
Abstract
Sparassis crispa (SC), known as cauliflower mushroom, possesses a wide variety of health-promoting properties and a high content of β-glucans. Its nutritional properties are enhanced by fermentation. In this study, we examined the efficacy of Lactobacillus-fermented (lacto-fermented) SC against obesity using a zebrafish model. We first fermented SC by Lactobacillus paracasei, denoted as lacto-fermented SC (L-SC), for 48 h and then orally administered SC or L-SC to diet-induced obese zebrafish for 4 weeks. Results demonstrated that the L-SC group (20 μg/gBW/day) significantly (P < .01) suppressed body weight gain and ameliorated lipid accumulation in liver tissues, whereas SC did not exhibit antiobesity effects. We further performed expression analysis of genes related to lipid metabolism in the liver and visceral adipose tissues (VAT) in L-SC-administered fish. In liver tissues, L-SC upregulated (P < .05) expression of genes involved in peroxisome proliferator-activated receptor alpha pathways, suggesting that the lipid-lowering property of L-SC is caused by activation of beta-oxidation. In VAT, L-SC did not show significant changes between the experimental groups. No difference was observed between the β-glucan contents of SC (43.8 g/100 g) and L-SC (44.3 g/100 g); however, β-glucan levels in the hot-water extracts increased 20-fold in L-SC (37.2 g/100 g) compared with those in SC (1.8 g/100 g). In summary, lacto-fermentation of SC enhances its lipid-lowering property and can prevent hepatic steatosis through activation of beta-oxidation. Dietary supplementation of fermented L-SC as a functional food may be suitable for obesity prevention and reduction in the prevalence of obesity-related diseases.
Introduction
According to the global obesity survey conducted in 2013, an estimated 2.1 billion people were reported to be overweight or obese. 1 Obesity is associated with several adverse health consequences, including dyslipidemia, type 2 diabetes mellitus, cardiovascular diseases, and some types of cancers. Diet and exercise regimens are the mainstays of obesity treatment. If these approaches are ineffective, antiobesity medicines that reduce appetite or inhibit fat absorption may be taken. However, most of these treatments are associated with side effects, including high blood pressure, restlessness, insomnia, and drug addiction. 2 Thus, many classes of natural products have been studied for their potential to treat obesity with fewer and milder side effects.
Mushrooms have been extensively used as foods, nutraceuticals, and medicines since ancient times worldwide. They contain various bioactive compounds such as primary metabolites that are known to reduce oxidative stress, inhibit cancer cell proliferation, antibiotics, and anti-inflammation, and suppress obesity. 3 Of these, Sparassis crispa (SC), also known as cauliflower fungus, is one of the promising mushrooms that can potentially prevent broad-spectrum human diseases. Two antifungal compounds in SC were first identified in 19934; furthermore, SC was shown to have broad-spectrum therapeutic or preventive properties, 5 –7 including antiobesity functionality. 8
Fermentation is a natural process through which bacteria and yeast act on carbohydrates such as starch and sugars. A number of health benefits are associated with fermentation, and the fermented forms, termed probiotic fermented foods, are often more nutritious than their unfermented forms. In particular, Lactobacillus-fermented (lacto-fermentation) foods exhibit various health-promoting functions, for example, fermented milk exhibits hypotensive function, 9 prevention against rotavirus-associated diarrhea, 10 and reduction of serum triglyceride. 11
Lacto-fermentation is not only restricted to milk and dairy products such as yogurt but is also applicable to vegetables and fruits, such as sauerkraut, and is used for preserving foods by production of lactic acid. Furthermore, some kinds of mushrooms display enhanced biological effectivity as prebiotics, postlacto-fermentation. 12,13 It is possible that hydrolytic enzymes produced by lactobacilli during fermentation break down the cell walls and release intracellular β-glucans or other bioactive compounds, thus increasing bioavailability. Thus, we hypothesized that lacto-fermentation enhances therapeutic properties of SC as a probiotic functional food.
Zebrafish (Danio rerio) are small teleost fish that are increasingly used to model human obesity and obesity-related diseases, including visceral adiposity, 14 hepatic steatosis, 15 and type 2 diabetes. 16 The organs that show symptoms of adiposity, such as the liver and visceral adipose tissues (VAT), are similar to those found in humans, and the pathophysiological pathways of visceral adiposity and hepatic steatosis are common to those found in obese humans. 17 Thus, the diet-induced obesity (DIO) zebrafish model faithfully reproduces the characteristics associated with human obesity.
To enhance bioactivities of SC, we prepared lacto-fermented SC (L-SC) and evaluated it in DIO zebrafish. We further investigated the mechanisms by which L-SC exerts its antiobesity effects by performing a gene expression analysis.
Materials and Methods
Zebrafish
Wild-type zebrafish (AB line; the Zebrafish International Resource Center, Eugene, OR, USA) were maintained in our facility according to standard operational guidelines, complying with international guidelines. Ethical approval from the local Institutional Animal Care and Use Committee was not sought as the Japanese animal welfare regulation, Act on Welfare and Management of Animals (Ministry of the Environment, Government of Japan), does not mandate protection of fish.
Preparation of L-SC
SC and L-SC were provided by Katsuragi Sangyo (Wakayama, Japan). For lacto-fermentation of SC, we used the Lactobacillus paracasei NITE-P-01960 strain, which lives on the surface of SC in nature. SC dried powder was suspended in water containing 0.5% glucose and autoclaved. The SC mixture was then inoculated with the Lactobacillus bacteria and incubated at 30°C for 48 h. The cultured medium was dried by heating at 60°C to obtain L-SC. The major nutritional compositions (Supplementary Table S1), amino acid components (Supplementary Table S2), and β-glucans were measured by Japan Food Research Laboratories (Tokyo, Japan). For the SC or L-SC extracts, 20 g of SC or L-SC was extracted with 1 L of hot water and concentrated to 150 mL by continuous heating, followed by measurement of β-glucan contents. To perform the zebrafish obesogenic test (ZOT), L-SC and SC hot-water extracts were diluted using zebrafish breeding water (0.3 × Danieau's solution).
Feeding experiment
For oral administration of L-SC or SC to zebrafish, we prepared gluten-based fish foods, as we have previously described. 18 The experimental schedule was similar to that in our previous studies. 19 In the first 4 weeks, 3-month-old female zebrafish were divided into 15 fish per 2-L tank for dietary restriction and fed once a day (4 mg/fish/day) with the regular diet. Zebrafish were divided into five groups; normal feeding (NF), overfeeding (OF), OF + L-SC (10 μg/g body weight [BW]), OF + L-SC (20 μg/g BW), and OF + L-SC (60 μg/g BW). The details for standard zebrafish feed are shown in Supplementary Table S3. For the first 2 weeks, gluten granules (6 mg per fish) were fed three times a day, followed by Artemia (6 mg per fish) ∼30 min later. For the next 4 weeks, gluten granules containing L-SC or SC were fed (6 mg per fish), three times a day, 30 min before the Artemia feed. Leftover food was removed daily to avoid water pollution. Zebrafish body weights and Artemia feeding volumes were measured weekly, as previously reported. 19
Measurement of plasma TG and FBG in zebrafish
At the end of the feeding experiment, zebrafish were starved overnight to obtain fasting blood glucose (FBG) measurements as previously reported. 20 FBG was measured using a handheld glucometer (Glutest Neo Super; Sanwa Kagaku, Nagoya, Japan). Plasma triacylglycerol (TG) levels were measured using Wako L-type TG assay kits (Wako Pure Chemicals, Osaka, Japan) according to the manufacturer's protocol.
Oil Red O staining
Liver tissues were collected from zebrafish by surgical manipulation. The preparation of liver sections and Oil Red O (ORO) staining were performed as described previously. 21 The ORO-positive area was quantified using ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Total RNA isolation, cDNA synthesis, and real-time qPCR
After bead homogenization of liver and mesenteric adipose tissues, total RNA purification and cDNA synthesis were performed as we previously described. 19 qPCR was conducted on cDNA samples using a Power SYBR Green Master Mix (Applied Biosystems, Foster City, CA, USA) and the ABI StepOnePlus Real-Time PCR System (Applied Biosystems) according to the manufacturer's instructions. Primer sequences are listed in Supplementary Table S4.
Zebrafish obesogenic test
We performed a 1-week ZOT using the previously reported 22 method with some minor modifications. Zebrafish larvae, ∼10 mm in length, were selected and placed in a six-well plate, with each well containing 5 mL 0.3 × Danieau's solution and five larvae. On the first day of the ZOT, the larvae were fed 0.1% hard-boiled chicken egg yolk as the high-fat diet. On the second day, the larvae in the control group were starved and the experimental groups were treated with the SC or L-SC hot-water extracts for the next 3 days. The volume of VAT was measured before and after SC or L-SC treatment using Nile Red staining. 23 Fluorescent images were captured using a BZ-X710 fluorescence microscope (Keyence, Tokyo, Japan). The Nile Red-positive intensities were quantified using ImageJ software.
Statistical analyses
Data are expressed as means ± standard errors (SE). Statistical analyses were performed using Student's t-tests or one-way analysis of variance with the Bonferroni–Dunn multiple comparison procedure, depending on the number of comparisons (GraphPad Prism, version 7; GraphPad Software, San Diego, CA, USA). A P-value of less than .05 was considered statistically significant.
Results
L-SC ameliorates obesity in DIO zebrafish
We first overfed zebrafish and then administered L-SC or SC for 4 weeks to evaluate their antiobesity activities. As shown in Figure 1A, the OF group fish significantly (P < .01) increased in body weight compared with NF fish from week 1. The L-SC (20 μg/BW) group showed a significantly (P < .05) suppressed body weight gain compared with OF fish from week 5, 3 weeks after the start of L-SC administration (0.91 ± 0.03 g in the OF group vs. 0.74 ± 0.05 g in the OF + L-SC 20 μg/g BW group at week 6). Food intake, which was measured as the average number of Artemia consumed by one fish, was not significantly altered between OF and OF + L-SC groups (Supplementary Fig. S1). In the SC group, even at the end of the feeding experiment (week 6), no difference was observed between the OF fish with or without SC administration (Supplementary Fig. S2). Computed tomography analyses showed a significant (P < .01) increase of VAT in OF fish compared with NF fish. L-SC showed a tendency (P < .2) to reduce VAT at week 6 (2.25 ± 0.37 mm3 in the OF group vs. 1.83 ± 022 mm3 in the OF + L-SC 10 μg/g BW group; Fig. 1B), whereas SC administration did not have any effect (Supplementary Fig. S3). Corresponding to the improvement in visceral adiposity, L-SC significantly suppressed (P < .05) plasma TG [551.56 ± 68.28 mg/dL in the OF group vs. 350.13 ± 87.50 mg/dL in the OF + L-SC (10 μg/gBW) group; Fig. 1C] at week 6. SC did not affect plasma TG (Supplementary Fig. S4), consistent with the results of body weight (Supplementary Fig. S2) and CT analyses (Supplementary Fig. S3). Hyperlipidemia is accompanied by hepatic lipid deposition in obese zebrafish, 24 and ORO staining revealed that the L-SC (20 μg/gBW) group showed lower lipid accumulation in liver tissues than that in the OF group (Fig. 1D, E). Compared with OF alone, L-SC (20 μg/gBW) administration with OF showed a tendency (P = .3) to reduce FBG levels (Supplementary Fig. S5). Contrary to this finding, SC did not affect blood glucose (Supplementary Fig. S6).

L-SC ameliorates obese phenotypes in zebrafish.
L-SC affects expression of genes associated with beta-oxidation in liver and adipose tissues
To determine whether the lipid-lowering effect of L-SC depends on facilitation of lipolysis (beta-oxidation) or inhibition of lipogenesis, we analyzed gene expression profiles in the liver tissues and VAT of DIO zebrafish. In the liver, the expression of genes associated with beta-oxidation, such as peroxisome proliferator-activated receptor alpha b (pparab), an ortholog for human PPARA, was significantly (P < .05) upregulated by L-SC administration (Fig. 2A; 10 μg/gBW L-SC). Additionally, its target gene, acyl-CoA dehydrogenase, C-4 to C-12 straight chain (acadm), was significantly (P < .05) upregulated by L-SC administration (Fig. 2B). Another pparab target gene, acyl-CoA oxidase 1, palmitoyl (acox1), was not affected by L-SC (Fig. 2C; P < .2). With respect to lipogenic genes, expression levels of fatty acid synthase (fasn) and acetyl-CoA carboxylase alpha (acc) were not affected by L-SC administration (Fig. 2D, E). These results indicate that the lipid-lowering effect of L-SC is affected by induction of genes associated with beta-oxidation. In VAT, no significant differences were observed in the associated gene expression profiles among the experimental groups (Supplementary Figs. S7–S11).

L-SC-induced gene expression changes in liver tissues.
Lipid-lowering effects of hot-water L-SC extracts
Because SC is well known for its high levels of 1, 3-β-D-glucans, 25 we measured the β-glucan levels of SC and L-SC and found no difference in the total content of β-glucans between them (Supplementary Table S1; L-SC: 44.3 g/100 g, SC: 43.8 g/100 g). To evaluate the β-glucan release from SC and L-SC powders, we performed hot-water extraction and found that the L-SC extract contained 20 times more β-glucan than SC extracts treated in the same way (Table 1; 37.2 g/100 g in L-SC vs. 1.8 g/100 g in SC). To demonstrate that the high β-glucan content of L-SC extracts exerts a lipid-lowering effect in animal models, we performed a ZOT as previously reported (Fig. 3A) 22,23 ; L-SC hot-water extracts, which contain about 200 μg/mL β-glucan, significantly (P < .05) suppressed increases in VAT, while the SC extract did not (P = .16; Fig. 3B, C).
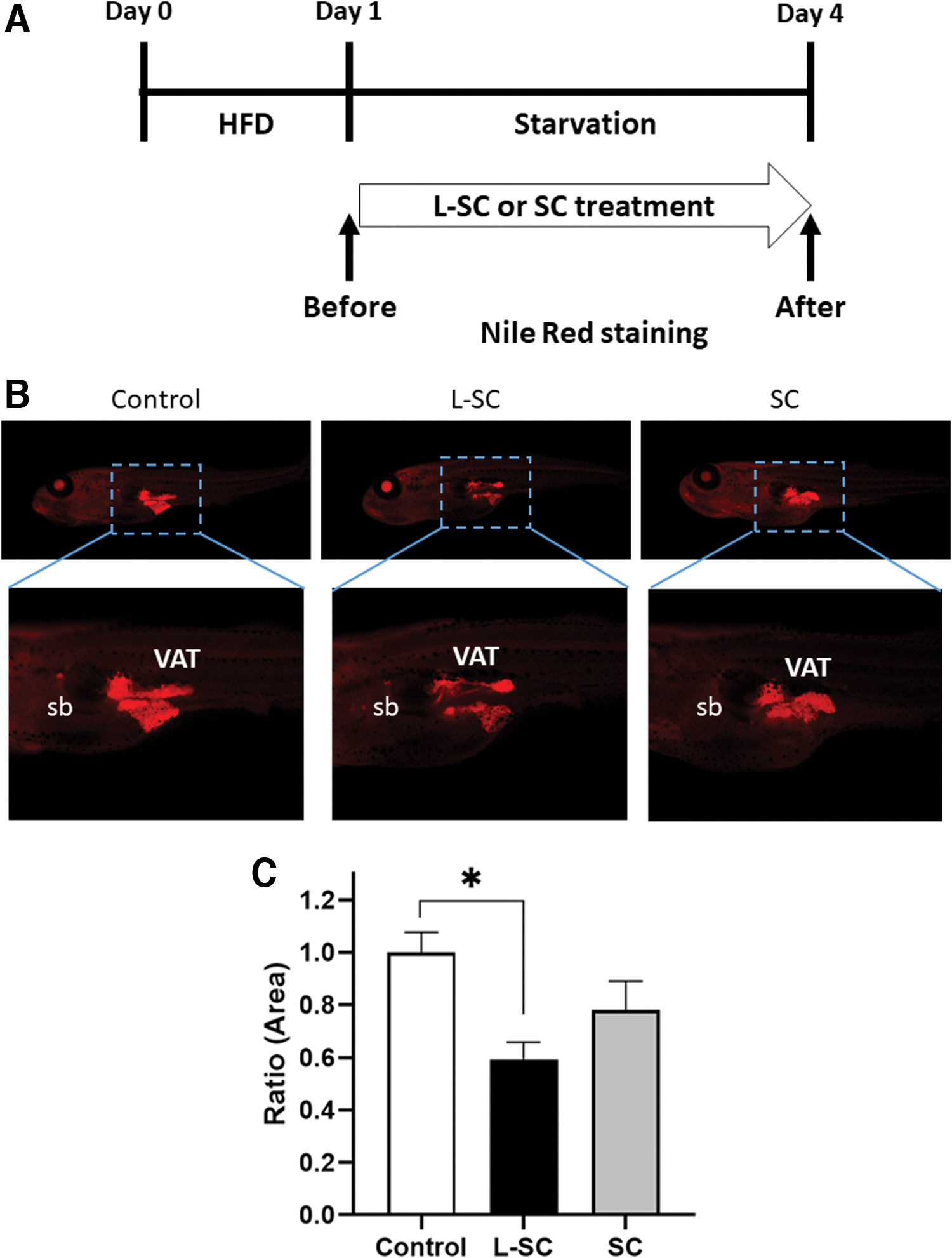
Evaluation of the obesogenic properties of L-SC and SC hot-water extracts in zebrafish.
Content of β-Glucans
HWE, hot-water extract.
Discussion
Lacto-fermentation enhances antiobesity properties of S. crispa
In this study, we demonstrated that L-SC showed strong antiobesity effects compared with those by SC. L. paracasei is a gram-positive, facultative heterofermentative species of lactic acid bacteria commonly used in dairy product fermentation and probiotics. L. paracasei and its products (lacto-fermentation) improve the conditions associated with obesity and its related diseases. 26,27 To our knowledge, this is the first study elucidating antiobesity properties of lacto-fermented mushrooms. Specifically, the L. paracasei NITE-P-01960 (Japanese patent: JP2016123382) strain that thrives on the surface of SC in nature would be suitable for inducing fermentation of SC. In 2018, Takeyama et al. reported that oral administration of SC reduced body fat mass by enhancement of energy expenditure and suppression of lipogenesis in obese rats 8 ; however, this could not be confirmed in our zebrafish model (Supplementary Figs. S2–S4). The authors revealed that the highest dose (4% food volume = ∼3 mg/g BW, more than 50 times higher than our protocol) of SC administration increased fecal fatty acid content, indicating that SC impairs lipid absorption in the rat intestine, which could not be detected in our zebrafish models probably because of the lower volume of SC administered in our study. L-SC significantly suppressed plasma TG (Fig. 1D) and liver lipids (Fig. 1E, F) (P < .05), but not VAT (Fig. 1C, P < .2). However, L-SC suppressed body weight gain in the OF group (Fig. 1A); the primary target organ of L-SC was the liver rather than VAT.
In our study, treatment with high doses of L-SC (60 mg/kg BW) did not exert any significant effects on obese phenotypes. Because SC contains highly active biological and pharmacological compounds, with the exception of β-glucans, 28 we hypothesized that high doses of L-SC or SC may induce some toxic effects in zebrafish. To validate this, we performed a zebrafish embryo acute toxicity test using L-SC or SC suspensions. As shown in Supplementary Figure S12, 60 μg/mL L-SC-treated embryos still hatched at 72 hpf, while SC-treated embryos did not. In addition, there was no effect on survival rates with embryos treated with 250 μg/mL L-SC or SC. These data support our hypothesis that L-SC is slightly toxic to zebrafish embryos.
Lipid-lowering mechanism of L-SC
Lipid metabolism in the zebrafish liver is very similar to lipid synthesis and fatty acid beta-oxidation in humans. 29 Recent studies have explored zebrafish homologs of human genes involved in lipid metabolism; for example, several transgenic and mutant zebrafish with hepatosteatosis exhibit upregulation of key lipogenic genes, such as acc and fasn, which show mechanisms similar to those observed in patients. 30 In this study, L-SC upregulated the PPARα-driven beta-oxidation pathway in the liver tissue (Fig. 2), and liver PPARα is crucial for whole-body fatty acid homeostasis with protective function against nonalcoholic fatty liver disease. 31 In fact, our feeding protocol was focused on lipolysis (or adipolysis) considering the 2-week pre-OF period. Furthermore, our findings from gene expression analysis and phenotypic changes were consistent with those of L-SC-induced beta-oxidation activation to promote lipid clearance. In VAT, we did not find significant changes in gene expression (Supplementary Figs. S7–S11), consistent with the CT results (Fig. 1C). These results indicate that the main therapeutic target organ of L-SC could be the liver and not VAT in our zebrafish model.
β-glucans, possible antiobesity constituents in L-SC
In general, mushrooms contain β-glucans that protect against cancer, microbial infections, hyperlipidemia, and diabetes. 32 In this study, we were able to confirm that both L-SC and SC contain high levels of β-glucans and that hot-water extraction increased β-glucan isolation in L-SC extracts when compared with SC (Supplementary Table S1). In addition, the L-SC hot-water extract significantly (P < .05) suppressed lipid accumulation in rapid zebrafish testing (Fig. 3B, C), while SC extracts did not. This suggests that β-glucans are one of the compounds that could be used to improve obesity treatments. These results also indicate that the bioavailability of β-glucans in SC extracts could be drastically improved by lacto-fermentation, which enhances the antiobesity properties of L-SC. We speculate that lacto-fermentation hydrolyzed the cell wall of SC, releasing the intracellular β-glucans into the hot water. In addition, oat β-glucan reduces lipid accumulation in high-fat diet-fed mice with upregulation of the PPARα pathway in the liver, 33 which is consistent with our results. There is a possibility that abundance of lactobacilli on the L-SC increases the total quantity of available β-glucans. However, this increase would be negligible in comparison with the amount found in native SC.
In this study, we showed for the first time (to the best of our knowledge) that lacto-fermentation of SC enhances its antiobesity effects, particularly by lipid-lowering activity in liver tissues, with upregulation of the PPARα-driven beta-oxidation pathway. We expect that L-SC will aid people suffering from obesity and hepatosteatosis and that lacto-fermentation would be applied to other mushroom species to enhance their health-promoting properties.
Footnotes
Acknowledgments
The authors thank Ms. Taguchi and Ms. Kato for their secretarial assistance and Ms. Inoue for zebrafish care and maintenance. The authors also thank Editage for English language editing.
Author Disclosure Statement
Nobuo Matsuura is the CEO of Bankyo Pharmaceutical Co., Ltd., a chemical company. Other authors declare no conflicts of interest directly relevant to the content of this article.
Funding Information
This work was supported by the Japan Science and Technology Agency, Grant Number VP29117938605.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Figure S10
Supplementary Figure S11
Supplementary Figure S12
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.