
Abstract
Myo-inositol supplementation may reduce insulin resistance (IR) with few serious side effects in patients with polycystic ovary syndrome (PCOS). To explore the mechanism of this action in an animal model, a PCOS-IR rat model was generated. Enzyme-linked immunosorbent assay was used to assess changes in ovulation function during treatment with a myo-inositol supplement, and Western blotting, real-time polymerase chain reaction, and immunohistochemistry were performed to investigate the underlying molecular mechanisms. The results showed that the myo-inositol supplement decreased the homeostatic model assessment of insulin resistance (HOMA-IR) index and significantly decreased the serum levels of luteinizing hormone (LH), LH/follicle-stimulating hormone ratio, and testosterone, while increasing the serum level of estradiol. Upregulation of interleukin 6 (IL-6), phospho-STAT3 (p-STAT3), Mir-21, and Mir-155 and significant downregulation of PPAR-γ and GLUT4 were detected in the untreated PCOS-IR rat model. However, downregulation of IL-6, p-STAT3, miR-21, and miR-155 and significant upregulation of PPAR-γ and GLUT4 were detected with myo-inositol supplementation. Thus, myo-inositol supplementation may reduce Mir-21 and Mir-155 levels by downregulating IL-6 and p-STAT3 and, subsequently, reverse the expression of PPAR-γ and GLUT4, leading to a decreased HOMA-IR index. In conclusion, the identification of an IL-6/p-STAT3/Mir-155/Mir-21/PPAR-γ/GLUT4 system in the PCOS-IR rat model provides insight into the pathogenesis of PCOS and may indicate a possible therapeutic strategy. Amelioration of the basal serum glucose levels and of the HOMA/HOMA-IR index may be achieved by the reversal of the expression of PPAR-γ and GLUT4 through the downregulation of IL-6, p-STAT3, miR-21, and miR-155 with myo-inositol supplementation.
Introduction
Polycystic ovary syndrome (PCOS) is one of the most common female endocrine and metabolic disorders. Although PCOS can affect women of all ages, its prevalence can be as high as 20% in women of reproductive age. 1 Its clinical expression typically includes oligo-ovulation or anovulation, hyperandrogenism, and polycystic ovaries. 2 A diagnosis of PCOS is associated with an increased risk of type 2 diabetes and cardiovascular problems, mainly due to insulin resistance (IR). 3 Moreover, PCOS is frequently accompanied by IR and obesity. IR represents the pathogenic link between metabolic and reproductive disorders in PCOS and plays a central pathological role in 70–80% of women with obesity. 2
Drugs and surgery are the main choices for comprehensive PCOS therapy. The increasing incidence of PCOS has been accompanied by a growth in the number of studies of potential therapies. In the last decade, myo-inositol was introduced for the treatment of PCOS. Myo-inositol is a sugar alcohol and a glucose isomer found in many foods, including grains and fruits. Myo-inositol is a precursor to numerous secondary messengers and was once considered a B-complex vitamin, and a recent review reported that it reduces IR status and improves ovarian function and androgen levels in women with PCOS. 4
However, the mechanism underlying the decrease in IR afforded by myo-inositol in women with PCOS remains unclear. One study found that total phosphoinositide 3-kinase signaling was unaffected in mice with myo-inositol induced expression of the oncogene Kras in the airway epithelium and that interleukin 6 (IL-6)-related pathways, including STAT3 phosphorylation, were largely downregulated. 5 Reduced IL-6 levels may be associated with decreased IR with myo-inositol supplementation.
PCOS is a pro-inflammatory state, and chronic low-grade inflammation plays an essential role in its pathogenesis. 6 Significant increases in the serum levels of inflammatory factors, including IL-6, have been observed in women with PCOS compared with those who do not have the disease. 7 IL-6, which is a major pro-inflammatory cytokine in chronic inflammation, is closely associated with PCOS and IR status and may be the precursor to the development of PCOS and IR and their metabolic consequences. 8 Therefore, these conditions may share identical or similar transduction pathways, and IL-6 may be an early low-grade chronic inflammatory marker and a key molecule in PCOS. It is reported that the IL-6-mediated JAK2/STAT3 signaling pathway plays an important role in PCOS, 9 and STAT3 signaling stimulates the expression of miR-21 and miR-155, 10,11 evaluated level of miR-21 and miR-155 was also found in PCOS patients; however, whether myo-inositol supplementation affects JAK2/STAT3/miR-21/miR-155 signaling pathway remains unclear, because the mechanism of myo-inositol action in the treatment of PCOS has not been studied. In this study, we established a PCOS rat model to explore the role of myo-inositol supplementation in PCOS. By studying the IL-6/STAT3 pathway, we reveal the potential mechanism of action of myo-inositol supplementation as an effective method with fewer side effects for the treatment of patients with PCOS.
Materials and Methods
Animal model
Female Sprague-Dawley rats (3–4 weeks of age, weighing 45–55 g at the beginning of the experiments) were fed with normal chow diet and tap water during the adaptation period and maintained at constant temperature on a 12-h light/12-h dark cycle. After adaptation to the environment for a week, the rats were used for the following experiments to establish PCOS-IR rat model. The PCOS-IR rat model was fed with HFD (high-fat diet) for 8 weeks and were administered with letrozole (HengRui Medicine, Jiangsu, China) at the concentration of 1 mg/kg dissolved in 0.5% CMC by gavage in the last 4 weeks of the feeding regimen at the dose of 1 mg/kg per day as previously reported.
The HFD comprised 36% carbohydrate, 20% protein, 40% fat, and 4% other material (total energy content 530 kcal). 12 Toluidine blue staining was used to monitor the vaginal exfoliative cytology. The presence of keratosis in vaginal epithelial cells for 10 consecutive days constituted validation of the PCOS rat model. This research was approved by the Experimental Animal Ethics Committee of Fujian Provincial Maternity and Children's Hospital, Affiliated Hospital of Fujian Medical University (Fuzhou, China), with approval number 2018-02-17. The PCOS-IR rat model was selected among the confirmed PCOS rats based on a homeostatic model assessment of insulin resistance (HOMA-IR) index of >2.8. 9 The HOMA-IR index was calculated as HOMA-IR = fasting blood glucose (FBG; mmol/L) × fasting insulin (FINS; mU/L)/22.5. The FBG and FINS values were assessed by collecting blood from the orbital vein of each PCOS rat after 12 h fasting.
Grouping and intervention
The female Sprague-Dawley rats were randomly divided into three groups (n = 15 in each group and rats in each group kept in two cages in the same environment), negative control group (chow diet, treated with physiological saline), PCOS-IR group (HFD and letrozole, treated with physiological saline), and PCOS-IR+Myo-inositol (HFD and letrozole, treated with Myo-inositol). Myo-inositol was administered by gavage at a dose of 50 mg/kg of body weight, as reported, 13 in 2 mL of drinking water per day for 4 weeks. Rats in the negative control and PCOS group were administered normal saline in a similar manner for 4 weeks. The rats were allowed free access to food and water only after 2 h of the drug administration. All the animals were closely observed for the initial 2 h. The rats were sacrificed after 4 weeks. The weight and amount of food intake were recorded in rats from all groups.
Toluidine blue staining
Toluidine blue staining was used to monitor the changes in vaginal exfoliative cytology in rats. An aseptic cotton swab soaked in physiological saline was used to sweep the rat's vaginal wall clockwise. The swab was then smeared on a glass slide in the same direction. Cells were fixed in 4% paraformaldehyde for 15 min, and the vaginal smears were stained with toluidine blue according to the manufacturer's protocol. Images from the negative control, PCOS, and PCOS+myo-inositol group were captured with a microscope imaging system.
Histopathological evaluation
Ovaries were processed with a 10% neutral formalin fixation (24 h) protocol, embedded in paraffin, and sectioned longitudinally and serially at a thickness of 4–5 μm on a microtome. The samples were stained with hematoxylin and eosin to count follicles and observe ovarian architecture.
Enzyme-linked immunosorbent assay
The testing kits for the follicle-stimulating hormone (FSH), luteinizing hormone (LH), testosterone (T), estradiol (E2), and IL-6 were purchased from Cusabio Biotech. Cholesterol and triglyceride levels were evaluated using standard colorimetric kits (Cormay, Łomianki, Poland). According to the manufacturer's protocols, enzyme-linked immunosorbent assay (ELISA) was used to determine the levels of FSH, LH, T, E2, cholesterol, triglycerides, and IL-6 in the sera of blood from the orbital veins of rats from the three groups. Blood samples were stored at −80°C until use.
Electron microscopy examination
Transmission electron microscopy (TEM) was used to observe the structure of organelles in ovarian cells. Half of the right ovary was cut into 1 mm3 strips, which were fixed with 2.5% glutaraldehyde. The ovary samples were rinsed in Millonig buffer (pH 7.4) and postfixed in 1% osmium tetroxide. Samples were then dehydrated in a gradient series of ethanol, placed in propylene oxide, and embedded in araldite (Sigma-Aldrich, St. Louis, MO, USA). A transmission electron microscope was used to analyze the samples. The structure of granulosa cells was assessed, and autophagosomes were detected.
Immunohistochemical staining
The expression of IL-6, STAT3, and phospho-STAT3 (p-STAT3) in ovarian tissues was determined by immunohistochemical (IHC) staining, according to the manufacturer's instructions. We used a sectioning system to obtain ovarian sections. Primary antibodies against STAT3, p-STAT3, and PPAR-γ were purchased from Abcam (Cambridge, United Kingdom). Ovarian sections were incubated with the primary antibody overnight at 4°C, followed by incubation with the secondary antibody at room temperature. Diaminobenzidine was applied to the sections after the last wash. A microscope imaging system was used for sample observation. IHC staining scores were assessed based on the percentage of positively stained cells as follows: samples with negative staining, IHC staining score of 0; samples with <20% positive staining, IHC staining score of 1; samples with 20–50% positive staining, IHC staining score of 2; and samples with >50% positive staining, IHC staining score of 3.
Real-time polymerase chain reaction
The expression of IL-6, PPAR-γ, Mir-21, and Mir-155 was investigated by real-time polymerase chain reaction (PCR). Ovary tissues were placed in liquid nitrogen, and RNA was extracted from ovaries using TRIzol (Takara Bio, Kusatsu, Japan). The Bestar real-time-PCR Kit (DBI Bestar) was used to perform reverse transcription. The Agilent Stratagene Mx3000P Sequence detection system in the presence of the Bestar SYBR Green reagent (DBI Bestar) was used to perform a real-time PCR analysis to quantify gene amplification. The cycling parameters were as follows: 95°C for 2 min, followed by 40 cycles of 94°C for 20 sec, 57°C for 20 sec, and 72°C for 20 sec, and a final incubation at 72°C for 10 min. A melting curve analysis was performed. The relative expression level, defined as fold change, of the target gene was normalized to that of the negative control group. The primer sequences are shown in Table 1.
The Primer Sequences of Interleukin 6 Messenger RNA, PPAR-γ Messenger RNA, Mir-21, and Mir-155
IL-6, interleukin 6; mRNA, messenger RNA.
Immunofluorescence assay
The expression of PPAR-γ and GLUT-4 was investigated by immunofluorescence assay. After incubation with 10% normal goat serum for 1 h at room temperature, to eliminate signals from endogenous peroxidases and nonspecific binding, ovarian tissue sections were incubated with the primary antibody in skim milk supplemented with Triton X-100 in 0.01 M Tris-buffered saline (TBST) at 4°C overnight. After washing with TBST, the sections were incubated with the secondary antibody for 1 h at room temperature. The sections were washed with TBST, covered with DAPI-containing medium, and examined and photographed in duplicate using a confocal microscope before and after DAPI staining. The images were merged using software to determine the location of the target protein. All immunofluorescence experiments were performed by at least two blinded investigators.
Western blot analysis
Solid tissues (40 mg) were shredded and lysed using RIPA buffer that was reconstituted with protease inhibitors (Roche Diagnostics, Basel, Switzerland) and phosphatase inhibitors (Roche Diagnostics), followed by break down of the tissues using a homogenizer. The Pierce BCA Protein Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) was used to measure protein concentrations. The protein samples (20 μg/lane) were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis, transferred onto PVDF membranes (Millipore, Burlington, MA, USA) and incubated with antibodies against IL-6 (1:1000; Antibody Solutions, Santa Clara, CA, USA), STAT3 (1:2000; CST, Danvers, MA, USA), p-STAT3 (1:1000; CST), PPAR-γ (1:1000; CST), GLUT4 (1:1000; CST), or GAPDH (1:10,000; Abcam). HRP-conjugated goat anti-rabbit secondary antibodies or HRP-conjugated goat anti-mouse secondary antibodies (1:20,000; all from BosterBio, Pleasanton, CA, USA) were used, and bands were visualized using enhanced chemiluminescence (Amersham Biosciences, Chalfont St. Giles, United Kingdom).
Statistical analyses
IBM SPSS 20.0 (SPSS, Inc., Chicago, IL, USA) was used to perform statistical analyses. The mean ± standard deviation of the results is reported, as assessed using either one-way analysis of variance with Tukey's post hoc test or the Kruskal–Wallis test with Dunn's post hoc test, which was applied to compare multiple samples. Tests were performed within a 95% confidence interval, and significance was set at P < .05.
Results
Effect of myo-inositol supplementation on food intake, body weight, and estrous cycles in rats
The weekly body-weight changes of the rats in the PCOS group showed a significantly greater weight gain than did animals in the PCOS+myo-inositol and negative control groups over the last 2 weeks of the experiment (Fig. 1A, **P < .01). The average weekly food intake of the PCOS, PCOS+myo-inositol, and negative control group is shown in Figure 1B. The food consumption per rat in the PCOS-IR group during the last 2 weeks of the experiment was significantly greater than that observed in the PCOS+myo-inositol and negative control groups (Fig. 1B, **P < .01).
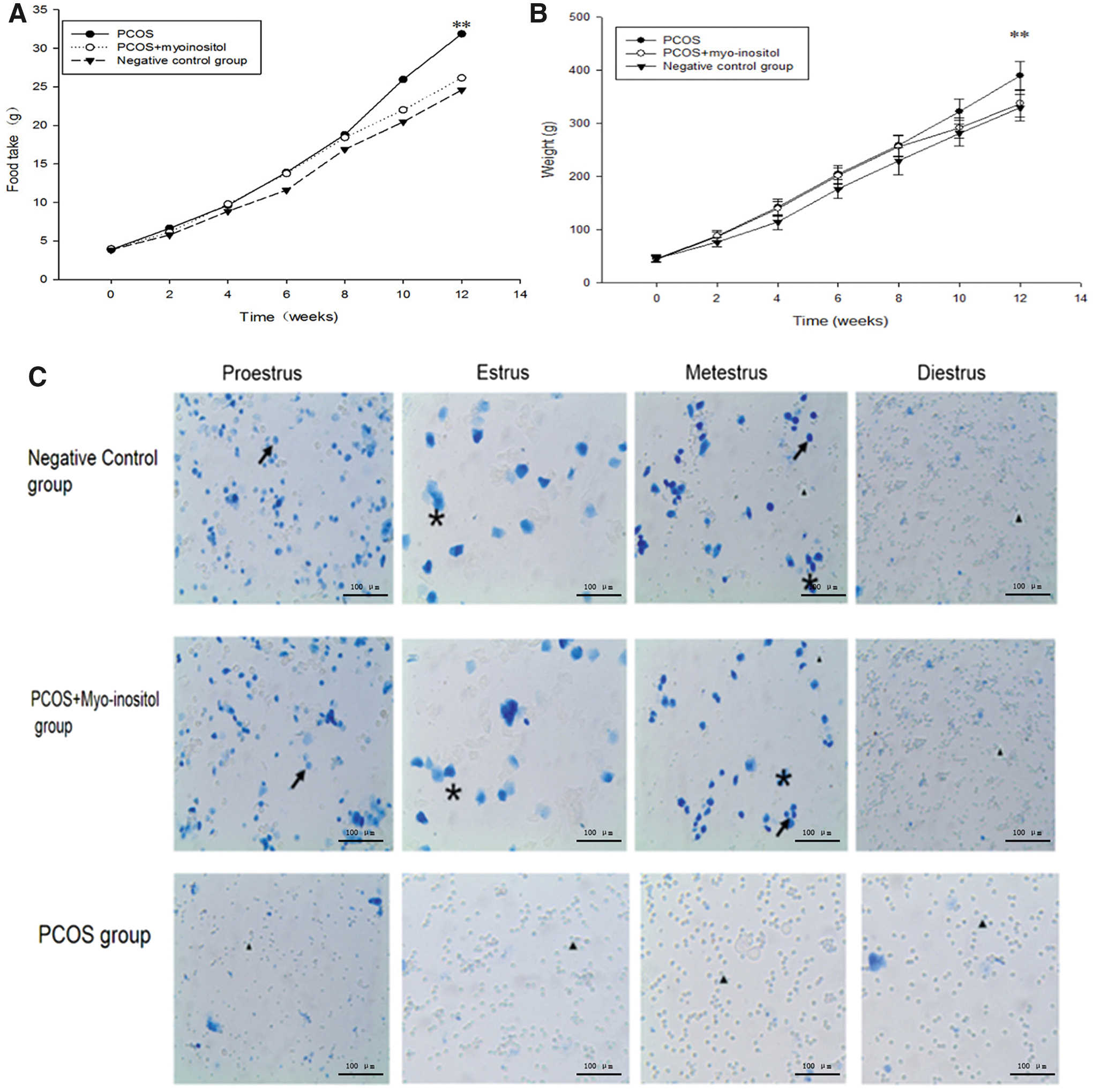
Food intake amount per rat was shown in
The vaginal exfoliated cells of rats were assessed by microscopic analysis. In the PCOS group, a large number of leukocytes and a small amount of mucus were observed in pre-estrus, estrus, postestrus, and anestrus. In contrast, in the PCOS+myo-inositol and negative control group, in pre-estrus, most cells were nucleated epithelial cells; in estrus, a large number of keratin epithelial cells were observed; in postestrus, several types of cells were observed, including nucleated epithelial cells, keratin epithelial cells, and leukocytes; and in anestrus, a large number of leukocytes and a small amount of mucus were observed. The vaginal exfoliation results demonstrated that the PCOS-IR group lost their normal estrus cycle, which was replaced by interestrus (Fig. 1C).
Effect of myo-inositol supplementation on the serum levels of FSH, LH, E2, and T and HOMA-IR index in the PCOS-IR rat model
Sex hormone (FSH, LH, E2, T) levels and the HOMA-IR index were analyzed in rats using ELISA (Fig. 2). The serum levels of LH, T, and the LH/FSH ratio were significantly higher in the PCOS group than in the negative control and PCOS+myo-inositol group (Fig. 2B, C, E, **P < .01). The level of E2 in the PCOS group was slightly lower than that observed in the negative control and PCOS+myo-inositol group (Fig. 2D, **P < .01). Moreover, there was no significant difference in the level of FSH among these groups (Fig. 2A, P = .157). These results suggest that myo-inositol supplementation significantly decreases the serum levels of LH, T, and the LH/FSH ratio, whereas it increases the serum levels of E2 sharply in PCOS-IR rats.

The orbital vein blood of the rats was obtained, the serum levels of FSH
After 12 h of fasting, we measured FBG and FINS, and IR was calculated according to the HOMA-IR method. The HOMA-IR index was 2.15 and 2.27 in the negative control and PCOS+myo-inositol group, respectively, whereas it was 4.22 in the PCOS model group (Fig. 2F, **P < .01). The PCOS rats that exhibited a HOMA-IR index of >2.8 were regarded as having IR. The results showed that a PCOS-IR model was established and that myo-inositol supplementation decreased the HOMA-IR index.
Effect of myo-inositol supplementation on histopathological outcomes and serum levels of cholesterol, triglycerides, and IL-6 in the PCOS-IR rat model
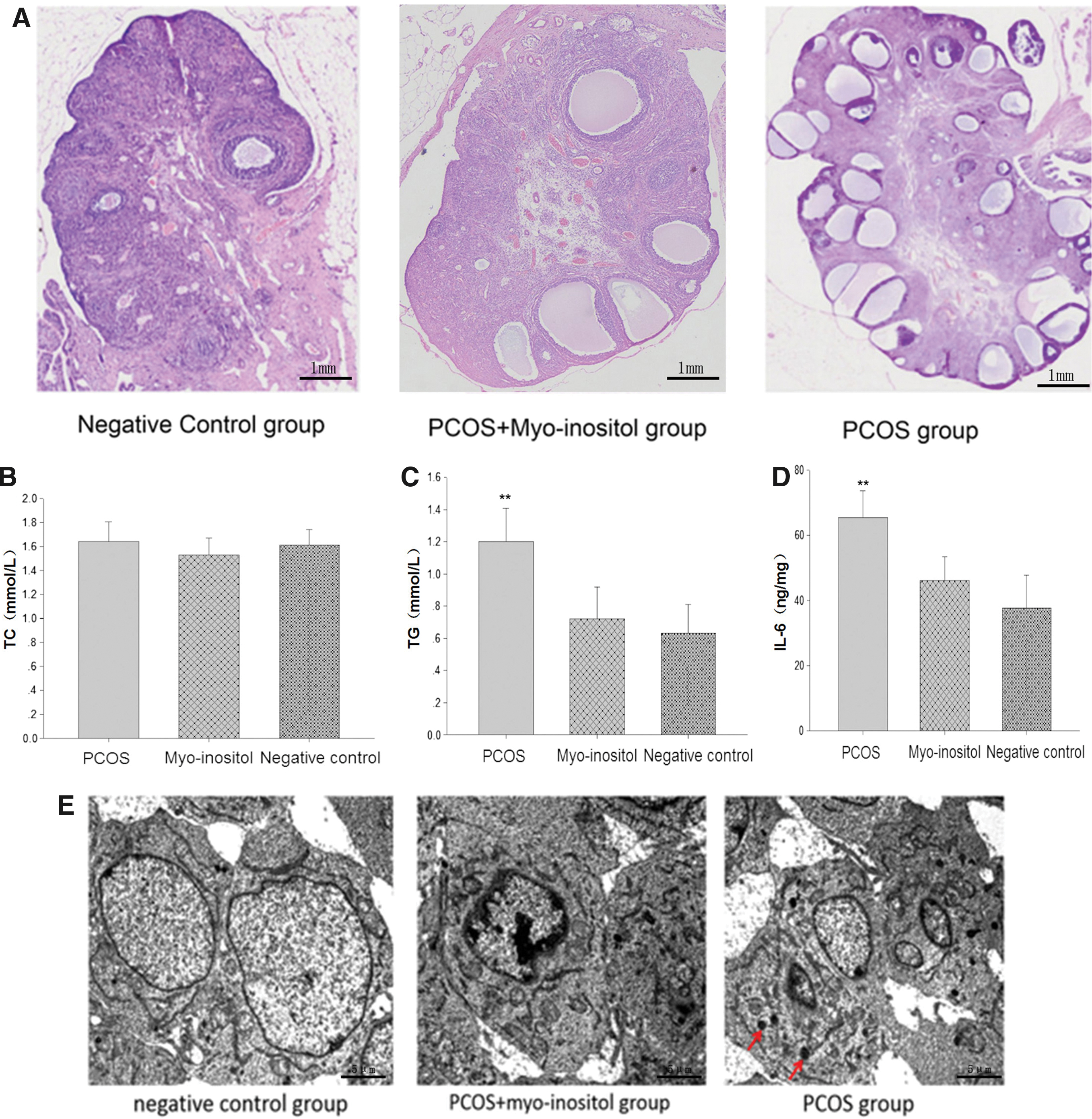
Paraffin sections of rat ovaries were assessed by microscopic analysis. Polycystic changes were observed in the rats of the PCOS group. In contrast, follicular development was normal in the PCOS+myo-inositol and negative control group (Fig. 3A). Observations stemming from the ovarian paraffin sections showed that myo-inositol supplementation can reverse polycystic changes. The levels of cholesterol, triglycerides, and IL-6 in the serum were analyzed by ELISA. The serum level of triglycerides and IL-6 in the PCOS group was slightly higher compared with the negative control and PCOS+myo-inositol group (Fig. 3C, D, **P < .01), whereas no significant differences in the cholesterol serum level were observed (Fig. 3C, D, **P < .01). ELISA revealed that myo-inositol supplementation reduced the level of triglycerides and IL-6 in PCOS model rats (Fig. 3B, P = .216). TEM showed that autophagy would be found in the PCOS group, but not in the PCOS+myo-inositol and negative control group (Fig. 3E).
Effect of myo-inositol supplementation on the expression of IL-6, STAT3, and p-STAT3 in the ovaries of PCOS-IR rats, as assessed by IHC staining
The levels of expression of IL-6, STAT3, and p-STAT3 in the ovarian tissues of rats from the three groups were detected by IHC staining (Fig. 4). The expression of IL-6 and p-STAT3 in the PCOS group was significantly higher compared with the negative control group and PCOS+myo-inositol group (Fig. 4B, **P < .01). In contrast, the expression of STAT3 in the PCOS group was similar to that of the control and PCOS+myo-inositol group (Fig. 4B, P = .314). These data indicate that myo-inositol supplementation reduces the expression of IL-6/p-STAT3 in the ovaries of PCOS rats.

Effect of myo-inositol supplementation on the expression of Mir-155, Mir-21, IL-6, and PPAR-γ messenger RNAs in the ovaries of PCOS-IR rats, as assessed by real-time PCR
To study the molecular mechanism underlying myo-inositol supplementation in the treatment of PCOS rats further, the expression of Mir-155, Mir-21, IL-6, and PPAR-γ messenger RNAs (mRNAs) was analyzed in the ovaries of PCOS-IR rats by real-time PCR. With the exception of the upregulation of the PPAR-γ mRNA, IL-6 mRNA, Mir-155, and Mir-21 tested were significantly downregulated in the negative control group and PCOS+myo-inositol group compared with the PCOS group (Fig. 5, **P < .01).

Effect of myo-inositol supplementation on the expression of IL-6, p-STAT3, and PPAR-γ in the ovaries of PCOS-IR rats
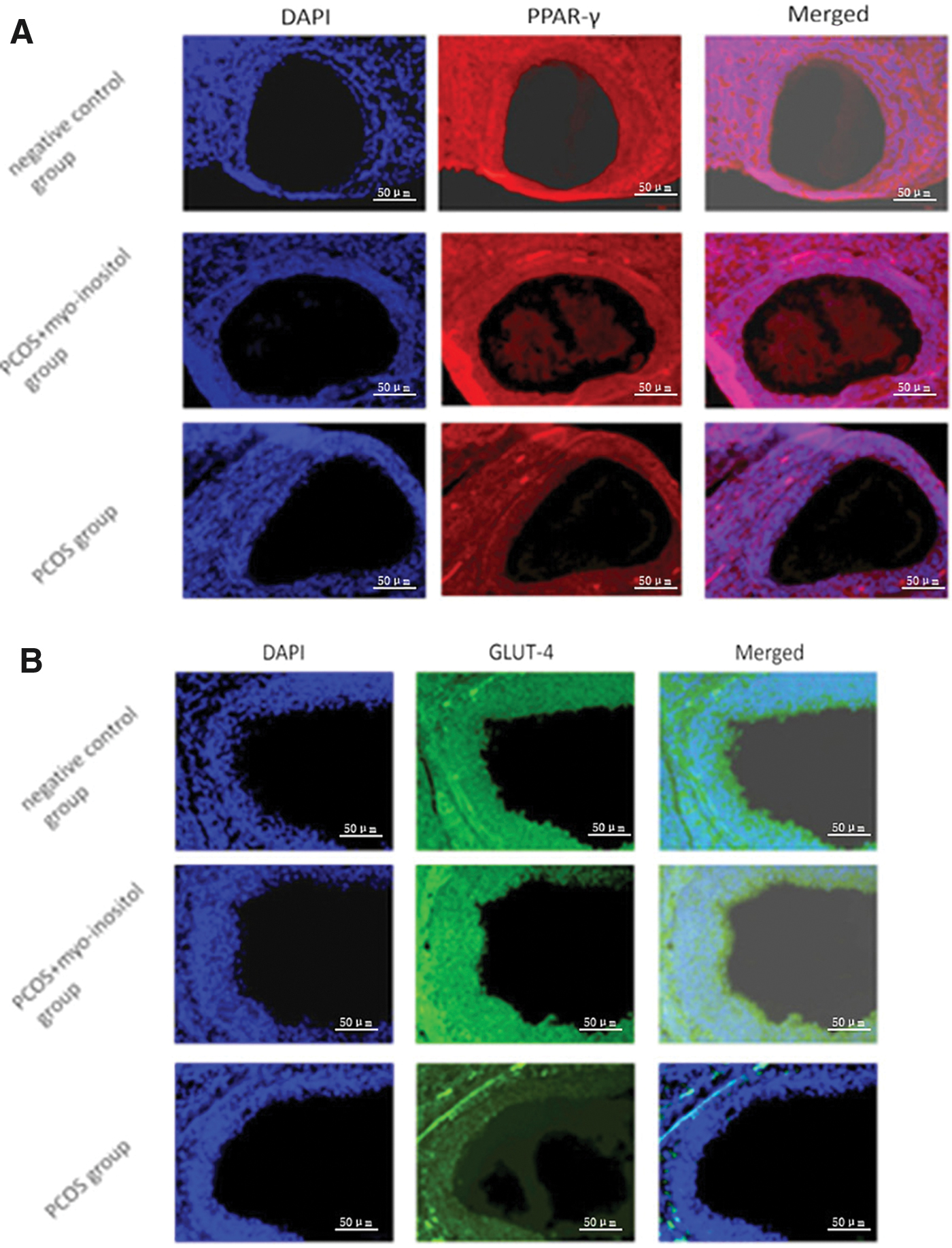
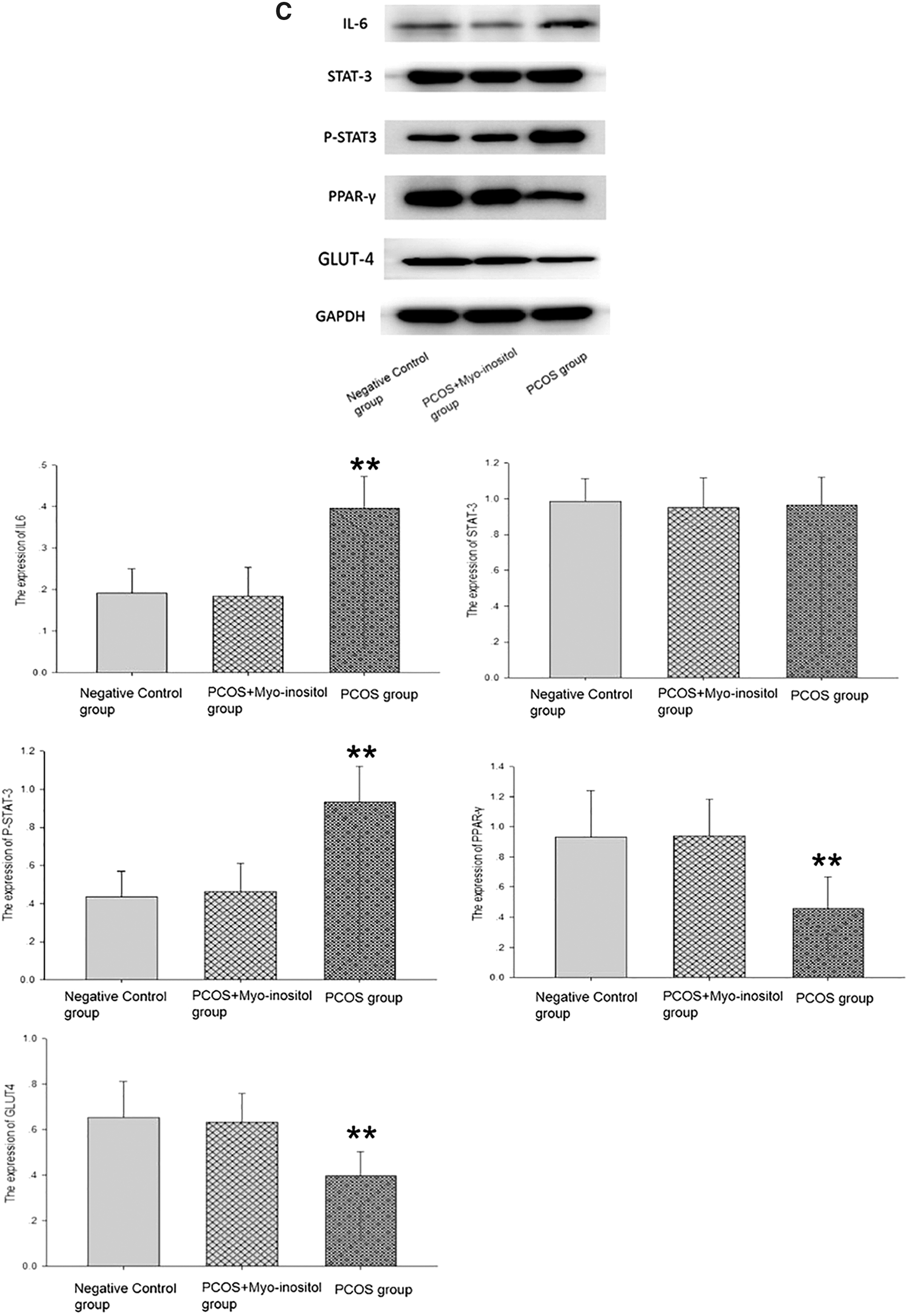
To discover new pathways of myo-inositol action, the expression of PPAR-γ and GLUT4 was detected by immunofluorescence assay in the ovarian tissues of rats from the three groups (Fig. 6A, B). The expression of PPAR-γ and GLUT4 in the PCOS-IR group was significantly elevated after the administration of myo-inositol. Similar results were obtained from western blotting. The expression of IL-6 and p-STAT3 in the PCOS group was significantly higher compared with the PCOS+myo-inositol and negative control groups, and the myo-inositol supplement did not affect the expression of STAT3 (Fig. 6C, **P < .01). However, the expression of PPAR-γ and GLUT4 was significantly lower in the PCOS group than that in the PCOS+myo-inositol and negative control groups (Fig. 6C, **P < .01). All data showed that myo-inositol supplementation reduced the expression of IL-6/p-STAT3, whereas it increased the expression of PPAR-γ and GLUT4 in the ovaries of PCOS-IR rats. Myo-inositol supplementation reversed the upregulation of IL-6/p-STAT3 and increased the expression of PPAR-γ and GLUT4 in those tissues.
Discussion
PCOS is the most common endocrine condition in women of reproductive age, with a prevalence of up to 18–20%. 14 The prevalence of IR in patients with PCOS has been reported to be as high as 77%, whereas the prevalence of type 2 diabetes in these patients was 10%. 15,16 Furthermore, the risk of coronary heart disease and stroke in women with PCOS was increased by twofold, whereas the prevalence of metabolic syndrome by 43%. 17 –19 Currently, drugs and surgery can treat PCOS; however, these approaches present several drawbacks. 20 –22 Surgical treatments, such as laparoscopic ovary perforation and ovary wedge resection, 23,24 have major drawbacks, including destruction of the ovary and premature ovarian failure in some patients. 23,25 Therefore, the surgical treatment of PCOS is rarely used by clinicians. Myo-inositol treatment has the advantages of being natural, less toxic, and effective; therefore, it is increasingly used to treat PCOS. 26 –28
We found that myo-inositol significantly increased the serum levels of E2, decreased the serum levels of LH, T, and triglycerides sharply, and decreased the HOMA-IR index in PCOS-IR rats. In contrast, myo-inositol supplementation led to the recovery of the estrous cycle in PCOS-IR rats, indicating that this molecule can restore ovulation function. Similarly, treatment with myo-inositol reduced the elevated triglyceride content in rats with nonalcoholic fatty liver induced by a high-fructose diet. 29 We found that autophagy was activated in the ovarian tissues, which agrees with the results of another study. 30
However, the mechanism underlying the action of myo-inositol supplementation in reversing IR was unclear. We found a significant reduction in the expression of both IL-6 and p-STAT3 and an increase in PPAR-γ expression in PCOS-IR rats. Our data indicated an elevation of the IL-6/p-STAT3/PPAR-γ/GLUT4 system in PCOS-IR, rats providing an insight into the pathogenesis of PCOS, and a possible therapeutic strategy.
IL-6 has a close relationship with type 2 diabetes and IR. In obese individuals and patients with type 2 diabetes, elevated blood sugar levels, low sugar tolerance, and decreased sensitivity to insulin are related to the upregulation of IL-6. 31,32 Recently, PCOS was significantly related to IL-6 levels, probably because of chronic low-grade inflammation. 6 IL-6 can increase the activity of the androgen receptor indirectly. 33 Elevated IL-6 levels in the high HOMA-IR ratio and T ratio subgroups were found in patients with PCOS, and some studies reported treatment-related reductions in IL-6 levels in these patients. 6 The results of this study showed that IL-6 expression was significantly increased in the ovarian tissues of PCOS-IR rats and that this was related to a disorder of the estrous cycle of these rats and a high HOMA-IR index. In turn, myo-inositol supplementation led to the recovery of the estrous cycle and a decrease in the elevated levels of IL-6 in PCOS-IR rats, indicating that myo-inositol supplementation may be effective for the treatment of PCOS because it downregulates IL-6 expression and promotes the recovery of ovarian function.
STAT3 signaling is activated in women with PCOS, 34 and myo-inositol supplementation reduced the level of p-STAT3. 5 Moreover, the IL-6-mediated JAK2/STAT3 signaling pathway played an important role in PCOS. 9 In our study, we also demonstrated that the level of expression of p-STAT3 was decreased significantly after myo-inositol administration to PCOS-IR rats, probably mostly because of the downregulation of IL-6.
We also showed that PPAR-γ and GLUT4 were sharply downregulated in PCOS-IR rats and that this effect was partially reversed after treatment with the myo-inositol supplement. PPAR-γ, which governs pathways involved in energy metabolism and homeostasis, 35 has an important role in improving insulin sensitivity 36 and has anti-inflammatory properties. 37 Treatment with a PPAR-γ agonist was associated with lower triglyceride and phospholipid levels compared with the control. 38 In this study, we found that the downregulation of PPAR-γ and GLUT-4 was related to the elevation in the level of triglycerides and of the HOMA-IR index. In turn, myo-inositol supplementation reversed the elevation of triglycerides and HOMA-IR, which was accompanied by the upregulation of PPAR-γ and GLUT-4. These results were consistent with those of other studies, as we found that myo-inositol supplementation increased the expression of PPAR-γ mRNA. 39 Another study found that myo-inositol supplementation increased the expression of PPAR-γ and GLUT4. 13
STAT3 signaling stimulates the expression of miR-21 and miR-155. 10,11 Physical interactions between activated STAT3 and its cognate binding sites in the human miR-21 promoter have also been demonstrated, 40 and the Bos taurus miR-21 promoter contains two functional STAT3 binding sites. 41 These results are consistent with our findings: the upregulation of miR-21 and miR-155 was related to the increase in the expression of p-STAT3. PPAR-γ is a target of miR-155, which is a microRNA (miRNA) that is overexpressed in obese individuals. 42 Moreover, it was found that the downregulation of PPAR-γ was associated with upregulation of miR-21. 43 Based on these observations, miR-155 and miR-21 seem to link IL-6/STAT3 signaling to the control of PPAR-γ expression.
To explore the mechanism underlying the action of myo-inositol supplementation in downregulating IL6 and p-STAT3 and upregulating PPAR-γ, the expression levels of miR-21, miR-155, and IL-6 and PPAR-γ mRNAs were also determined by real-time PCR. We found that the IL-6 mRNA was upregulated in PCOS-IR rats and that myo-inositol supplementation reversed this result. Our findings also indicated that the expression of miR-21 and miR-155 was elevated in PCOS-IR rats and that myo-inositol supplementation significantly reversed this upregulation. Finally, the expression of PPAR-γ mRNA was decreased, leading to the downregulation of PPAR-γ.
In conclusion, we observed the downregulation of PPAR-γ and GLUT4 and the significant upregulation of IL-6, p-STAT3, miR-21, and miR-155 in PCOS-IR rats. Myo-inositol supplementation led to the downregulation of IL-6, p-STAT3, miR-21, and miR-155 and to the significant upregulation of PPAR-γ. Myo-inositol supplementation reduced miR-21 and miR-155 levels by downregulating IL-6 and p-STAT3 and that, as the expression of miR-21 and miR-155 was suppressed, the expression of PPAR-γ and GLUT4 increased. The elevation of the IL-6/pSTAT3/miR-155/miR-21/PPAR-γ system in PCOS-IR rats as indicated by our results provides an insight into the pathogenesis of PCOS and hints at a possible therapeutic strategy.
Footnotes
Author Disclosure Statement
The authors declared no conflicts of interest.
Funding Information
This work was supported by Fujian Provincial Health and Youth Research Projects Funding Scheme (2019-1-13).