
Abstract
Platycodon grandiflorum (PG) has been extensively utilized as an herb to relieve phlegm. In this study, the effects of PG root extracts on airway inflammation and cough reflex were investigated, especially using fermented PG extracts (FPE) to increase an active compound, platycodin D by fermentation. FPE significantly reduced the numbers of eosinophils and total cells in the bronchoalveolar lavage fluid (BALF) obtained from lipopolysaccharide/ovalbumin (LPS/OVA)-induced asthma mice versus those of vehicle control. Moreover, in the BALF and the serum, FPE significantly reduced the concentration of IL-17E, a proinflammatory cytokine that causes TH2 immunity, including eosinophil amplification. It was also demonstrated that FPE might relieve inflammations through histological analysis of the lung separated from each mouse. Furthermore, in cough reflex guinea pigs induced by citric acid treatment, FPE treatment significantly reduced the number of coughs versus that of vehicle control, and consequently decreased cough reflex sensitivity. In addition, the total cell number and eosinophils significantly decreased in the BALF obtained from each guinea pig versus that of vehicle control. In in vitro study, pretreatment with FPE in LPS-stimulated RAW264.7 cells significantly reduced the levels of proinflammatory cytokines such as TNF-α, IL-6, and IL-1β, and inducible nitric oxide synthases (iNOS). Therefore, we demonstrated that FPE relieved airway inflammation and cough reflex sensitivity in vivo, and exhibited anti-inflammatory effects through suppression of iNOS and several proinflammatory cytokines. These findings suggest that FPE might have a beneficial effect on respiratory health, and may be useful as a functional food to prevent respiratory diseases.
Introduction
In recent decades, the prevalence of airway diseases, such as chronic obstructive pulmonary disease and asthma, have increased worldwide. They present similar symptoms, such as coughing, expectoration, wheezing, and dyspnea, but different clinical outcomes. In particular, asthma is clinically characterized by high levels of eosinophils and IgE in the blood. 1,2 Bronchial asthma affecting the airways of lung is a chronic inflammatory disease, and distinguished by goblet cell hyperplasia and eosinophilia. 3,4 Allergic asthma can be characterized by gathering eosinophils, and allergen-specific TH2 memory cells at inflammatory regions. 5,6 The activated local TH2 cells cause the production of proinflammatory cytokines and recruit other inflammatory leukocytes. 5,7 According to several previous studies, IL-17E (alternatively called IL-25), induced by mast cells, elicits TH2 cell-mediated inflammatory responses unlike other members of IL-17 family, 8 –11 even though mechanisms to induce allergic immune responses remain unclear.
Platycodon grandiflorum (Jacq.) A. DC., which is indigenous to several East Asian countries, is an herbaceous perennial plant. 12 The root of Platycodon grandiflorum (PG) has been known to have various health benefits, and is widely utilized in food and traditional oriental medicine to relieve cough, excessive phlegm, chest congestion, sore throat, and so on. 13,14 It contains triterpenoid saponins such as platycodins (A, D, D2, and D3), and platycosides. 15 In particular, platycodin D was identified as a potent component having significant biological activities, that is, anti-inflammatory, neuroprotective, antitumor, and antimicrobial activities. 16 –20 It inhibited the gene expression of cyclooxygenase-2 (COX-2) stimulated by phorbol 12-myristate 13-acetate in mouse macrophages, followed by suppression of prostaglandin E2 production. 16,21 It also had an impact on chronic and acute airway disorders through regulating the expression of mucin mRNA and protein levels in an in vitro study, and controlling macrophage infiltration in ovalbumin-stimulated mice, respectively. 22,23 Thus, the amount of platycodin D in the PG extracts is believed to affect its biological activities. Our previous study demonstrated that the amount of platycodin D in the crude extract of PG increased by four times when PG was fermented with Saccharomyces cerevisiae compared with that in nonfermented PG extracts. 24,25
In this study, the suppressive effects of fermented PG extracts (FPE), with an elevated platycodin D content, on airway obstruction and inflammation were explored.
Materials and Methods
Materials
Prednisolone was obtained from Yuhan Co., Ltd. (Seoul, South Korea). Lipopolysaccharides, sodium nitrite, ovalbumin, citric acid, and Griess reagent were purchased from Sigma Aldrich (St. Louis, MO, USA). Fetal bovine serum, Dulbecco's modified eagle medium (DMEM), and penicillin/streptomycin were purchased from GIBCO Invitrogen (Grand Island, NY, USA). Monoclonal antibody against COX-2, polyclonal antibody against inducible nitric oxide synthases (iNOS), and monoclonal antibody against β-actin were obtained from Cell Signaling (Danvers, MA, USA), NOVUS Biologicals (Littleton, CO, USA), and Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA), respectively. Protease inhibitor tablets were obtained from Roche Diagnostics GmbH (Mannheim, Germany). The enzyme-linked immunosorbent assay (ELISA) kits were obtained from R&D Systems (Minneapolis, MN, USA) for IL-6, IL-1β, and TNF-α, and from Thermo Fisher Scientific (Bender MedSystems, Vienna, Austria) for IL-17E and IL-17.
Preparation of FPE
PG roots collected from Geumsan (Chungnam, South Korea) were washed and crushed. It was mixed in water in a ratio of 1 to 4, and sterilized at 121°C for 1 h utilizing KFC-30L fermenter (PharmswellBio Co., Ltd., Seoul, South Korea). Then, the extracts were cooled to 30°C at 100 rpm, 0.3 vvm, and 0.5 bar in the fermenter, and inoculated into culture medium, including S. cerevisiae to be finally 3% (v/v). After incubation for ∼14–16 h, cellulase and amylase were added to that to be finally 0.4% and 0.2%, respectively, followed by incubating it at 50°C for 16 h. After perlite was added to the fermented extracts to be 4% (v/v), it was filtered and then concentrated to 10 Brix utilizing an evaporator (DooYoung Hi-Tech Co., Seoul, South Korea). Cyclodextrin was added to the concentrate (1:1), and sterilized at 95°C for 2 h. It was dried using a spray drying method, and the powder was sieved using a 60-mesh sieve. The content of an active compound, platycodin D isolated from the prepared final product, was analyzed using Alliance HPLC System (Waters, MA, USA). FPE used in this study contained 0.09 mg of platycodin D per 1 g of PG crude extracts.
Experimental animals
Five-year-old male C57BL/6 mice, and 6-year-old male Hartley guinea pigs were obtained from Orient Bio, Inc. (Gapyeong, Gyeonggi, South Korea), and from Samtako, Inc. (Osan, Gyeonggi, South Korea), respectively. The animals were maintained under the controlled conditions of temperature (23°C ± 3°C), ventilation frequency (10–20 times/h), relative humidity (55% ± 15%), and artificial lighting for 12 h (from 8:00 to 20:00). All animals were acclimated before starting the experiment for 7 days, and during the study period, provided freely with water and standard diets. The animal experiments were approved by Institutional Animal Care and Use Committee (IACUC) of the KNOTUS Co., Ltd. (17-KE-168 for mice and 17-KE-097 for guinea pigs).
Experimental design for an asthma mouse model
Male C57BL/6 mice were distributed into five groups (n = 6) postacclimation for 1 week as follows; (1) control group sensitized with a vehicle and challenged with ovalbumin (OVA), (2) negative control group sensitized with lipopolysaccharide (LPS)/OVA and challenged with OVA, (3) positive control group treated with prednisolone once a day during the OVA-challenged period after sensitization to LPS/OVA (day 14–21), (4) experimental groups treated with 200 and 400 mg/kg of FPE, respectively, once a day during the OVA-challenged period after sensitization to LPS/OVA. Through nasal instillation of 10 μg of LPS and 75 μg of OVA, animals were sensitized on 0, 1st, 2nd, and 7th day, respectively, and thereafter challenged through nasal instillation of 50 μg of OVA on the 14th, 15th, and 21th day, respectively.
Bronchoalveolar lavage fluid and blood collection
To evaluate FPE effect on inflammatory responses, bronchoalveolar lavage fluid (BALF) and blood were collected from each mouse on the day after last administration. To obtain BALF, the left bronchus was ligated with 3-0 silk sutures, and a catheter was inserted into the right one. Using the syringe attached to the catheter, phosphate buffer saline (pH 7.4) was injected, and BAL fluid was collected. Blood was collected from the postcaval vein using vacutainer tubes with clot activator. It was incubated for 15 min at room temperature to be allowed to clot, followed by centrifugation for serum separation.
Differential cell counting in BALF
Total differential cells in BALF were immediately counted utilizing a TC10 cell counter (Bio-Rad, Hercules, CA, USA) according to the manufacturer's instructions. The numbers of macrophages, neutrophils, lymphocytes, and eosinophils were determined by flow cytometry as previously described. 26,27 In brief, BALF was incubated with the antibody solution containing CD11b-APC Alexa750, CD11c-FITC, CD3-BV605, CD19-BV650, Siglec F-PE (BD Bioscience, Franklin Lakes, NJ, USA), MHC II-PerCP-Cy5.5, CD45-AlexaFluor700 (BioLegend, San Diego, CA, USA), and Ly-6G-PEeFluor610 (eBioscience, San Diego, CA, USA) for 1 h at 4°C. After monoclonal antibody solution staining, 4′,6-diamidino-2-phenylindole (DAPI) was added to BALF to identify live cells. Cells were sorted on BD FACS Diva software and analyzed on LSR II flow cytometry (BD Bioscience). To remove debris, erythrocytes, and dead cells, live leukocytes were identified by DAPI/CD45 scatter, and separated in a CD11c+ and a CD11c− scatter. Within the CD11c+ scatter, macrophages were gated on Siglec F+/MHC II− plot. Lymphocytes and nonlymphocytes were identified as CD3/19/MHC II plot within CD11c− scatter and B cells and T cells included in lymphocytes were distinguished by MHC II expression. Neutrophils, and eosinophils were gated on CD11b+/Ly-6G+ plot, and on CD11b+/LY-6G−/Siglec F+ plot within the CD11c−/CD3/19−/MHC II scatter, respectively.
Histological analyses
The lungs collected from each mouse after sacrificing were fixed in 10% formaldehyde and were then dissected and embedded using paraffin wax. After sectioning, it was stained with hematoxylin and eosin. They were observed using BX53 light microscope (Olympus, Tokyo, Japan). Histopathologic scoring to determine the level of perivascular and peribronchial inflammation was conducted by masking of the pathologist to experimental groups, preventing bias from entering into the examination and scoring of tissues. A total of lung inflammation was described as the sum of the perivascular and peribronchial scores, and it was scored on four scales: 0, normal; 1, mild; 2, moderate; 3, marked; 4, severe. 28
Determination of IL-17 and IL-17E concentrations in LPS/OVA-stimulated mice by ELISA
The concentrations of IL-17 and IL-17E in the BALF and the serum obtained from LPS/OVA-stimulated C57BL/6 mice administrated to each test article were measured by ELISA in accordance with the manufacturer's instructions (Cat. No. BMS6001 for IL-17, BMS6046-IL-17E; Thermo Fisher Scientific, MA, USA).
Experimental design for citric acid-induced cough reflex animal model
Guinea pigs were distributed into six groups (n = 5) as follows; vehicle-treated control group, 50 mg/kg theobromine-treated group as a positive control, and three different concentrations of FPE-treated groups (50, 200, and 400 mg/kg, respectively). Guinea pigs orally received treatments for seven consecutive days, and then they were exposed to 0.2 M citric acid in glass chambers equipped with a plethysmography system for 10 min postlast treatment. When coughing was induced, cough numbers were measured for 15 min.
FPE treatment in LPS-stimulated RAW264.7 cells
RAW264.7 cells were maintained in DMEM supplemented with 10% fetal bovine serum, streptomycin (100 μg/mL), and penicillin (100 U/mL), and placed in a humidified 5% CO2 incubator at 37°C. Cells were treated with the five different concentrations of FPE (100, 200, 400, 800, and 1600 μg/mL, respectively), and after 2 h, stimulated with 1 μg/mL of LPS, and incubated for 24 h.
Western blot analysis
Cells were lysed in RIPA lysis buffer (50 mM Tris-Cl [pH 8.0], 150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, and 0.1% SDS) with protease inhibitors, and then centrifuged at 10,000 g for 15 min. Total protein concentrations were quantified by bicinchoninic acid assay (Thermo Fisher Scientific). To be separate proteins, sodium dodecyl sulfate polyacrylamide gel electrophoresis was performed and the gel was transferred onto a polyvinylidene difluoride membrane. The blot was blocked with 5% (w/w) skim milk in phosphate-buffered saline with 0.05% tween 20 (PBST), and incubated with first antibodies at 4°C overnight, followed by being incubated with second antibodies for 1 h. Protein bands were detected by an enhanced chemiluminescence detection system (GE Healthcare Life Sciences, PA, USA).
Nitric oxide assay
The level of nitric oxide (NO) was determined through measuring the NO2 − concentration in the supernatant. Each sample supernatant (100 mL) was incubated with Griess reagent for 15 min at room temperature. The absorbance at 540 nm was measured by a microplate spectrophotometer (Epoch2; BioTek Instruments, Inc., Winooski, VT, USA). The concentrations of NO were determined by a calibration curve obtained by a serial dilution of nitrite ion standard solution.
Determination of proinflammatory cytokines concentration by ELISA
TNF-α, IL-6, and IL-1β concentrations in supernatants obtained from LPS-stimulated RAW264.7 cells pretreated with various concentrations of FPE, were measured by ELISA, respectively, in accordance with the manufacturer's instructions (Cat. No. MTA00B for TNF-α, MLB00C for IL-1β, M6000B for IL-6; R&D Systems).
Statistical analysis
Prism 5.03 software was used for statistical analysis (GraphPad Software, Inc., CA, USA). The results for the murine models of asthma were presented as the mean ± SD of six mice. Student's t-test was utilized to analyze statistical differences. The results for the cough reflex animal model were presented as the mean ± SD of five guinea pigs. One-way analysis of variance was utilized to analyze statistical differences, followed by Dunnett's post hoc test. The P value <.05 was regarded as statistically significant.
Results
FPE effects on inflammatory cell recruitments in an asthma-induced mouse model
To examine the effect of FPE on inflammatory cell infiltration into lung, total cells, eosinophils, neutrophils, macrophages, and lymphocytes were counted in the BALF obtained from LPS/OVA-induced asthma mice. By OVA airway challenge after sensitization to LPS/OVA, the numbers of other inflammatory cells except for lymphocytes in the BALF significantly increased versus those of control mice (Fig. 1A, C–E). However, FPE treatments significantly reduced the infiltrations of total cells (FPE 400 mg/kg-treated group only), and eosinophils versus those of the vehicle-treated group (Fig. 1A, E).

Effects of FPE on inflammatory cell infiltration in LPS/OVA-induced allergic asthma mice. Inflammatory cells were counted in the BALF obtained from LPS/OVA-induced mice. Male C57BL/6 mice were divided into five groups (n = 6) as follows: PBS-sensitized and OVA-challenged mice (control); LPS/OVA-sensitized and OVA-challenged mice treated with vehicle (negative control); LPS/OVA-sensitized and OVA-challenged mice treated with prednisolone (10 mg/kg, positive control); LPS/OVA-sensitized and OVA-challenged mice treated with FPE (200, 400 mg/kg). The number of total cells
FPE effects on inflammatory cytokines in BALF and serum
To elucidate FPE effects on TH2-mediated inflammatory responses, ELISA for IL-17 and IL-17E was performed. IL-17 and IL17E cytokines in both serum and BALF from LPS/OVA-sensitized and OVA-challenged mice significantly increased versus those in the phosphate buffered saline sensitized and OVA-challenged mice (control). The treatment with 400 mg/kg of FPE significantly decreased IL-17E concentrations in the serum and the BALF versus the vehicle-treated control in LPS/OVA-stimulated mice (Fig. 2C, D). However, differences were not significant for the concentration of IL-17 between the two groups (Fig. 2A, B).
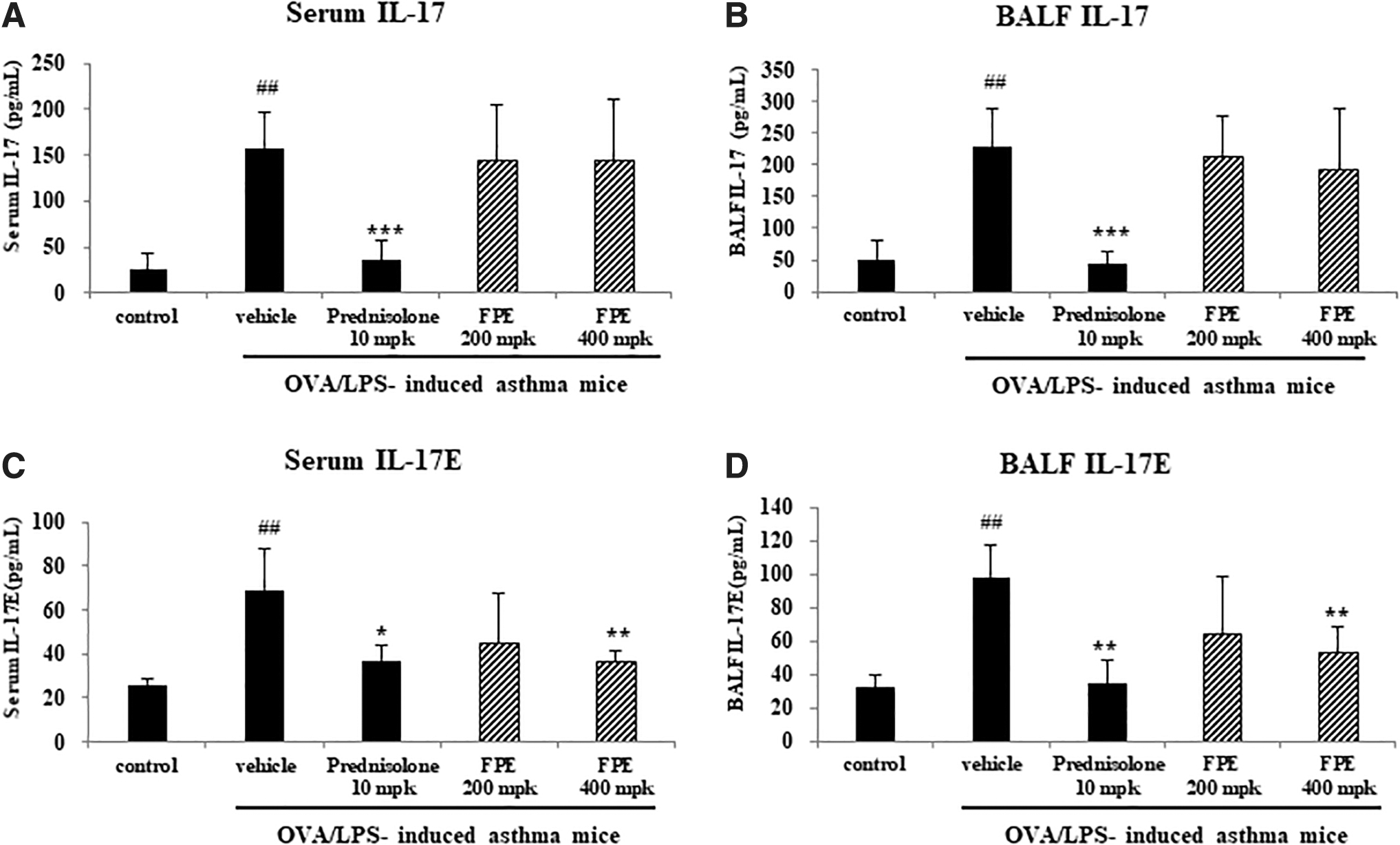
Effects of FPE on IL-17 and IL-17E levels in the serum and the BALF derived from each mouse. BALF and blood were collected from LPS/OVA-induced mice on the day after last administration. IL-17
FPE effects on perivascular and peribronchial inflammations by histological analysis
For further assessment of the effect of FPE on inflammation, histological analysis was performed. As shown in Figure 3A, there were no signs of lung inflammation observed in representative histological sections from the control group. By contrast, peribronchial and perivascular inflammations were induced in LPS/OVA-sensitized and OVA-challenged mice, which indicate the augmented thickness of alveolar septa. In histological examinations, the lung tissues isolated from mice to receive FPE showed alterations in tissue structure. FPE treatment seemed to have an effect on reducing the LPS/OVA-induced inflammation compared with the vehicle treatment. Moreover, the total lung inflammation score combined with bronchiole and blood vessel scores was significantly reduced in 400 mg/kg FPE-treated group versus that of the vehicle-treated group (Fig. 3B).

Effects of FPE on pathological changes in LPS/OVA-induced mice. Lungs were separated from each mouse after sacrificing. The lungs were fixed in 10% buffered formalin solution, and stained with H&E. The slides were visualized through a light microscope (BX53; Olympus, Tokyo, Japan). The representative images of lung were shown by H&E staining
FPE effects on cough reflex sensitivity and inflammatory cell infiltration in the BALF from cough reflex-induced guinea pigs
To assess the effect of FPE on cough reflex sensitivity, the numbers of coughs were counted using an animal model to induce cough reflex by 0.2 M citric acid treatment. The FPE-treated groups (200 and 400 mg/kg, respectively) significantly diminished the number of coughs versus vehicle-treated group, and inhibited cough reflex sensitivity by 25% (Fig. 4). Furthermore, the number of total cells and eosinophils in BALF significantly decreased in FPE-treated group, versus the vehicle-treated group (Fig. 5A, E), whereas in that of lymphocytes, neutrophils, and macrophages, statistical differences were not significant.
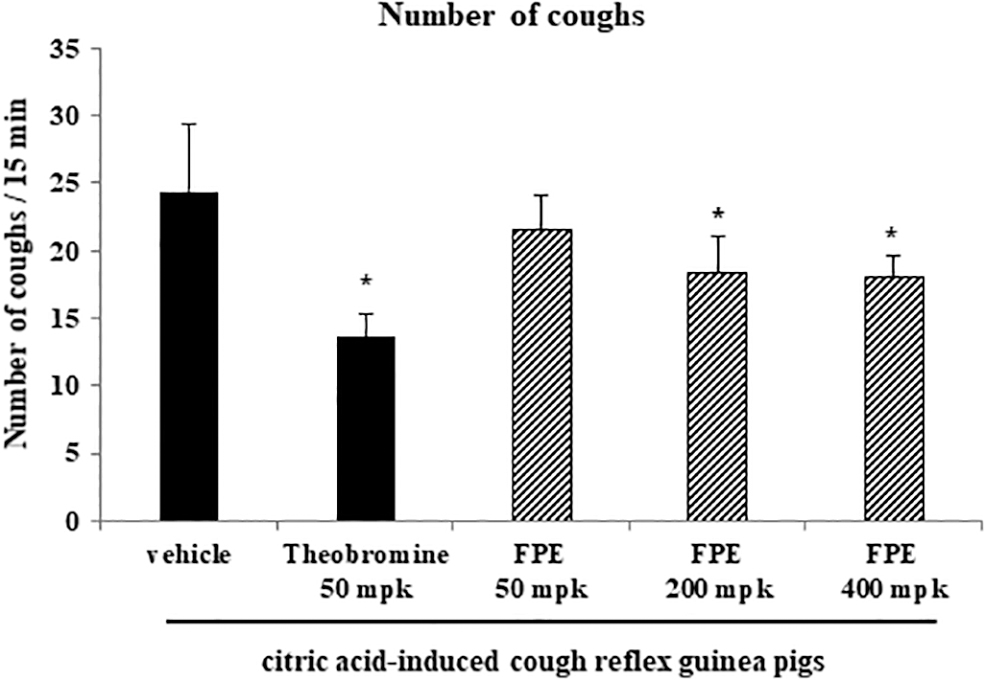
Effect of FPE on cough reflex sensitivity in citric acid-induced cough guinea pig model. The number of coughs was measured using citric acid-induced cough reflex guinea pig model. FPE was orally administered to guinea pigs daily for 1 week, and then all the guinea pigs were exposed to 0.2 M citric acid for 10 min after last administration. Guinea pigs were divided into six groups (n = 5) as follows: vehicle-treated control group, theobromine-treated group (positive control; 50 mg/kg), and FPE-treated groups (50, 200, and 400 mg/kg). Statistical results were presented as the mean ± SD. *P < .05, compared with the vehicle-treated control.

Effects of FPE on inflammatory cell infiltration in the BALF from citric acid-induced cough guinea pigs. Inflammatory cells were counted in the BALF obtained from citric acid-induced cough guinea pigs. Guinea pigs were divided into six groups (n = 5) as follows: vehicle-treated control group, theobromine-treated group (positive control; 50 mg/kg), and FPE-treated groups (50, 200, and 400 mg/kg). The number of total cells
FPE effects on inflammation-mediated protein expression in LPS-stimulated RAW264.7 cells
The main proinflammatory cytokines in charge of early responses are TNF-α, IL-6, IL, and IL-1β, upregulating the synthesis of secondary mediators such as COX-2 and iNOS, and other proinflammatory cytokines induced by macrophages or mesenchymal cells. To explore the molecular mechanism of FPE on anti-inflammatory effects, ELISA for proinflammatory cytokines, and western blot analysis for COX-2 and iNOS were carried out in RAW264.7 cells stimulated by LPS. As shown in Figure 6, FPE treatment inversely reduced the increased production of proinflammatory cytokines by LPS treatment. The concentration of IL-6 was significantly reduced when treated with 400, 800, and 1600 μg/mL of FPE (P < .01), respectively, for IL-1β, when treated with 800 and 1600 μg/mL of FPE (P < .01), respectively, and for TNF-α, when treated with 1600 μg/mL of FPE (P < .05). Furthermore, pretreatment with FPE significantly suppressed LPS-induced iNOS protein expression in dose-dependent manners; however, it did not affect COX-2 protein expression (Fig. 7A). Consistent with the results, NO production increased by LPS treatment was significantly inhibited by FPE treatment (800 and 1600 μg/mL, respectively) (P < .01) (Fig. 7B).

Effects of FPE on proinflammatory cytokines induced by LPS in RAW264.7 cells. RAW264.7 cells were pretreated with FPE (100, 200, 400, 800, and 1600 μg/mL) for 2 h, and then stimulated with 1 μg/mL of LPS for 24 h. Each supernatant was collected and the concentrations of IL-6
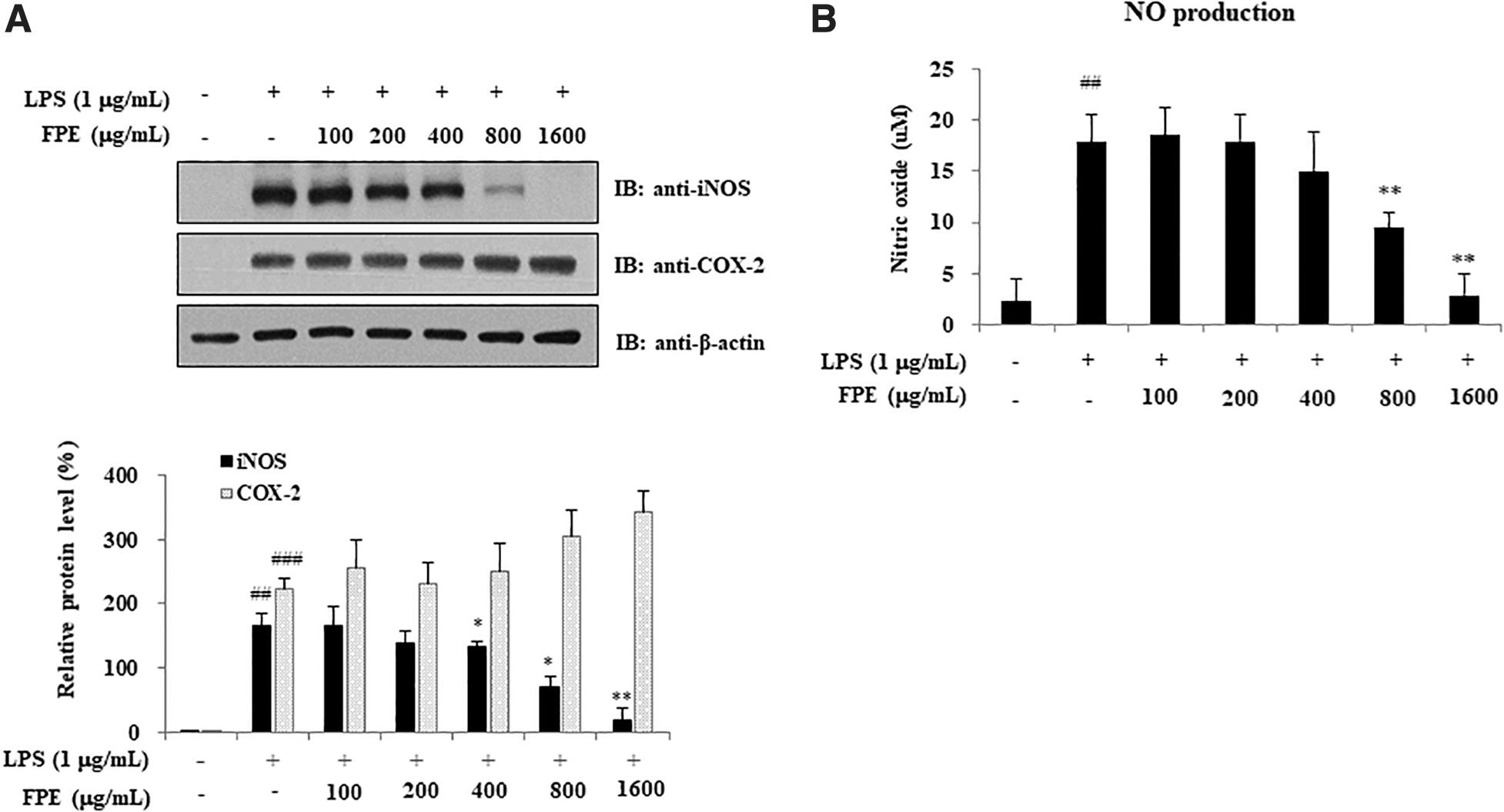
Effects of FPE on iNOS and COX-2 protein expression in LPS-stimulated RAW264.7 cells. RAW264.7 cells were pretreated with FPE (100, 200, 400, 800, and 1600 μg/mL) for 2 h, and then further stimulated with 1 μg/mL of LPS for 24 h. The cells were then harvested, and western blot analysis was performed using anti-iNOS, anti-COX-2, and anti-β-actin antibodies
Discussion
In this study, FPE effects on allergic and respiratory diseases were investigated, and anti-inflammatory functions of FPE involved with eosinophils and macrophages were explored to elucidate molecular process that produce its effects on relieving inflammation.
Allergic asthma is characterized by the elevated eosinophils in the bone marrow, tissue, and blood, and its expansion usually correlates with disease severity. 29 –31 Eosinophil recruitment at inflammatory sites causes it to produce IL-17E that has been reported to enhance allergic responses through a variety of pathways. When overexpressed or acting systemically, IL-17E promotes production of eosinophil chemoattractant, and TH2 cytokines such as IL-4, IL-5, and IL-13, 8,9,32 which increases the infiltration of eosinophils into lungs, increases serum IgE levels, and induces mucus hyperplasia. 33 These previous results suggest that IL-17E might play important roles in developing TH2 immunity and expanding the eosinophils involved in allergic inflammation. IL-17E induces intercellular adhesion molecule-1 expression on eosinophils, 34 and reduces eosinophil cell death. 35 These results show that the increased level of IL-17E receptor and IL-17E in the blood in asthmatics might be related to the continuous elevation in airway eosinophils. In our study, the concentration of IL-17E was markedly elevated in the serum and the BALF derived from LPS/OVA-sensitized and OVA-challenged mice, and infiltrations of inflammatory cells around the airways and pulmonary blood vessels were increased. In contrast, FPE administration significantly reduced IL-17E levels, and further infiltrated eosinophils, which indicates the effect of FPE on reducing airway inflammation.
Activated macrophages stimulated by a variety of stimuli induce various inflammatory mediators such as chemokines, NO, and cytokines. NO is produced by an inducible isoform of iNOS in LPS-stimulated macrophages, which is mediated by a single enzyme process. 36 Thus, to inhibit the expression of iNOS is an essential basis to block NO production in LPS-stimulated cells. 37 In this study, FPE treatment significantly reduced the protein level of iNOS, but not COX-2 in LPS-stimulated cells, suggesting that FPE may differentially regulate the protein levels of COX-2 and iNOS. 38
Therefore, our findings demonstrated that FPE relieved airway inflammation and cough reflex sensitivity in animal models, suggesting that the anti-inflammatory effect of FPE may be based on molecular mechanisms through suppression of iNOS and IL-17E protein expression. These findings indicate that FPE containing increased contents of an effective compound, platycodin D by fermentation, may be more effective as a functional dietary intervention for respiratory health than nonfermented PG.
Footnotes
Acknowledgment
The authors express their gratitude to Korea University.
Author Disclosure Statement
None of the authors have any conflicts of interest to declare.
Funding Information
This research was supported by a grant (code no. P20170015) from Gyeonggi Technology Development Program funded by Gyeonggi Province.