
Abstract
A variety of natural compounds have been used to reduce the oxidative stress associated with Alzheimer's disease, and many of these defend cells from oxidative stress-induced neuronal toxicity. In this study, the protective effect of radish (Raphanus raphanistrum) extract was investigated in mice and PC12 cells. In vivo behavioral tests were completed to examine the protective effects of the extract on amyloid beta (Aβ)-peptide1–42-induced learning and memory impairment in a mouse model. The extract increased spontaneous alternation behaviors and step-through latency in mice. We discovered that administration of the extract reduced lipid peroxidation and Aβ aggregation in a biochemical study of mice brain tissues. Treatment with the extract also increased acetylcholine and catalase activity in the brain. Furthermore, the radish extract attenuated H2O2-induced oxidative stress in cells. Through sequential fractionation of the radish extract, the active compound was identified as oleamide. These results suggest that the radish extract could have a protective role against oxidative stress-induced neuronal toxicity, possibly owing to the antioxidative activity of oleamide.
Introduction
Alzheimer's disease (AD) is a neurodegenerative disorder of major importance and is the principal cause of dementia. The disease is named after Dr. Alois Alzheimer. In 1906, Dr. Alzheimer observed changes in the brain tissue of a patient who had died of an unconventional mental illness. He found many abnormal amyloid plaques and neurofibrillary tangles in the patient's brain tissue. 1
AD is a chronic neurodegenerative disease that generally starts slowly and steadily worsens over time. The cause of AD is poorly comprehended. About 70% of the risk is believed to be transferred from parents, with many genes usually involved. Other risk features include a history of head injuries, depression, and hypertension. 2 The disease process is associated with plaques and neurofibrillary tangles in the brain. Many researchers suggest that the accumulation of amyloid beta-peptide (Aβ) is a critical step in the initiation of AD. 3,4 Aβ has been shown to induce cytotoxic actions toward neuronal cells and animal. Deposition of Aβ leads to free radical-induced oxidative stress. 5 This oxidative stress is showed by the formation of reactive oxygen species (ROS), lipid peroxidation, and DNA oxidation, as well as the subsequent modification of proteins by the reactive lipid peroxidation products. 6 Since free radicals partially mediate the neurotoxic effects of Aβ, the removal of free radicals or the prevention of their formation might be beneficial in the treatment of AD.
Radish (Raphanus raphanistrum subsp. sativus) is an edible root vegetable of the Brassicaceae family. Radishes are grown and consumed throughout the world and are mostly used as a salad. There are various varieties, varying in size, flavor, color, and length of time taken to mature. Radish contains various nutrients, including potassium, calcium, sodium, and vitamins such as vitamin A, B, C, and K. It also provides minerals such as iron, magnesium, phosphorus, and zinc. Korean radish, such as other radishes, is an annual or biennial crop grown for its taproot.
In this research, we studied the antioxidative effects and protective effects of radish extracts against H2O2-induced neuronal cell death in cultured PC12 cells. The PC12 cell line was derived from a rat pheochromocytoma, an adrenal medullary tumor. After its derivation, this cell line was widely used for studies as a neuronal model. 7,8 We also established a mouse model of dementia with learning and memory dysfunction by injecting Aβ into the ventricle of the mouse brain. Biochemical testes have demonstrated a connection between changes in lipid peroxidation, Aβ aggregation, catalase activity, and damage to cholinergic neurons in Aβ-treated mice. Active compounds were isolated and identified using high-performance liquid chromatography (HPLC), and Aβ-injected mice were used to observe the effects of the active compound on memory impairment.
materials and Methods
Chemicals
Aβ 42–1 was purchased from Bachem (Bubendorf, Switzerland), and 2,2′-azobis-(2-amidinopropane) dihydrochloride (AAPH) was purchased from Wako Chemicals USA, Inc., (Richmond, VA, USA). Other chemicals were purchased from Sigma Chemicals (St. Louis, MO, USA).
Sample preparation
Dried radish was purchased from the Gyeongdong medicinal herb market (Seoul, South Korea). The dried radish (4 kg) was ground into a powder and mixed in ethanol (20 L) by shaking for 24 h at 1.57g. The ethanol extract was filtered through No. 42 filter paper (Whatman International Ltd., Middlesex, UK) with five replicates, and evaporated in a rotary evaporator (Eyela, Tokyo, Japan).
Antioxidative capacity measurement
The antioxidant capacity of the radish extract was compared using a typical 2,2-diphenyl-1-picrylhydrazyl (DPPH) assay, as described previously. 9 The 1.5 mL of DPPH radical solution (0.2 mM) was mixed with 0.5 mL of sample extract solution for 30 min. Finally, the absorbance was measured at 517 nm (SpectraMax M2, Molecular Devices, CA, USA).
Vitamin C equivalent antioxidant capacity assay using ABTS radicals
The 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) radical anion was used according to a previous research. 10 Briefly, 1.0 mM AAPH was mixed with 2.5 mM ABTS2− in phosphate-buffered saline (PBS; pH 7.4). The mixed solution was heated at 70°C for 30 min. The resulting ABTS solution was diluted to an absorbance of 0.650–0.020 at 734 nm. The sample (20 μL) was mixed with the ABTS radical solution (980 μL). The mixture was incubated at 37°C for 10 min. The decrease in absorbance at 734 nm was measured after 10 min.
Cell culture
The rat pheochromocytoma (PC12) cell line (CRL-1721) was purchased from the American Type Culture Collection (Manassas, VA, USA). Other materials were obtained from Gibco-BRL (Grand Island, NY, USA). The PC12 cells were cultured in the RPMI 1640 medium with 10% (v/v) horse serum from a donor herd, 5% (v/v) fetal bovine serum, and 1% (v/v) antibiotic/antimycotic. The cells were cultured on 100 mm tissue culture dishes (Falcon; BD Biosciences, Franklin Lakes, NJ, USA) and maintained in an incubator (37°C, 5% CO2 atmosphere).
DCF-DA assay
The PC12 cells were pretreated for 48 h with the sample, followed by exposure to H2O2 for 2 h. At the end of the treatment, the cells were incubated with 50 μM 2′,7′-dichlorofluorescin diacetate (DCF-DA) for 50 min at 37°C, and dichlorofluorescein was quantified using a microplate reader (SpectraMax M2; Molecular Devices, LLC, San Jose, CA, USA) using 485 nm excitation and 535 nm emission fluorescence filters. The results are presented as a percentage relative to the oxidative stress of the control cells (set to 100%).
Oxidative stress (%) = 100 × (quantified DCF of sample and H2O2-treated cells/quantified DCF of untreated control cells).
MTT assay
Cells were incubated with the sample for 48 h before H2O2 addition, followed by treatment with or without H2O2 for 2 h. Reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) was initiated by the addition of 10 μL of MTT stock solution (2.5 mg/mL) per well, followed by a 3-h incubation of the plates at 37°C. The reaction was stopped by adding 150 μL of DMSO. The absorbance was measured at 570 nm and 630 nm reference.
Analysis of DNA fragmentation
The PC12 cells were pretreated for 48 h with the radish extract, followed by exposure to H2O2 for 24 h. The cells were harvested and washed with PBS. DNA was extracted using the DNA
Animals
ICR mice (Institute of Cancer Research, 5-week-old males) were purchased from DBL Co. (Chungbuk, South Korea). The radish extract was mixed with feed for 4 weeks. Aβ 1–42 was injected via intracerebroventricular (ICV). The control group was injected with the reverse fragment, Aβ 42-1 (nontoxic, 410 pmol/mouse), and the Aβ group was injected with Aβ 1–42 (toxic, 410 pmol/mouse). The sample groups were injected with Aβ 1–42 after their diets were supplemented with radish extract (400, 800, and 1200 mg/kg body weight per day) or oleamide (10, 20, and 40 mg/kg body weight per day). The experimental procedures were approved by and followed the guidelines established by the Animal Care and Use Committee of Korea University (KUIACUC-2017-150).
Y-maze test
The Y-maze test was performed 3 days after the Aβ administration. Each mouse was placed at the end of one arm and permitted to move freely through the maze during an 8-min period. The sequence of arm entries was recorded. The percentage of spontaneous alternation behaviors was calculated as the ratio of actual to possible alternations. 11
Passive avoidance test
The passive avoidance test was performed 7 days after the Aβ injection. The chamber consisted of an illuminated room connected to a dark room. An acquisition trial was performed on day 1. Each mouse was placed in the chamber and left for 1 min with no light or shock, followed by a 2-min period with light and no shock to habituate the mouse to the chamber. The mice were individually placed in the illuminated chamber. Electric shock (0.5 mA for 1 sec) was delivered through the floor grid after entering the dark chamber. Each mouse was placed in the illuminated chamber again, 24 h later (retention trial). The interval between placement in the illuminated chamber and entry into the dark chamber was measured. The maximum testing limit for step-through latency was 300 sec. 11
Determination of lipid peroxidation
The mouse brains were homogenized in cold PBS. The homogenates were centrifuged at 33,600 g for 10 sec. The malondialdehyde (MDA) level was measured for lipid peroxidation by monitoring the formation of thiobarbituric acid reactive substances. 12
Quantification of ACh content
Acetylcholine (ACh) content was measured using the Hestrin method as described previously. 13 Brain homogenate (1 mL) was mixed with 2 mL of alkaline hydroxylamine reagent. After 1 min, 1 mL of hydrochloride solution and 1 mL of iron solution were added. The density of the purple–brown color was determined (540 nm).
Determination of catalase activity
Catalase activity was measured using a previous method. 12 Brain homogenate (50 μL) and 650 μL of 50 mM phosphate buffer (pH 7.0) were added into a quartz cuvette. The reaction was initiated by 30 mM H2O2 (300 μL). The breakdown capability of H2O2 was monitored at 240 nm.
Quantification of Aβ aggregation
Aβ aggregation was measured using a protocol modified from the Congo red binding assay. 14,15 Brain homogenates (75 μL) were mixed with 225 μL of Congo red solution (20 μM) and incubated at room temperature for 30 min before measurement of the absorbance. The concentration of the Aβ-fibrillar species in the peptide solutions was determined using a ratio of A540 to A405. A540 and A405 are the absorbances of the Aβ peptide and Congo red fibrillar at the wavelengths of 540 nm and 405 nm, respectively.
Isolation and identification of the active compound
The active compound was isolated by solvent partition, silica gel column chromatography, thin-layer chromatography (TLC), and HPLC. The evaporated radish ethanol extract (1194 g) was dissolved in distilled water (1 L) and partitioned with hexane (3 L × 3), chloroform (3 L × 3), and ethyl acetate (3 L × 3) sequentially for 24 h. Each layer was separately concentrated in a rotary evaporator. The first ethyl acetate fraction (2.44 g) was isolated by silica-gel column chromatography. The column was eluted with a gradient chloroform and ethanol (10:0, 9:1, 8:2, 7:3, 6:4, 5:5, 4:6, 3:7, 2:8, 1:9, and 0:10, v/v, sequentially; repeated three times). The second fraction of the 8:2 elution was concentrated in an evaporator. The evaporated sample (150 mg) was isolated by TLC. The fraction of the sixth band (R f = 0.39) was collected, and the isolated sample (31.9 mg) was eluted in ethanol and analyzed with an HPLC system (YL9100) using a C18 column (4.6 × 250 mm, 5 μm Capcell Pak; Shiseido, Chuo-ku, Tokyo, Japan). The structure of the isolated active compound was determined by GC-MS and 13 C/ 1 H-NMR (Korea Basic Science Institute, Western Seoul Center).
Statistical analysis
Results are expressed as mean ± SD. Data were analyzed by one-way ANOVA, followed by Duncan's multiple range tests using SAS software (Cary, NC, USA).
Results
Radical scavenging effect of radish extract
The radical scavenging activity of the radish extract was confirmed using DPPH and vitamin C equivalent antioxidant capacity (VCEAC) assays. The change in DPPH absorbance after the addition of radish extract was used as an index of the antioxidant capacity. The radish extract scavenged DPPH radicals in a dose-dependent manner. As shown in Figure 1A, the coefficient of determination between the radish extract and DPPH radical scavenging was calculated (R 2 = 0.9924). Concentration of the extract necessary to decrease the initial concentration of DPPH by 50% (IC50) under the experimental conditions was calculated to be 11.18 mg/L. The radical scavenging potential of the radish extract was estimated using ABTS radical anions. The VCEAC standard curve was calculated by plotting the decrease in absorbance against the concentration of vitamin C. The antioxidant capacity of the radish extract was then converted to VCEAC. The correlation between the radish extract and VCEAC was 0.9901. The antioxidant capacity of the radish extract was higher than the capacity of vitamin C (Fig. 1B, C). In both the DPPH and VCEAC assays, the radish extract was shown to possess potent antioxidative activity.
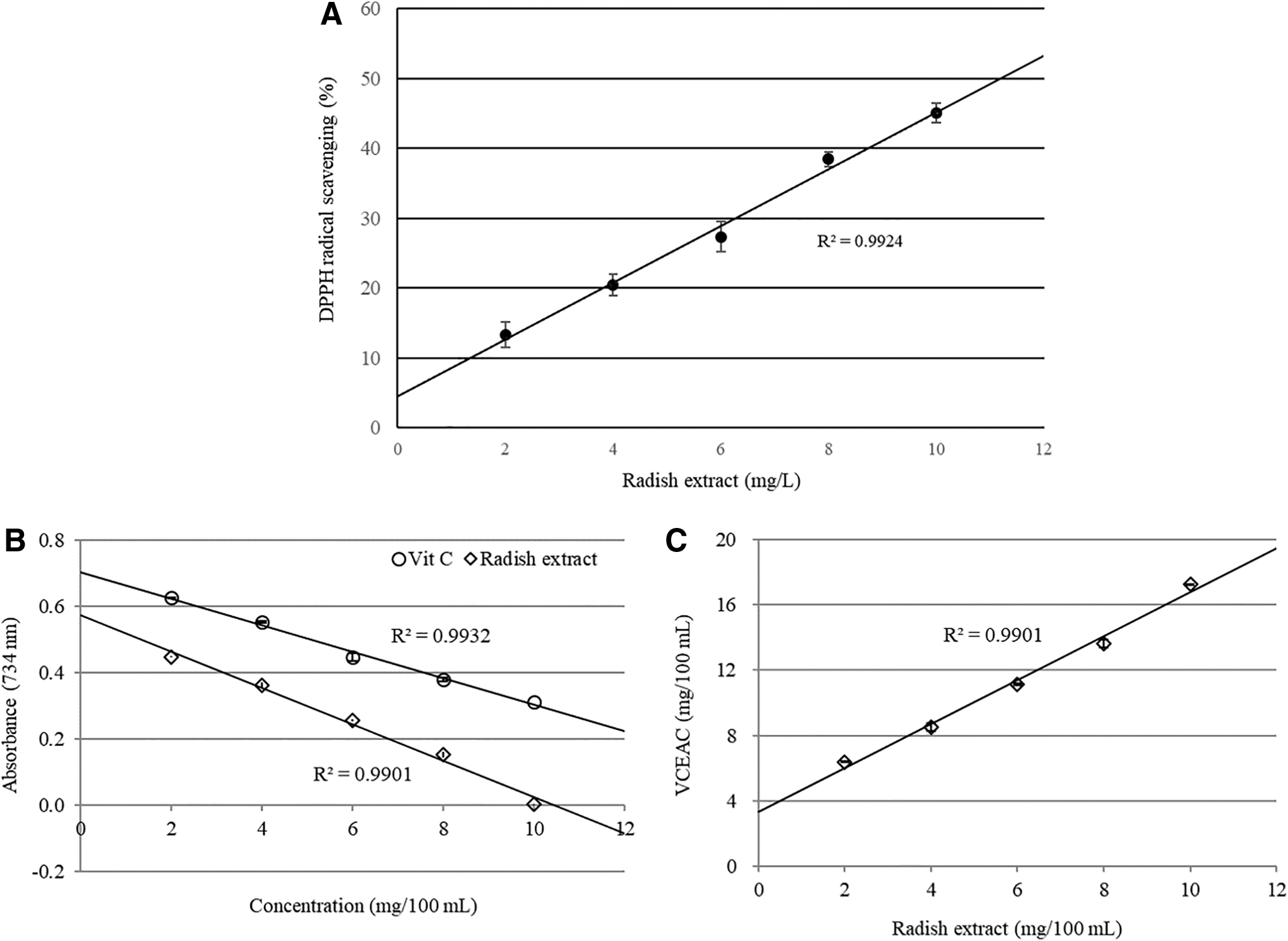
Antioxidant capacity of radish extract.
Effect of radish extract on DNA fragmentation
The defensive effect of the radish extract on H2O2-induced DNA fragmentation was estimated using gel electrophoresis. A DNA fragmentation pattern was discovered on H2O2-treated PC12 cells. Pretreatment of cells with the radish extract diminished DNA fragmentation compared with the cells treated with H2O2 alone (Fig. 2).
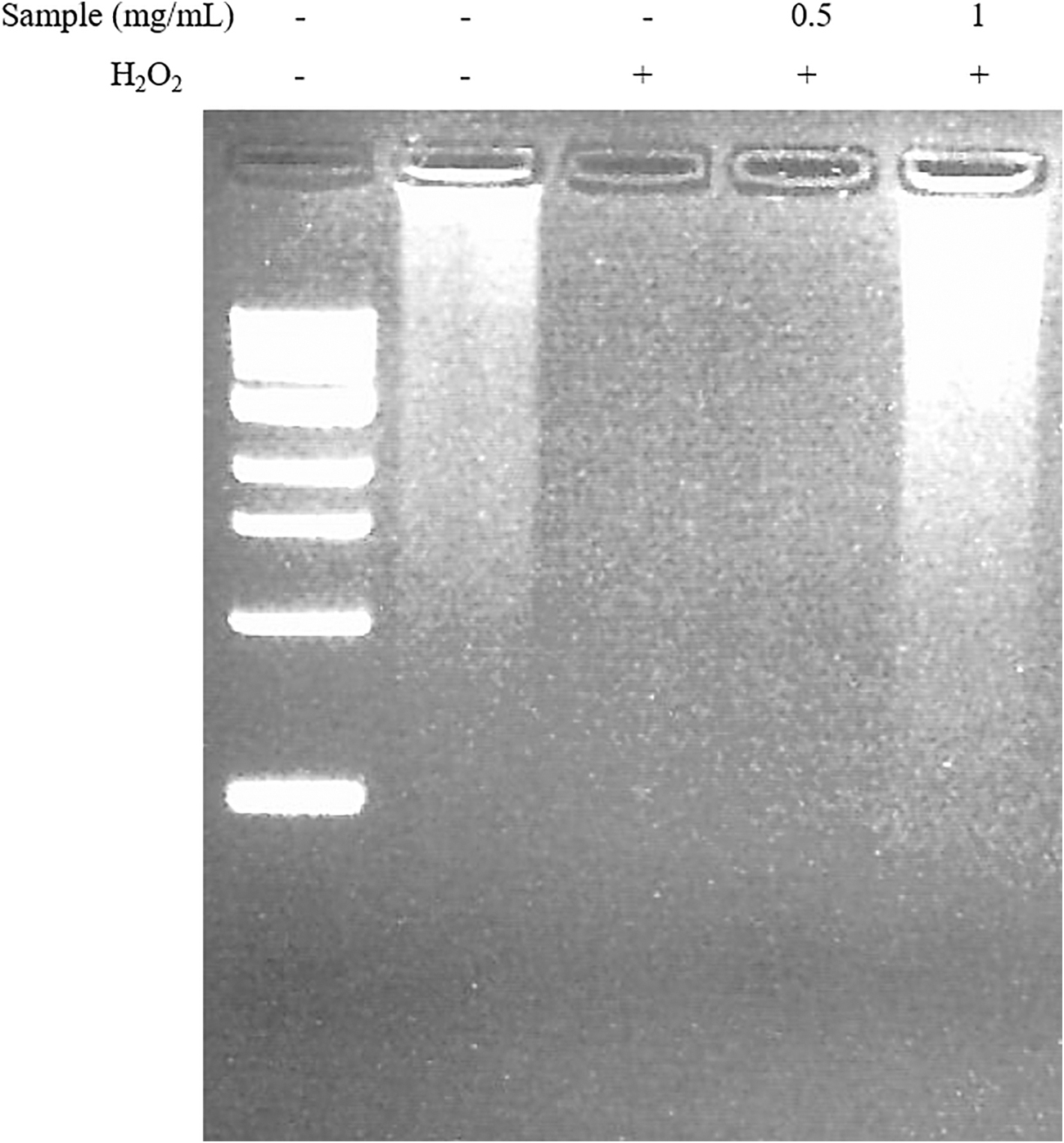
The effect of the radish extract on H2O2-induced DNA fragmentation in PC12 cells. DNA extracts from PC12 cells exposed to H2O2 with various concentrations of the extract were subjected to agarose gel electrophoresis. Lane 1, 0.5–10.0 kb DNA ladder marker; lane 2, control; lane 3, 200 μM H2O2 alone; lanes 4–5, radish extract treatment +200 μM H2O2.
Y-maze test
The Aβ 1–42-injected group (Aβ group) showed significantly reduced spatial working memory (17% decrease in alternation behavior) compared with the control group (Fig. 3). The radish extract diet group exhibited increased spontaneous alternation behavior even after Aβ injection. However, the total number of arm entries did not change among all of the experimental groups (data not shown).
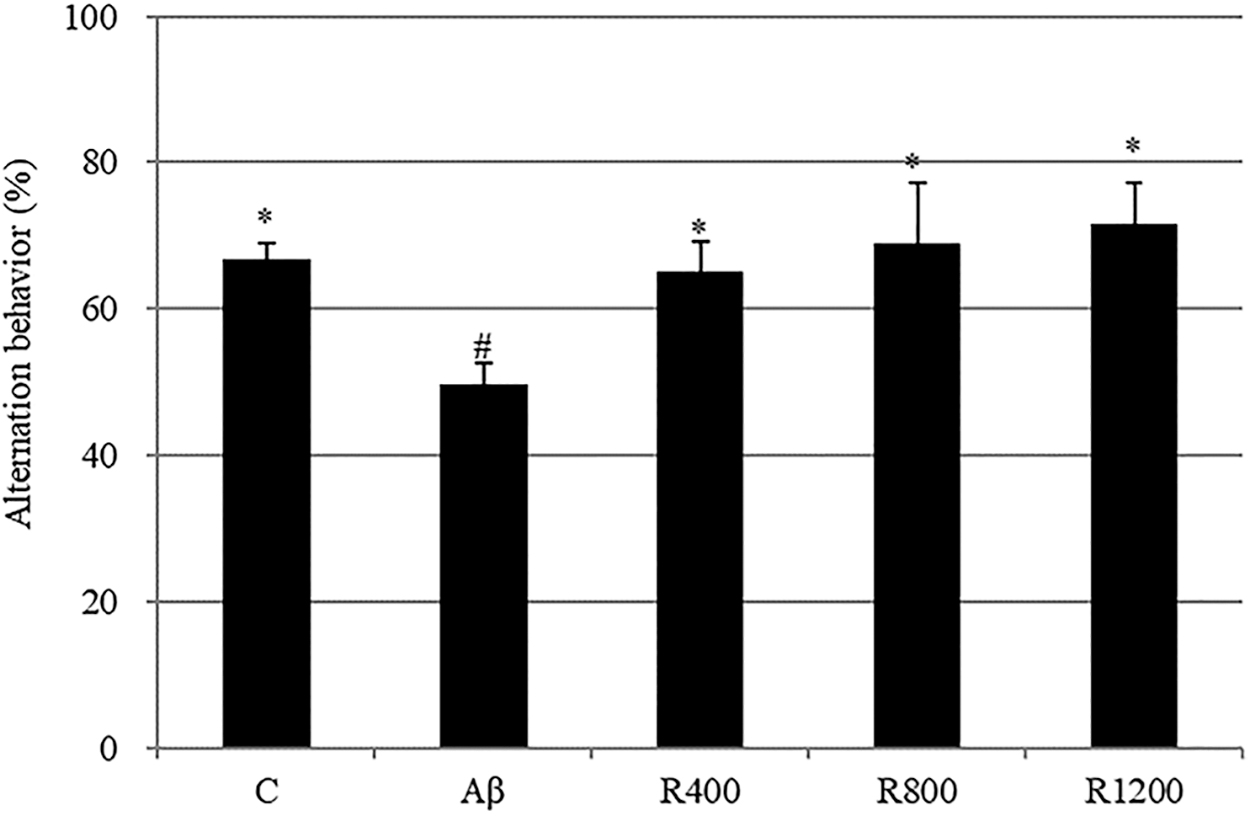
The effect of the radish extract on spontaneous alternation behavior in the Y-maze test. Controls were injected with Aβ 1–42. Aβ was injected with 410 pmol of Aβ 1–42 per mouse. Sample groups (R400, R800, and R1200) were injected with Aβ 1–42 followed by feeding with radish extract (400, 800, and 1200 mg/kg per day, respectively). Spontaneous alternation behaviors were measured during 8 min. Values indicate the mean ± SD (n = 8). *, # Represent statistical differences between groups (P < .05).
Passive avoidance test
The Aβ 1–42-injected group (Aβ group) exhibited a significant reduction (40-sec decrease) in step-through latency time compared with the control group. The radish extract diet delayed Aβ 1–42-induced impairment in mice during the passive avoidance test (Fig. 4).
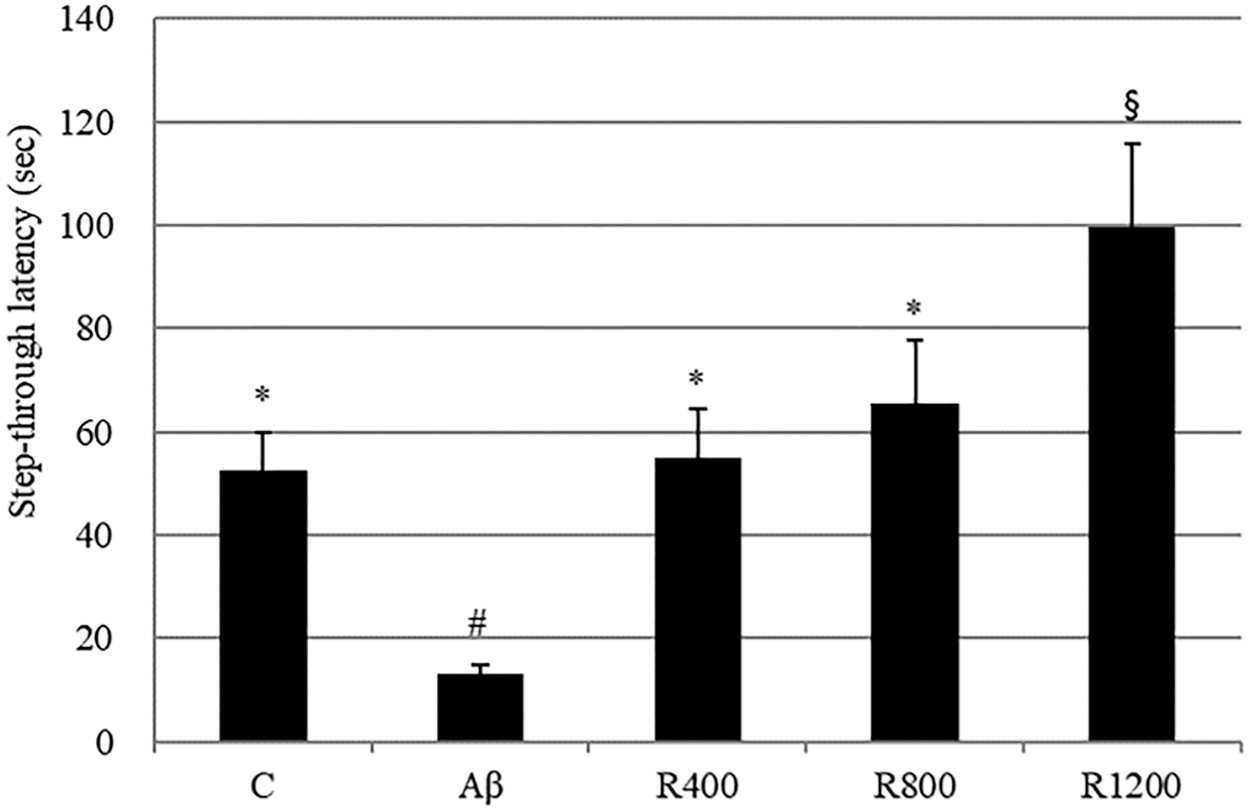
The effect of the radish extract on step-through latency in the passive avoidance test. Controls were injected with Aβ 1–42. Aβ was injected with 410 pmol of Aβ 1–42 per mouse. Sample groups (R400, R800, and R1200) were injected with Aβ 1–42 followed by feeding with radish extract (400, 800, and 1200 mg/kg per day, respectively). The step-through latency was determined during 300 sec. *, #, § Represent statistical differences between groups (P < .05). Values indicate the mean ± SD (n = 8).
Aβ aggregation
Congo red assay was performed to confirm the accumulation of Aβ in the brain tissue. As shown in Figure 5, Aβ aggregation was more abundant in the Aβ group than in the control group, and Aβ accumulation was decreased in the radish extract-fed group.
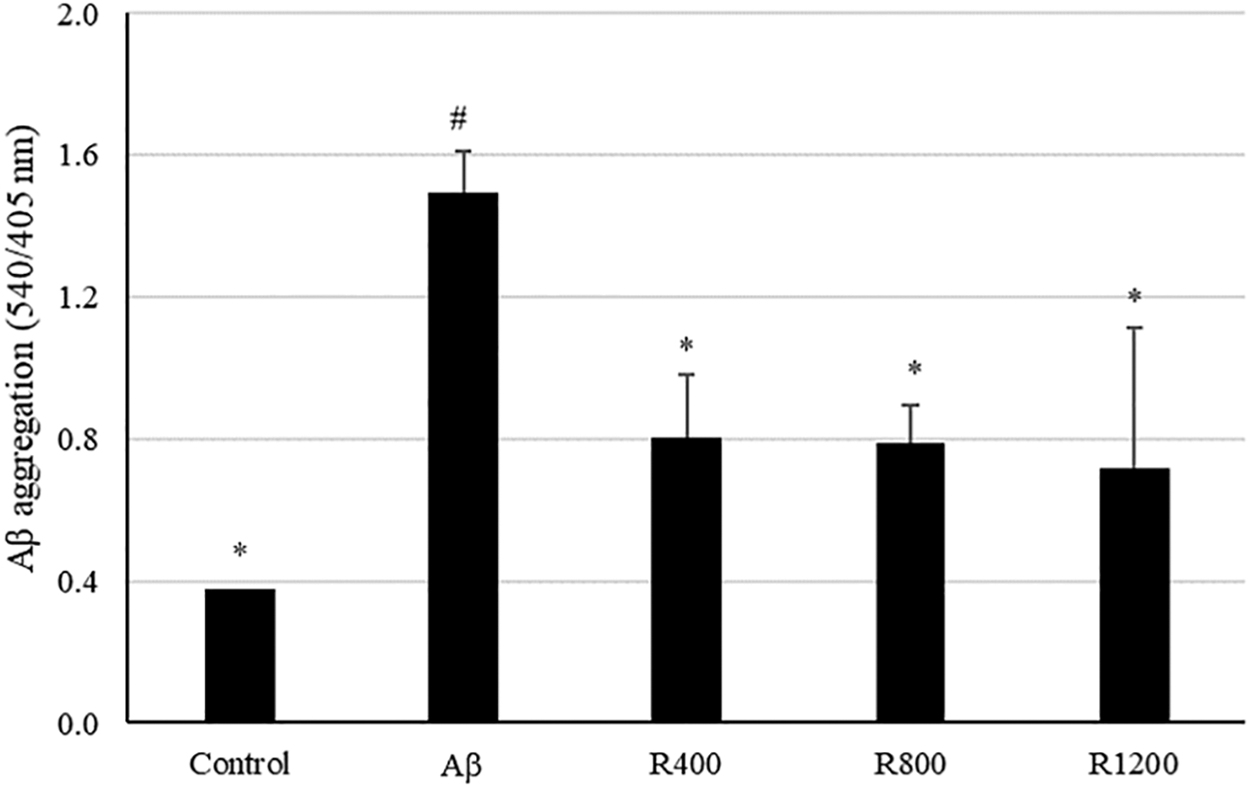
Protective effect of radish extract against Aβ aggregation in mouse brain tissue. Controls were injected with Aβ 1–42. Aβ was injected with 410 pmol of Aβ 1–42 per mouse. Sample groups (R400, R800, and R1200) were injected with Aβ 1–42 followed by feeding with radish extract (400, 800, and 1200 mg/kg per day, respectively). *, # Represent statistical differences between groups (P < .05). Values indicate the mean ± SD (n = 8).
Determination of lipid peroxidation
Lipid peroxidation was tested using brain tissue after behavior tests were completed. The MDA levels were dramatically increased (5.6 μmol/mg protein) in the Aβ 1–42-injected group compared with the control group. This increase indicated an increment of lipid peroxidation in the brains of Aβ-injected mice. Diets containing 400, 800, and 1200 mg/kg of radish extract overturned the effects of Aβ-induced lipid peroxidation (Fig. 6).
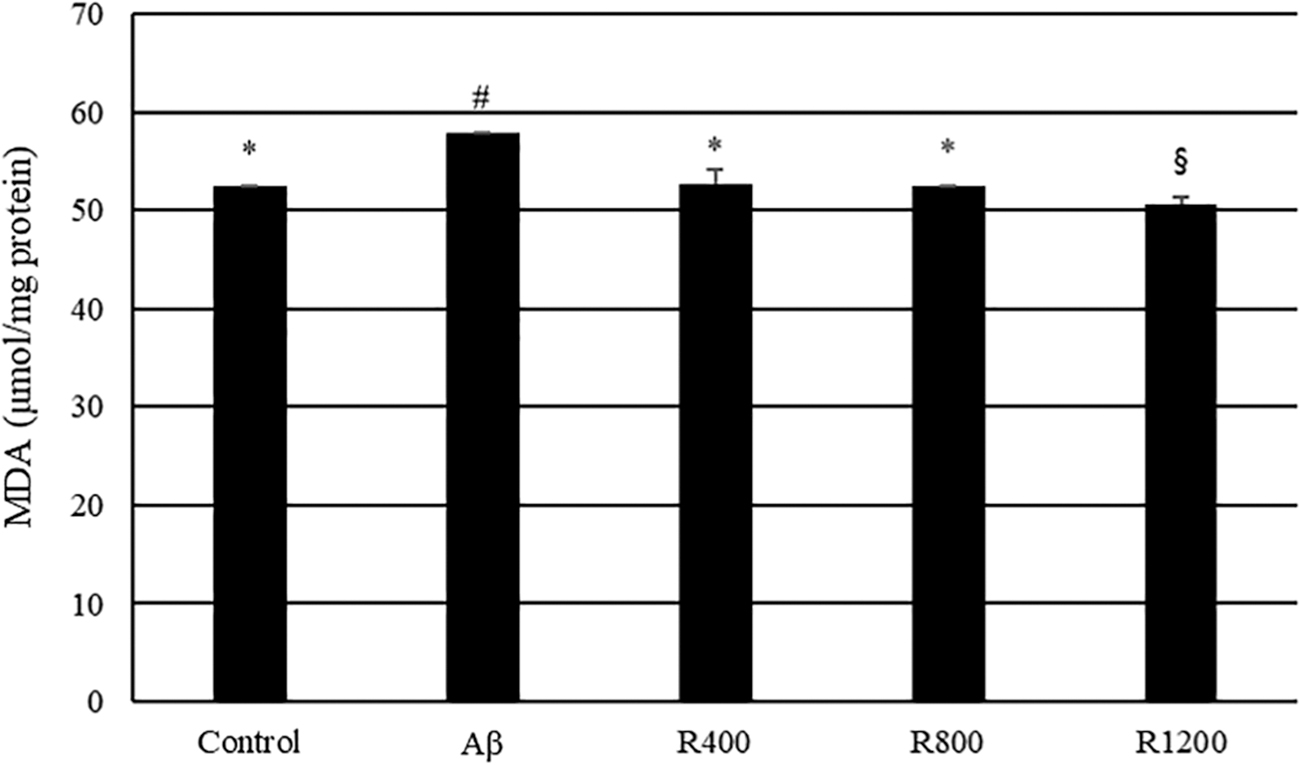
Effect of radish extracts on lipid peroxidation in the mouse brain. Controls were injected with Aβ 1–42. Aβ was injected with 410 pmol of Aβ 1–42 per mouse. Sample groups (R400, R800, and R1200) were injected with Aβ 1–42 followed by feeding with radish extract (400, 800, and 1200 mg/kg per day, respectively). *, #, § Represent statistical differences between groups (P < .05). Values indicate the mean ± SD (n = 8).
Catalase activity in mouse brain tissue
To confirm Aβ-induced damage to the mouse brain, the catalase activity was tested. No significant decrease in catalase activity was observed in the mice fed radish extracts. Antithetically, catalase activity was significantly decreased (0.26 U/mg protein) in the Aβ-treated mice in comparison with the control group (Fig. 7).
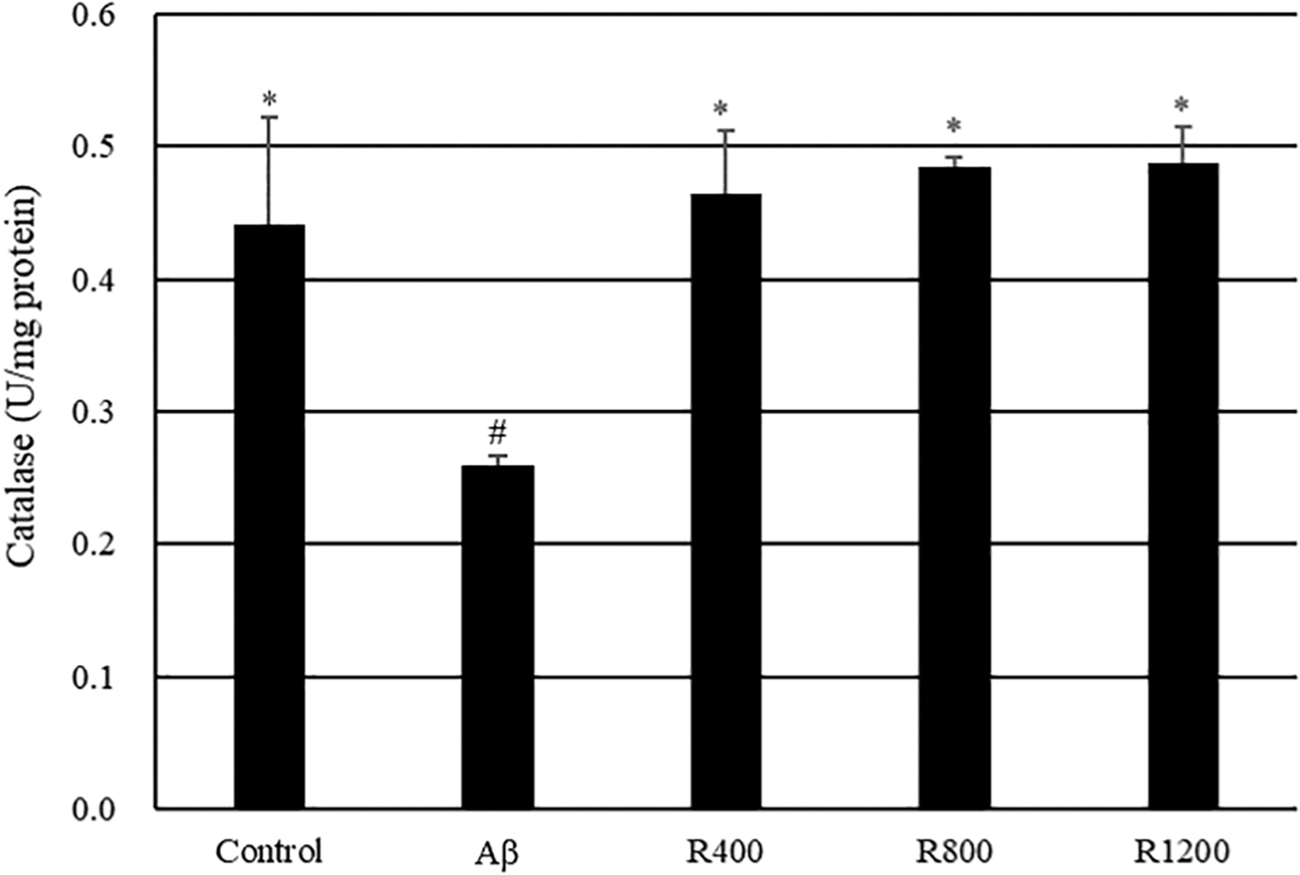
Effect of radish extracts on catalase activity in the mouse brain. Controls were injected with Aβ 1–42. Aβ was injected with 410 pmol of Aβ 1–42 per mouse. Sample groups (R400, R800, and R1200) were injected with Aβ 1–42 followed by feeding with radish extract (400, 800, and 1200 mg/kg per day, respectively). *, # Represent statistical differences between groups (P < .05). Values indicate the mean ± SD (n = 8).
ACh content
ACh content of mouse brain tissue was measured by spectrophotometric analysis. As shown in Figure 8, the ACh level was lower (39.29 mg/mg protein) in Aβ-treated mice than in the control group. In contrast, administration of the radish extract inverted the decrease in ACh content compared with that of mice treated with Aβ only.
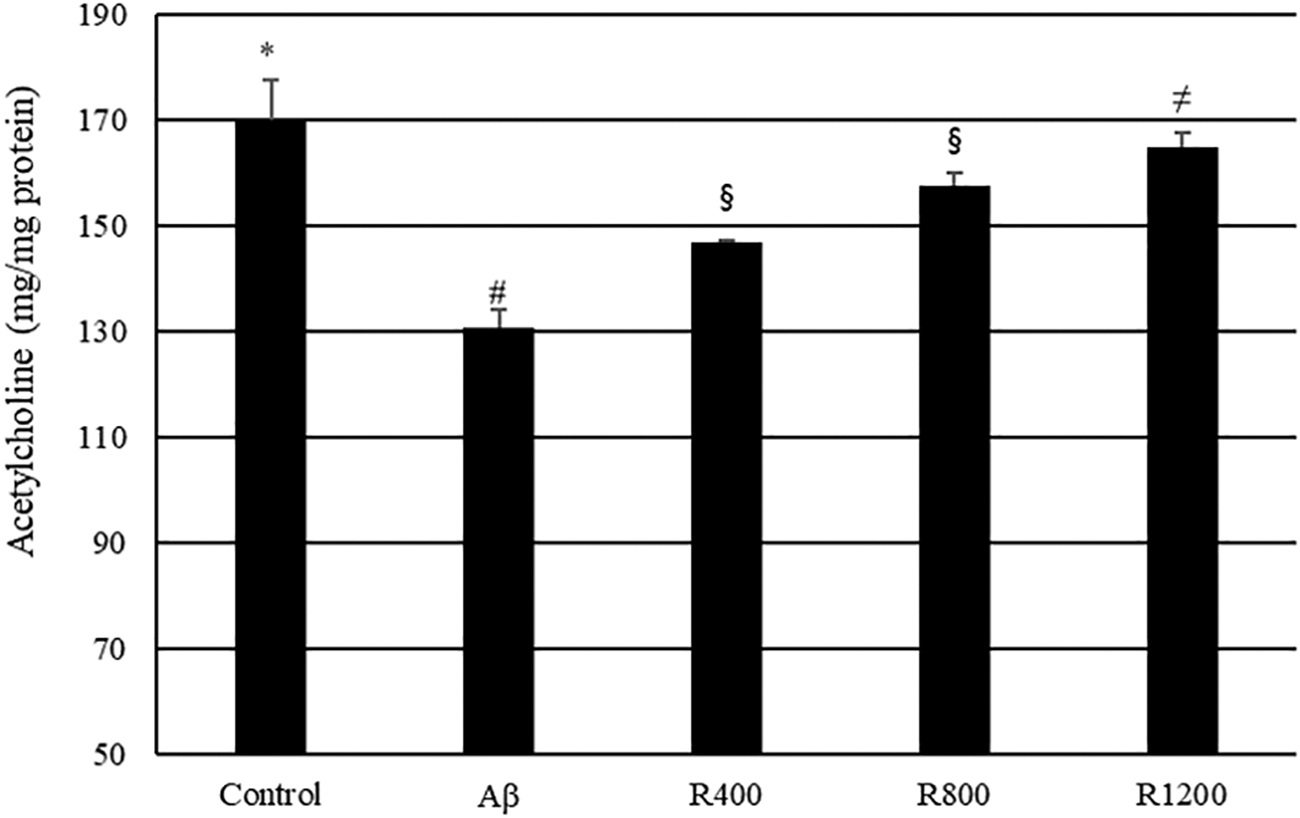
Effect of radish extracts on ACh contents in the mouse brain. Controls were injected with Aβ 1–42. Aβ was injected with 410 pmol of Aβ 1–42 per mouse. Sample groups (R400, R800, and R1200) were injected with Aβ 1–42 followed by feeding with radish extract (400, 800, and 1200 mg/kg per day, respectively). *, #, §, ≠ Represent statistical differences between groups (P < .05). Values indicate the mean ± SD (n = 8). ACh, acetylcholine.
Isolation of active compound
To isolate the active compound in the radish extract, solvent partitioning and open column chromatography were used. The extract was partitioned three times using n-hexane, chloroform, and ethyl acetate. The first fraction of ethyl acetate decreased oxidation most effectively (676.49%; Table 1). In the MTT reduction assay, this fraction exhibited the highest cell viability (105.30%; Table 1). The first fraction of ethyl acetate was separated into 33 subfractions by silica-gel open column chromatography. Among these fractions, the second fraction, with a chloroform to ethanol ratio of 8:2, exhibited positive activity in both the DCF-DA assay (81.51%) and the MTT reduction assay (105.21%; Table 2). To purify the active compound, the selected fraction was separated by TLC. The TLC plate was visualized by ultraviolet (254 nm and 365 nm), and the ratio of front values was measured. The separated band (R f = 0.39) showed the best effect (Table 3). The selected band was extracted and evaporated, and the sample obtained was eluted in ethanol and analyzed with HPLC. Major active compounds were isolated on the column. A significant peak appeared at 24 min (Fig. 9). The structure of the active compound was analyzed by GC-MS and 13 C/ 1 H-NMR and was identified as oleamide (Fig. 10).
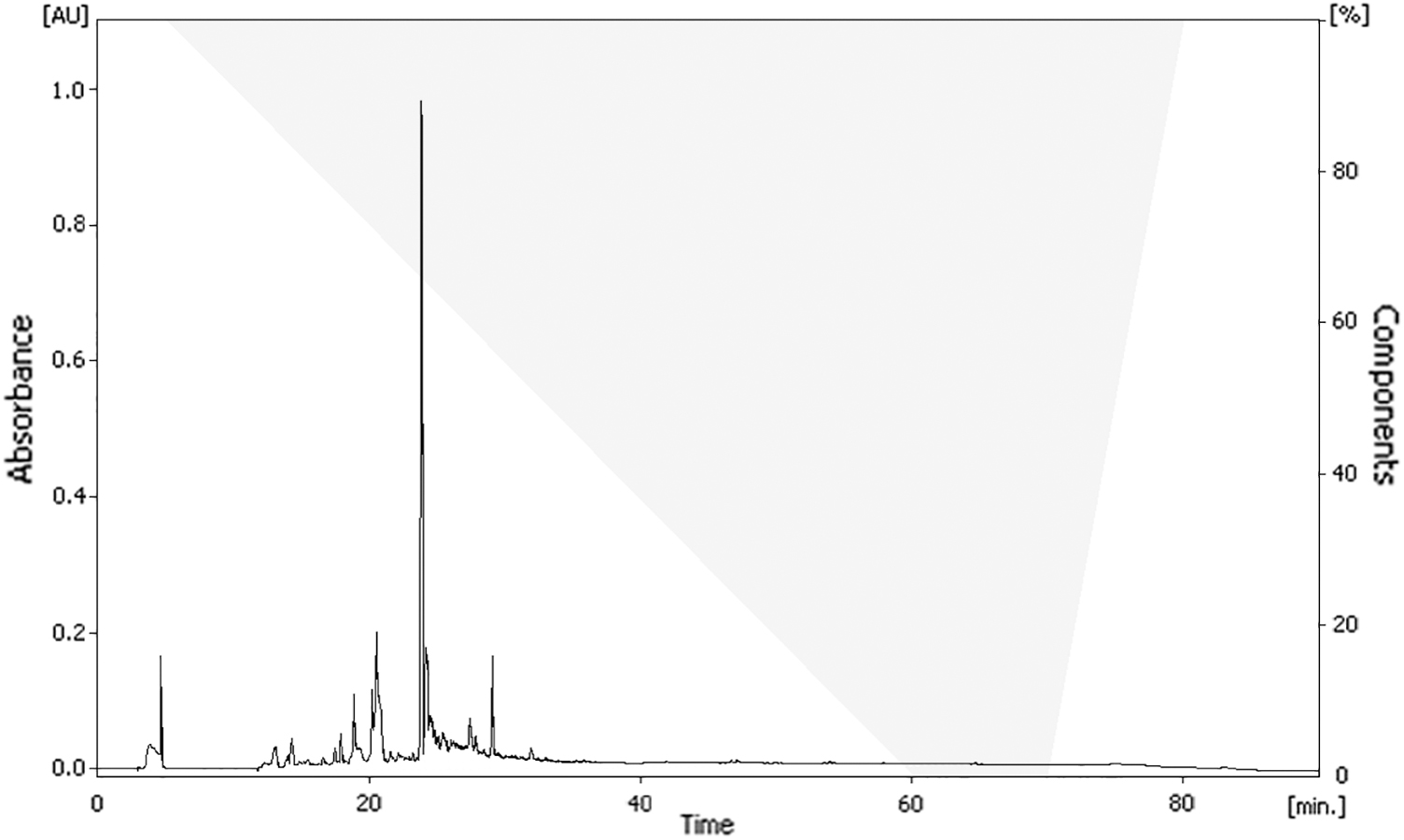
HPLC of isolated fraction of radish extract. HPLC, high-performance liquid chromatography.

Oxidative Stress Inhibitory Effect of Radish Extracts Separated by Solvent Partition
The bold fraction exhibited the highest positive activity.
Oxidative Stress Inhibitory Effect of Radish Extracts Separated by Silica-Gel Open Column Chromatography
The bold fraction exhibited the highest positive activity.
Protective Effect of Radish Extracts Separated by Thin-Layer Chromatography Against Oxidative Stress
The bold fraction exhibited the highest positive activity.
Effect of oleamide on memory impairment
To confirm the protective effect of oleamide against memory impairment, a Y-maze test and passive avoidance test were used. Compared with the control group, the Aβ-injected group exhibited significantly lower (decrease of 21.48%) spontaneous alternation behavior. However, this Aβ-induced decrease was significantly improved by pretreatment with oleamide compared with the control group (Fig. 11). No significant differences in the total number of arm entries were observed among the groups (data not shown). Learning and memory impairment was evaluated using a passive avoidance test (Fig. 12). The step-through latency of Aβ-treated group was shorter (43 sec decrease) than that of the control group. Treatment with an oleamide-supplemented diet reversed Aβ-induced impairment.
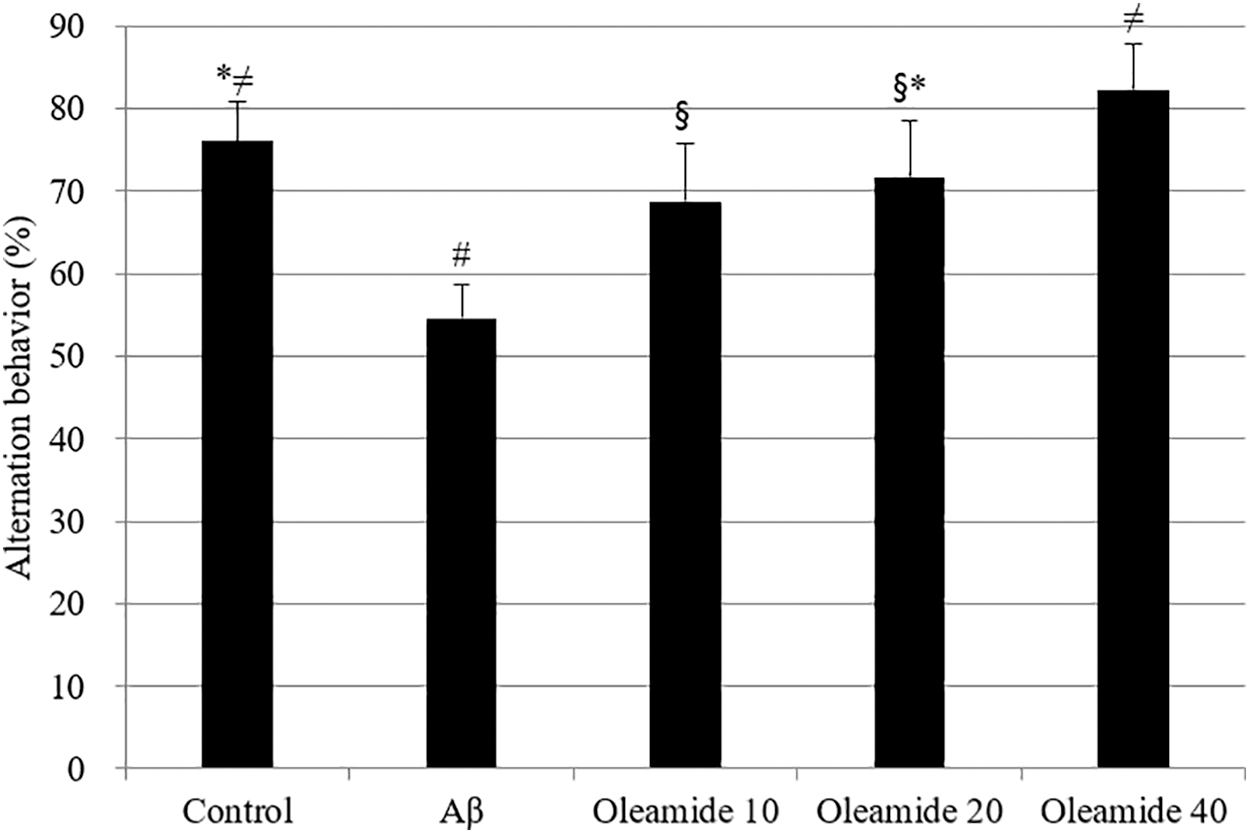
The effect of oleamide on spontaneous alternation behavior in Y-maze test. Controls were injected with Aβ 1–42. Aβ was injected with 410 pmol of Aβ 1–42 per mouse. Sample groups (oleamide 10, 20, and 40) were injected with Aβ 1–42 followed by intake of food with radish extract (10, 20, and 40 mg/kg per day, respectively). Spontaneous alternation behaviors were measured during 8 min. *, #, §, ≠ Represent statistical differences between groups (P < .05). Values indicate the mean ± SD (n = 8).
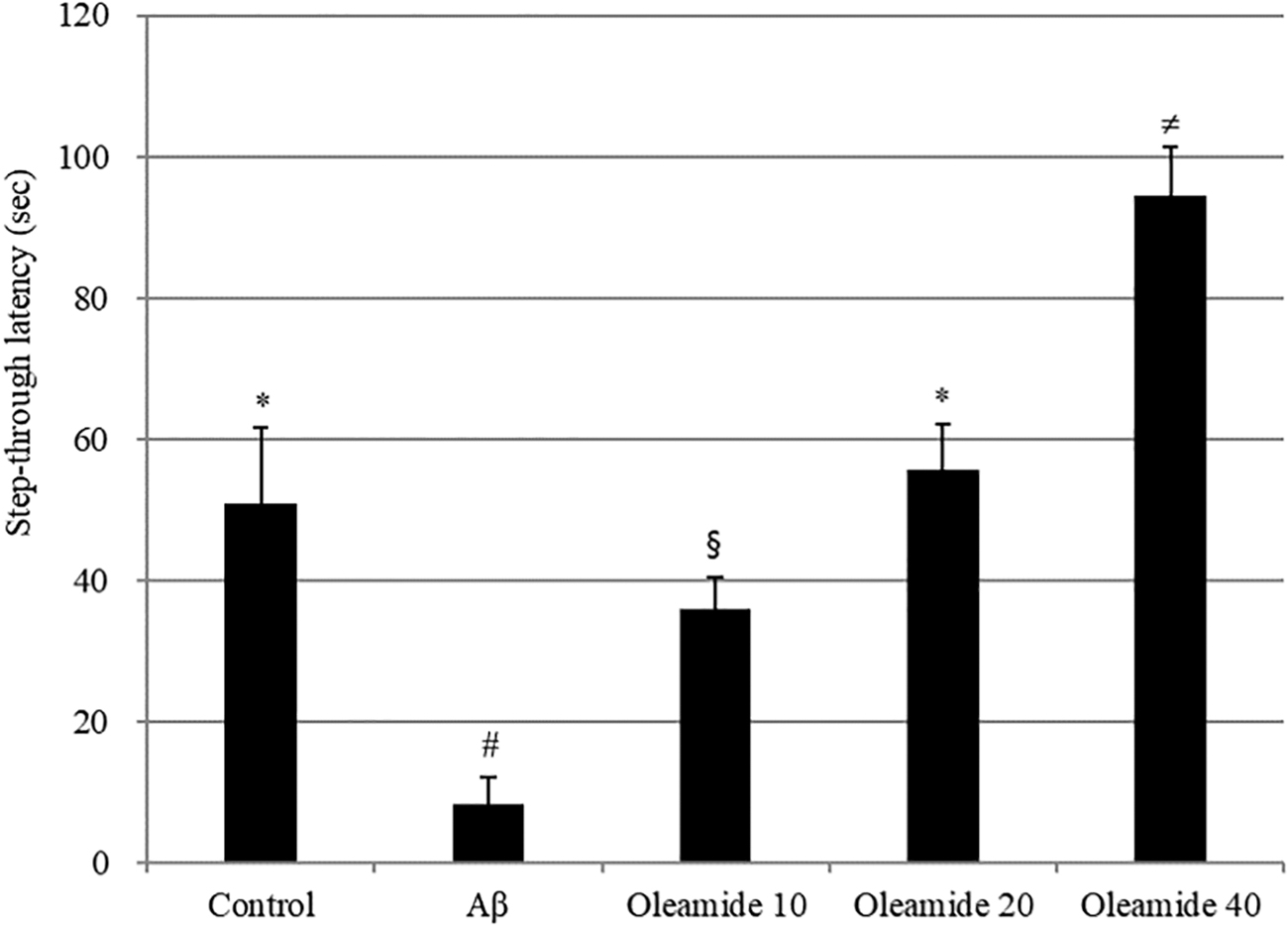
The effect of the oleamide on step-through latency in the passive avoidance test. Controls were injected with Aβ 1–42. Aβ was injected with 410 pmol of Aβ 1–42 per mouse. Sample groups (oleamide 10, 20, and 40) were injected with Aβ 1–42 followed by intake of food with radish extract (10, 20, and 40 mg/kg per day, respectively). The step-through latency was determined during 300 sec. *, #, §, ≠ Represent statistical differences between groups (P < .05). Values indicate the mean ± SD (n = 8).
Discussion
The amyloid plaques and neurofibrillary tangles in the brain are regarded as some of the main features of AD. Another reason is the loss of connections between neurons in the brain. AD disrupts communication among neurons, resulting in loss of function and cell death. Aβ-induced free radical oxidative stress is a factor in the neurodegeneration detected in the AD brain. Aβ may be key to the pathogenesis of AD. The AD brain is under strong oxidative stress, demonstrated by increased protein oxidation, lipid peroxidation, free radical formation, and DNA/RNA oxidation. 5 Aβ has been shown to be a source of free radicals that can initiate neurodegenerative disorders. Intracellular accumulation of ROS is the key cause of Aβ-induced cytotoxicity, ultimately leading to cell death. 5 In this study, we used PC12 cells and mice to check the protective effect of the radish extract against neurotoxicity.
The scavenging activity of the radish extract was confirmed using the DPPH radical scavenging assay (Fig. 1A). The DPPH assay is one of several analytical methods that have been developed to measure antioxidant capacity, mostly in plant and food extracts. 16 A linear correlation was observed between free radical scavenging capacity and extract concentration. In addition, ABTS assays have been widely used to evaluate the antioxidant capacities of natural compounds. ABTS assays are mainly related to the hydrogen-donating or proton radical-scavenging abilities of the target compounds. 17 Radish extracts showed a proportional decrease in radicals with various concentrations of extracts using the ABTS method (Fig. 1B). The extracts decreased the ABTS free radicals, which are related to the scavenging capacity of extracts and vitamin C. Radish extracts demonstrated around 1.5 times higher inhibition compared with vitamin C (Fig. 1C).
ROS can be the cause of cell death in PC12 cells, and such effects can be blocked or delayed by various natural compounds. 18,19 ROS are a group of molecules produced in the cell through metabolism of oxygen. Endogenous ROS such as H2O2 have been recognized as harmful molecules. 20 DNA laddering analysis is normally used as a method to identify protective systems associated with antioxidants against H2O2-induced cell death. 21,22 In the present study, the radish extract reduced DNA laddering resulting from fragmentation. The radish extract pretreatment significantly suppressed H2O2-induced cytotoxicity in PC12 cells. Attenuation of DNA damage confirmed its protective effects.
The effect of dietary intake of the radish extract on behavioral abilities was observed using a mouse model following ICV injection of Aβ 1–42. The ICV injection of Aβ 1–42 has been previously shown to induce memory deficits. 23 Next, memory and learning abilities were evaluated in a Y-maze test and a passive avoidance test. The Y-maze is widely used to test working memory in behavioral science. For this purpose, spontaneous alternation behavior is monitored, and an increased percentage of spontaneous alternation is regarded as an indication of enhanced working memory. The results demonstrate that Aβ induced memory impairments in spontaneous alternation behavior in a Y-maze (Fig. 3) and suggest that dietary radish extract had a protective effect against learning and memory deficits. Mice treated with the radish extract exhibited diminished Aβ 1–24-induced impairment of passive avoidance performance in a dose-dependent manner (Fig. 4). Thus, the radish extract displayed a significant antiamnestic effect in the mouse model. According to the oxidative stress theory of AD, Aβ 1–42 creates free radicals in the cell membrane, resulting in lipid peroxidation, loss of antioxidant defense enzymes, and cellular dysfunction. 24 Catalase is a very important enzyme in protecting the cell from oxidative damage by ROS. It catalyzes the decomposition of hydrogen peroxide to water and oxygen. Lack or malfunction of catalase is suggested to be related to the pathogenesis of many age-associated degenerative diseases. 25 To determine the Aβ-induced damage in the mouse brains, the activity of the antioxidant enzyme catalase was observed. In previous studies, the MDA level was increased and catalase levels were reduced in Aβ mice model. 24 All groups administered radish extracts displayed meaningfully reduced levels of lipid peroxidation in the brain tissues, which possibly explains the positive effects of the extract against cognitive deficits induced via ICV injection of Aβ 1–42. This result suggests that the extract mitigated Aβ 1–42-induced memory impairment in a mouse model and that a diet containing the radish extract protected mouse brain cells against Aβ aggregation, lipid peroxidation, and loss of antioxidant defense enzymes. As shown in Figure 8, the radish extracts inhibited neuronal abnormalities and affected neurotransmitter concentrations.
Many natural compounds can be used for chemoprevention. In addition, some compounds prevent neurotoxicity through their antioxidative properties. 26 –28 In this study, oleamide was identified using several stages of purification to derive the active substance of the radish extract. Oleamide is an amide of the fatty acid, oleic acid. Oleamide is naturally produced in the body of animals. It accumulates in the cerebrospinal fluid during sleeplessness and induces sleep in animals. It was studied as a potential treatment for mood and sleep disorders as well as cannabinoid-regulated depression. 29,30 The mechanism of action of the sleep-inducing effects of oleamide is an area of current research. It is likely that oleamide interacts with neurotransmitter systems, including choline acetyltransferase. In the previous research, oleamide was isolated from Ziziphus jujuba extract. We used a scopolamine-induced animal model as an experiment for cholinergic impairment. Oleamide diet (10–40 mg/kg body weight) protected against scopolamine-induced cholinergic toxicity in a mouse model. 31,32 Unlike our previous study, the radish extract showed strong antiamnestic properties by decreasing oxidative stress in the brain. This protective effect might be due to the antioxidative action of oleamide. The effects of dietary administration of this active compound on behavioral abilities were studied using an animal model. A Y-maze test was used to evaluate memory, learning ability, and spontaneous alternation behavior, which is regarded as a measure of spatial memory. As shown in the behavior test, Aβ 1-24-induced cognitive dysfunction was attenuated in mice treated with the oleamide diet. Thus, the oleamide diet mediated a significant antiamnesic effect in the mouse model. After behavioral test, hepatotoxicity was investigated using a serum transaminase reagent kit. The results of serum transaminases did not reveal significant differences among different groups, and all groups had normal levels of aspartate aminotransferase and alanine aminotransferase.
In conclusion, the radish extract effectively reversed oxidative damage in both in vitro and in vivo models of AD. Biochemical experiments using brain tissues showed lowered Aβ aggregation and oxidative stress levels. The radish extract containing the active compound oleamide inhibited cell death by reducing oxidative stress. These results suggest that the radish extract and oleamide might prevent AD by regulating Aβ-induced cytotoxicity.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by a Korea University Grant.