized using a Bio-Rad Clarity Western ECL (Hercules, CA, USA). The optical density of bands was evaluated using a ChemiDoc™ XRS + System (Hercules, CA, USA).
RNA interference
ARPE-19 cells were transfected with small interfering RNA (siRNA) targeting human Bip or nontargeting control siRNA using Lipofectamine RNAi MAX (Invitrogen, Carlsbad, CA, USA). The double-stranded siRNA was obtained from Invitrogen. The sense and antisense sequences were as follows: 5′-GACCCUGACUCGGGCCAAAUUUGAA-3′ (sense) and 5′-UUCAAAUUUG GCCCGAGUCAGGGUC-3′ (antisense).
Statistical analysis
Each experiment was repeated in triplicate. Results are presented as means ± standard deviations. Statistical significance was analyzed using Tukey post hoc test. A P value <.05 was considered significant.
Results
GSE attenuation of Aβ1–40-induced RPE impairment and suppression of proinflammatory cytokines in vivo
Retinal sections stained with H&E showed that the RPE cells of the control group exhibited normal morphology, whereas the RPE cells in the Aβ1–40 group exhibited morphometric changes. Treatment with 50 or 100 mg/(kg·d) of GSEs ameliorated the Aβ1–40-induced damage in the RPE layer (Fig. 1A). The effect of GSEs on Aβ1–40-induced retinal apoptosis was further explored. Immunofluorescent assays revealed TUNEL-positive staining in the outer nuclear layer (ONL). Apoptosis was found in the ONL only in the Aβ1–40 group compared with that of the control group at 4 days post-Aβ injection (indicated by white arrows in Fig. 1B). GSE significantly attenuated retinal apoptosis, demonstrating a protective effect of GSEs against Aβ-induced retinal degeneration.

The inhibitory effect of GSEs against Aβ1–40-induced RPE impairment and upregulation of proinflammatory cytokines in vivo. C57BL/6 mice were administered with or without GSEs per day through oral gavage for 4 weeks, and they were then injected intravitreally with 5 μL of 2.8 μg/μL of Aβ1–40. Aβ1–40-induced alterations in retinas were detected at day 4. Mice that were intravitreally injected with PBS and administered without GSEs were regarded as controls.
Next, RPE–choroid complexes were isolated and cytokine messenger RNA (mRNA) expression was measured by qRT-PCR after the injection of Aβ1–40. As shown in Figure 1C, the mRNA levels of IL-12, IL-18, IL-6, and IL-1β were increased in the RPE–choroid of Aβ1–40-injected mice compared with those of control mice. Treatment with 50 or 100 mg/(kg·d) GSEs decreased IL-12 mRNA by 33% or 37%, IL-6 mRNA by 26% or 35%, IL-1β mRNA by 17% or 13%, and IL-18 mRNA by 27% or 34% in the RPE–choroid of Aβ1–40-injected mice compared with those of mice without GSE treatment.
GSEs prevent Aβ1–40-induced increases in the expression of proinflammatory cytokines in ARPE-19 cells
To further investigate the protective effect of GSEs on RPE cells, the mRNA levels of IL-12, IL-6, IL-1β, IL-18, and IL-8 in ARPE-19, a human retinal epithelial cell line, were studied. Cells were pretreated with 5 μg/mL or 10 μg/mL GSEs for 24 h before the addition of Aβ1–40. The GSEs treatment dosage was not cytotoxic to cells (Supplementary Fig. S2). The mRNA levels of IL-12, IL-6, IL-1β, IL-18, and IL-8 were markedly increased by Aβ1–40. Treatment with 5 or 10 μg/mL GSEs reduced IL-12 mRNA by 63% or 75%, IL-6 mRNA by 25% or 50%, IL-1β mRNA by 60% or 57%, IL-18 mRNA by 54% or 50%, and IL-8 mRNA by 28% or 34% in Aβ1–40-induced ARPE-19 cells compared with those of vehicle-treated counterparts (Fig. 2), suggesting that GSEs attenuated the upregulation of inflammatory cytokines in RPE cells induced by Aβ1–40.
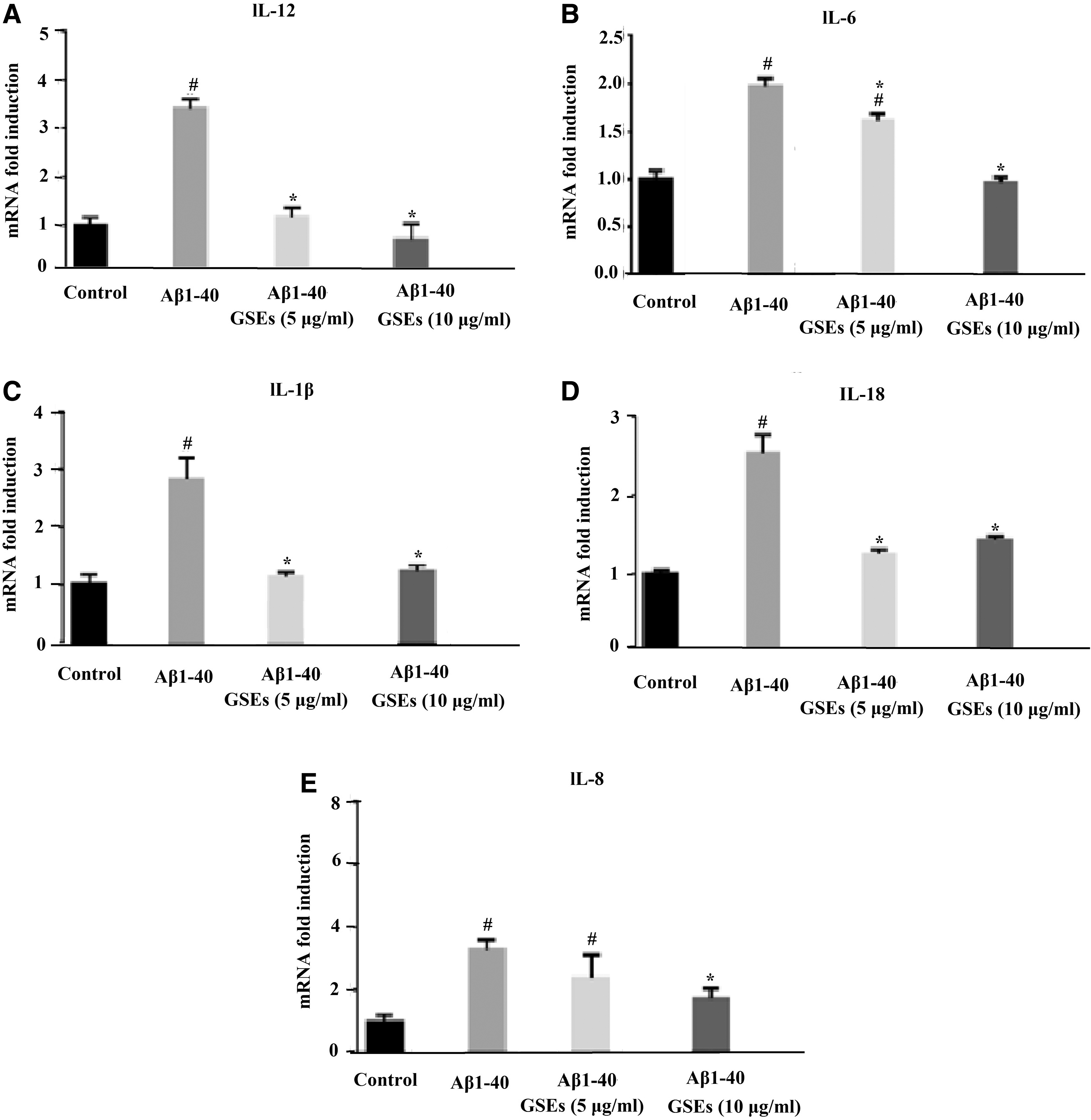
The inhibitory effect of GSEs against Aβ1–40-induced IL-12
Moreover, whether Aβ1–40 induces NLRP3 inflammasome activation in lipopolysaccharide (LPS)-primed ARPE-19 cells with or without GSEs treatment was tested using Western blotting, which confirmed the inhibition of inflammatory cytokines by GSEs at the protein level. Aβ-induced activation of NLRP3 is closely related to the chronic inflammatory process in age-related retinal degeneration. It cleaves caspase-1 precursor to produce active caspase-1. Caspase-1 cleaves the proform of IL-1β and IL-18 into mature cytokines IL-1β and IL-18. Our results demonstrated that Aβ-induced increases in the level of NLRP3, caspase-1, and IL-1β proteins in LPS-primed ARPE-19 cells were reduced by 10 μg/mL GSEs treatment (Fig. 3).
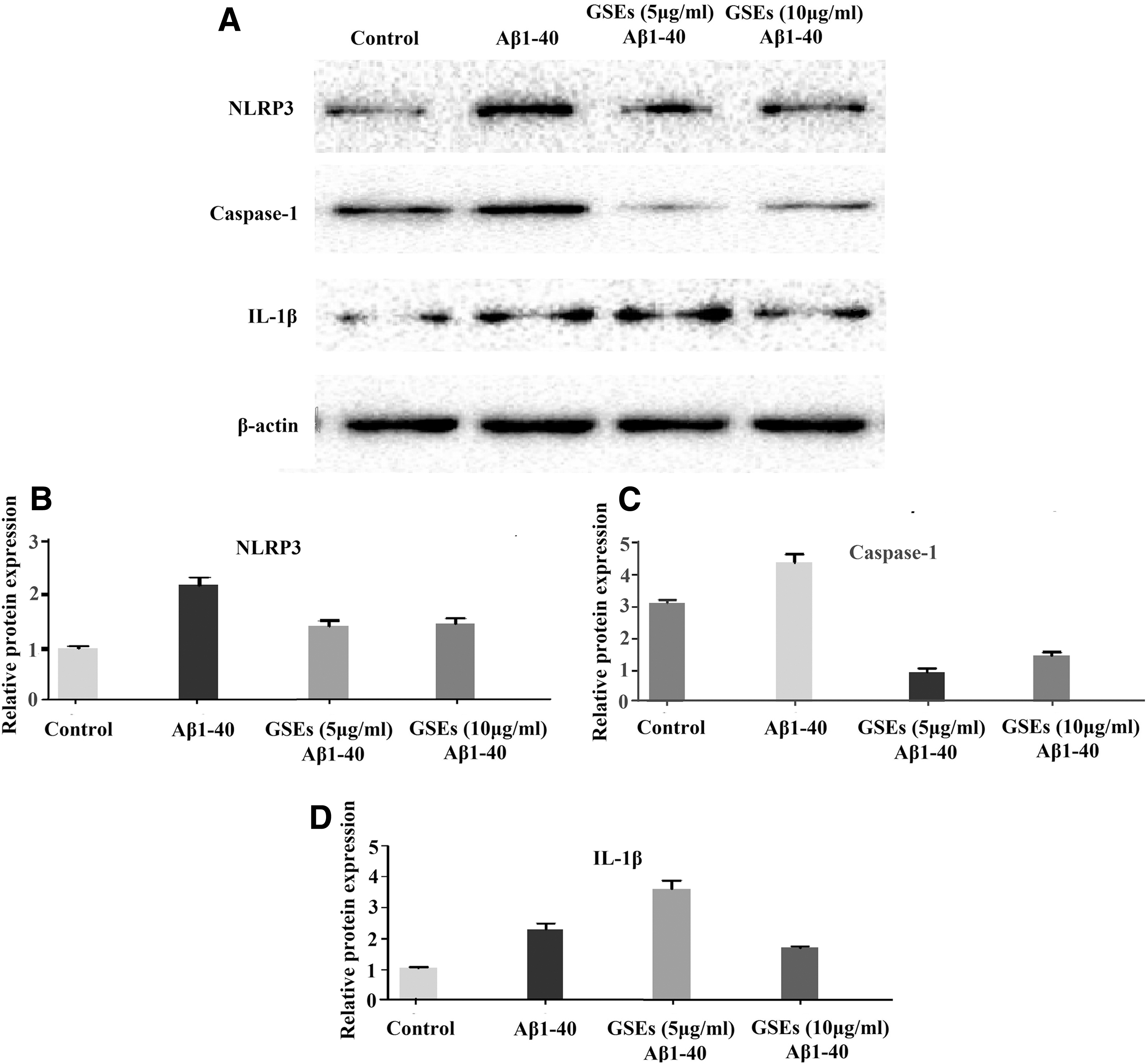
The inhibitory effect of GSEs against Aβ1–40-induced NLRP3 inflammasome activation in LPS-primed ARPE-19 cells. Cells were treated with LPS (10 μg/mL) for 3 h, then pretreated with or without GSEs for 24 h before the addition of Aβ1–40 (5 μM), and samples were collected after 12 h. The expression levels of NLRP3 inflammasome-related proteins were determined by Western blot analysis (
GSEs affect protein expression related to endoplasmic reticulum stress induced by Aβ1–40 in ARPE-19 cells
The endoplasmic reticulum (ER) stress markers—X-box binding protein 1 (XBP-1s), inositol-requiring enzyme 1a (p-IRE1 a), activating transcription factor 6 (ATF6), and 78 kD glucose-regulated protein (GRP78/Bip)—were significantly increased in Aβ1–40-loaded cells, as demonstrated by Western blotting. The effects of GSEs on XBP-1s, p-IRE1 a, ATF6, and Bip were analyzed. Figure 4 shows that treatment with 5 or 10 μg/mL of GSEs reduced XBP-1s protein by 30% or 29%, p-IRE1a protein by 36% or 47%, and ATF6 protein by 20% or 25% in Aβ1–40-loaded cells compared with those of vehicle-treated counterparts. Interestingly, treatment with GSEs resulted in increased GRP78 expression in Aβ1–40-loaded cells, indicating an important role of GRP78 in GSE protecting against Aβ-induced damage.
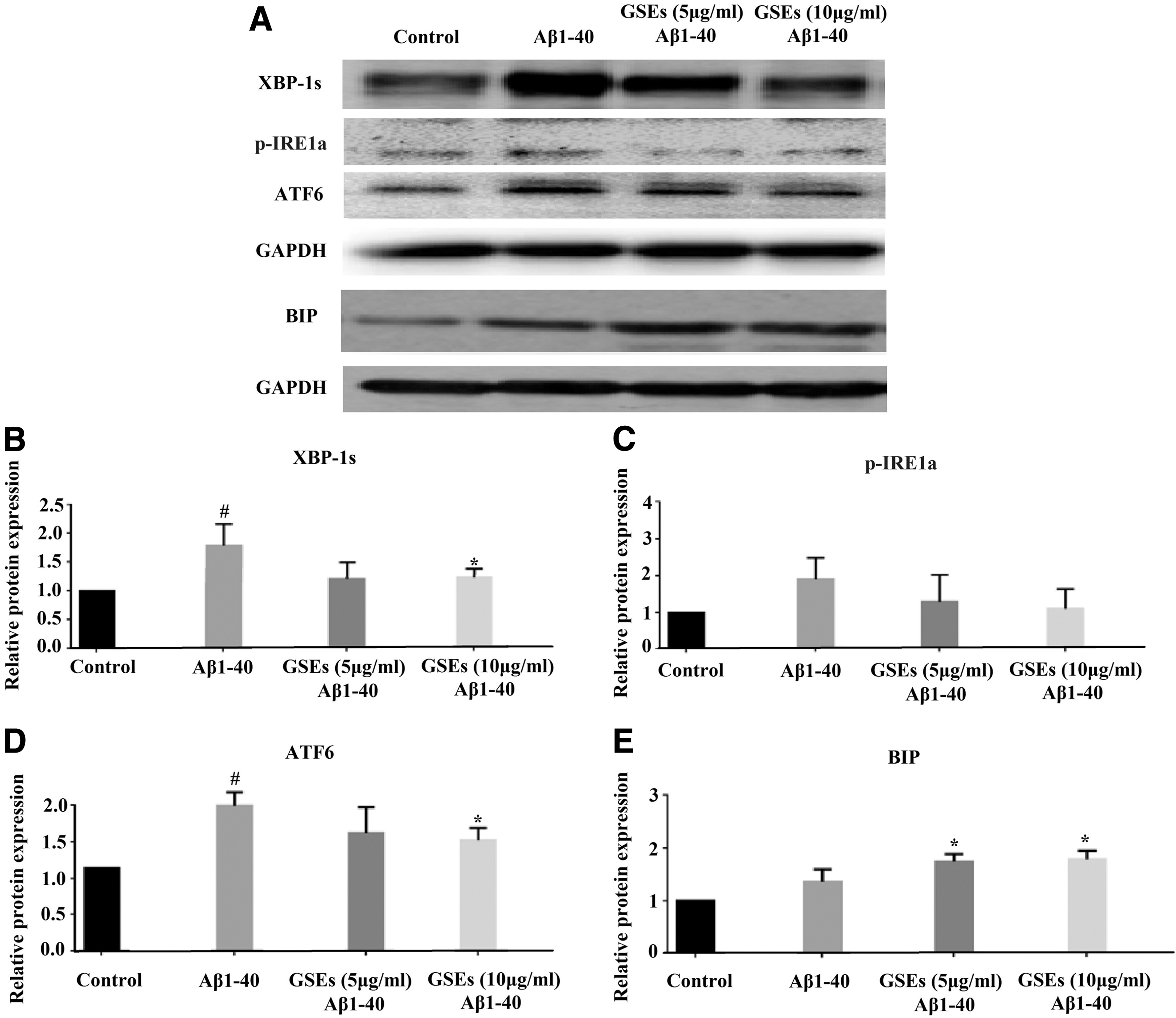
GSEs affect Aβ1–40-induced activation of endoplasmic reticulum stress. XBP-1s
GSEs alleviate Aβ1–40-induced increases in proinflammatory cytokines partly through promotion of Bip expression
To gain further insight into whether Bip is necessary for mediating the GSE-inhibiting effect on Aβ1–40-induced increases of proinflammatory cytokines, Bip was knocked down in ARPE-19 cells using siRNA (Fig. 5A, B). Silencing of Bip protein expression blocked the inhibiting effect of GSEs on mRNA levels of IL-1β (Fig. 5C), IL-6 (Fig. 5D), IL-8 (Fig. 5E), and IL-18 (Fig. 5F), suggesting that GSEs inhibit Aβ1–40-induced increases of these cytokines in a Bip-dependent manner. However, GSEs also attenuated Aβ1–40-induced IL-12 mRNA expression in cells in which Bip had been knocked down (Fig. 5G). This suggests that the inhibitory effects of GSEs on IL-12 mRNA expression may not be related to Bip.
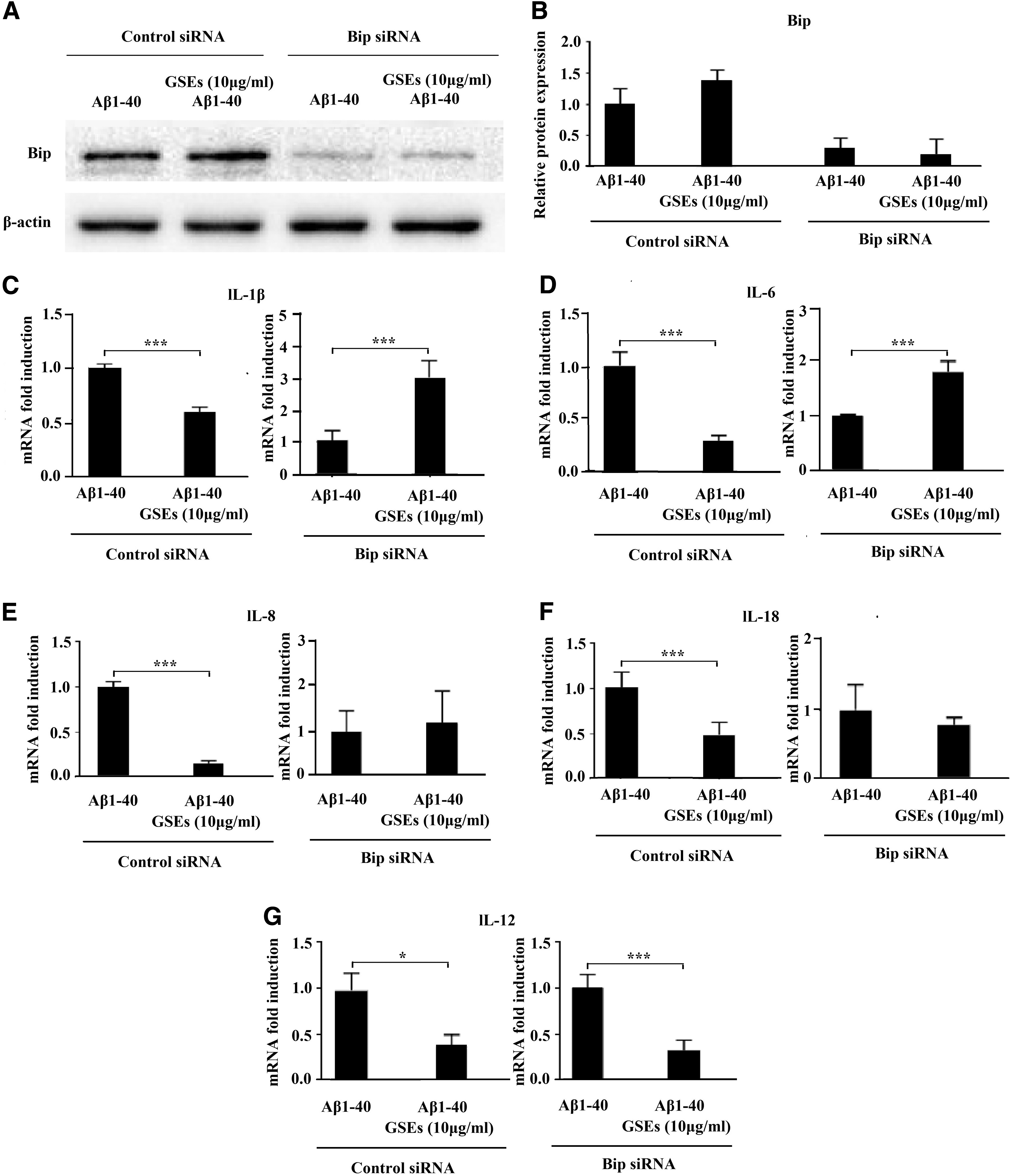
GSEs attenuated Aβ1–40-induced mRNA expression of proinflammatory cytokines in ARPE-19 cells partly by increasing GRP78 expression. The representative figures
Discussion
In this study, we demonstrated that GSEs inhibited Aβ1–40-induced increases in mRNA levels of proinflammatory cytokines in mouse retinas and ARPE-19 cells. The present data also showed that the attenuating effects of GSEs on proinflammatory cytokines might be associated with alleviating the ER stress markers—XBP-1s, p-IRE1, and ATF6—and promoting the expression of the unfolded protein response (UPR) chaperone, Bip.
First, we showed that GSEs attenuated the impairment of the RPE layer in mice intravitreally injected with Aβ1–40. Intravitreally injected Aβ1–40 in mice or rats has been successfully used to make in vivo models of Aβ stimulation in the retina. 6,13 Although this model is an acute retinal degeneration model rather than a progressive pathological model, 14 this model still is useful for investigating the pathogenesis of age-related retinal degeneration. Previous studies have shown that intravitreally injected Aβ1–40 reaches RPE cells and is abundant at day 4, resulting in an impairment of the retina. 8 Our present experimental results further demonstrated the Aβ-induced impairment of the RPE layer using H&E staining in the retina, whereas the RPE cells in control mice appeared normal in morphology (Fig. 1A).
RPE cells are important for the transport of nutrients and water, phagocytosis, and immune responses; hence, the impairment of RPE cells causes retinal injury. 15 As shown in Figure 1B, apoptosis was found in the retina of mice with intravitreally injected Aβ1–40 through TUNEL staining. GSE supplementation reversed both the damage of the RPE layer and apoptosis in the retina induced by Aβ1–40.
Moreover, inflammatory cytokines were increased within the retina in age-related retinal degeneration. 9,16 This study showed that mRNA expression levels of inflammatory cytokines—including IL-12, IL-6, IL-1β, and IL-18—were significantly increased under Aβ1–40 stimulation on day 4 in RPE–choroid complexes (Fig. 1C), which is consistent with the results of a previous study. 8 These cytokines have been found to be closely related to age-related retinal degeneration. 17 This study demonstrated that the oral supplementation of GSEs significantly inhibited the increase of these inflammatory cytokines in the RPE–choroid complex.
Furthermore, we showed that GSEs alleviated Aβ1–40-induced increases in the mRNA levels of IL-12, IL-6, IL-1β, IL-18, and IL-8 in ARPE-19 cells, indicating that the protective effect of GSEs against Aβ-induced retinal injury might be at least partly due to the inhibition of inflammatory cytokines in RPE cells. Other retinal cell types, such as ganglion cells and Muller cells, may also contribute to proinflammatory changes in the retina 18 and may be responsive to GSEs. For example, our previous study demonstrated that GSEs reduced hyperglycemia-induced increases in proinflammatory cytokines in Muller cells. 12 This study mainly focused on RPE cells due to their important role in age-related retinal degeneration. This is the first study to show that GSEs inhibit Aβ-induced mRNA expression of inflammatory cytokines in RPE–choroid complexes and in ARPE-19 cells.
Owing to the pivotal role of ER stress in Aβ-induced expression levels of proinflammatory cytokines, 19 we further studied whether GSEs imposed an effect on Aβ-induced ER stress in ARPE-19 cells. Results showed that the expression of p-IRE1α was elevated in Aβ1–40-treated cells (Fig. 4). Moreover, XBP-1 splicing, a result of IRE1α activation, was also induced after Aβ1–40 exposure. In parallel, Aβ1–40 increased the level of ATF6 protein. These changes indicated an induction of ER stress in RPE cells. GSE treatment could attenuate the activation of IRE1α/XBP1 and ATF6 pathways (Fig. 4). This is consistent with previous studies demonstrating that grape seed proanthocyanidin attenuated ER stress and inhibited inflammation in the retina. 20
However, the correlation between ER stress and the overexpression of proinflammatory cytokines in grape proanthocyanidin-treated RPE cells requires further investigation. In addition, we found that GSEs increased the levels of GRP78/Bip (78 kD glucose-regulated protein) expression. Bip is a UPR chaperone with cell-protective properties. During ER stress, BiP dissociates from its partners such as ATF6 and IRE1, and attenuates ER stress through promoting protein folding and degradation, which may initiate a survival signaling pathway. 21 We thus hypothesized whether GSEs attenuated Aβ1–40-induced increases in proinflammatory cytokines by promoting Bip expression. As shown in Figure 5C–F, the inhibitory effects of GSEs on IL-1β, IL-6, IL-8, and IL-18 mRNA levels were blunted by Bip knockdown.
Our previous research showed that procyanidin B2, a pivotal proanthocyanidin in grape seed, inhibited the phosphorylation of IRE1a and protected ARPE-19 cells against oxidative stress by promoting Bip expression, 22 which is consistent with this study. However, some other reports have demonstrated that grape seed proanthocyanidins inhibit Bip expression. For example, Fu et al. showed that grape seed proanthocyanidins attenuated ER stress-associated neuronal apoptosis by inhibiting Bip in ischemic stroke mice. 23 Wang et al. demonstrated that grape seed proanthocyanidins reduced Bip expression and protected cardiomyocytes against hypoxia/reoxygenation injury. 24 The different effects of grape seed proanthocyanidins on Bip might be due to cell type-specific differences in responses and stimuli, and should be further investigated.
In conclusion, this study demonstrated that GSEs attenuated Aβ-induced increases in proinflammatory cytokines in mouse retinas and ARPE-19 cells. The protective activities may be related to the inhibition of ER stress and promotion of the UPR (Fig. 6). The results showed a potential application of GSEs to reduce the risk of age-related retinal degenerative disease.

Schematic diagram depicting a possible mechanism of action of GSEs.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the National Natural Science Foundation of China (Grant No. 31571837) and the Scientific and Technical Committee of Shanghai (Grant No. 19DZ2284200).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.