
Abstract
Cisplatin is a widely used anticancer drug that has adverse effects on gastrointestinal function. Curcumin is a natural polyphenol extracted from the rhizome of turmeric that has a wide range of biological activities. The present study investigated the effects of cisplatin on gastric emptying in mice and examined whether these can be inhibited by curcumin. We found that pretreatment with curcumin (200 mg/kg/day) for 10–30 days partly inhibited the decreases in gastric emptying rate and body weight induced by cisplatin. Furthermore, cisplatin reduced acetylcholine (ACh) concentration and the messenger RNA (mRNA) level of ACh receptor (AChR) as well as acetylcholinesterase activity in the stomach of mice; caused ultrastructural damage to interstitial cells of Cajal (ICC); and altered the expression of c-kit/stem cell factor and the gap junction protein connexin 43 in ICC. Curcumin pretreatment inhibited the effects of cisplatin on ACh indicators and ICC. These results demonstrate that curcumin can protect against cisplatin-induced gastric emptying disorder and thus has therapeutic potential for alleviating this condition in cancer patients receiving cisplatin chemotherapy.
Introduction
Cisplatin (cis-diamminedichloridoplatinum[II]; CDDP) is a widely used anticancer drug. However, adverse effects of CDDP are obvious, especially on gastrointestinal system. 1 Some of these include chemotherapy-induced nausea and vomiting (CINV) and gastrointestinal dysmotility, which may involve loss of appetite and indigestion. These effects may aggravate cachexia in cancer patient. 2,3 Certain drugs such as 5-hydroxytryptamine (5-HT) receptor antagonist, neurokinin (NK)-1 antagonist, and dexamethasone are used to prevent nausea and vomiting by the lack of gastrointestinal motility, 4 but do not provide relief for dyspepsia and cachexia.
Curcumin is the active ingredient of turmeric, a spice extracted from the rhizomes of Curcuma longa. In recent years, studies have shown that curcumin has anti-inflammatory, antioxidant, antitumor, and other pharmacologic effects 5,6 and was shown to reduce symptoms of irritable bowel syndrome. 7 Previous studies in our laboratory showed that intragastric administration of curcumin (200 mg/kg) for 15 days improved gastrointestinal food propulsion that was delayed by cisplatin 8 through an unknown mechanism.
We also found that curcumin stimulated gastric food emptying that was delayed by atropine, a specific blocker of acetylcholine (ACh) receptor (AChR), 9 suggesting that curcumin exerts its effects by modulating ACh transmission. In addition to binding directly to its receptor located in smooth muscle cells, ACh also transmits its information to a special type of mesenchymal cell, known as the interstitial cells of Cajal (ICC), which plays a key role in normal gastrointestinal motility. 10,11 According to their relative locations in the gut wall, ICC are sorted into different types: Subserosal ICC (ICC-SS), myenteric ICC (ICC-MY), septal ICC (ICC-SEP), intramuscular ICC (ICC-IM), ICC of the deep muscular plexus (ICC-DMP), and submucosal ICC (ICC-SM). 12
Numerous studies have described neurotransmission between the enteric nerve fibers, ICC, and the smooth muscle cells. 12 –17 ICC are known as gastrointestinal pacemaker cells as they regulate gastric relaxation and emptying, food mixing, and the migrating motor complex of the gastrointestinal tract. 10,12 –15 Integrity of the ICC network plays significant roles on gastrointestinal system motility. 12 Abnormal distribution of ICC has been observed in gastrointestinal disorders, including inflammatory bowel disease, idiopathic slow-transit constipation, and necrotizing enterocolitis, 14,16 and ICC deficiency has been reported in a rat model of diabetic gastroparesis. 17
Curcumin was shown to relieve food retention in the stomach of diabetic rats by affecting the oxidation and expression of c-kit, 18,19 a tyrosine kinase receptor that is specifically expressed by ICC. Stem cell factor (SCF) is c-kit ligand. The SCF/c-kit signaling pathways plays a key role in the proliferation, development, and maintenance of ICC 20,21 ; inadequate SCF/c-kit signaling was found to contribute to intestinal motility dysfunction induced by hypoxia in neonatal mice. 22
We speculated that some of the adverse effects of cisplatin and the beneficial effects of curcumin on gastrointestinal peristalsis are related to signals targeting ICC. To test this hypothesis, in the present study, we investigated whether cisplatin-induced impairment of gastrointestinal motility in mice is associated with changes in ICC as well as in ACh/AChR signaling, including in the ultrastructure, SCF/c-kit signaling, and expression of the gap junction protein (Cx)43. Furthermore, we also examined whether curcumin can ameliorate the adverse effects caused by cisplatin-via modulating of changes in ACh and ICC.
Materials and Methods
Chemicals
Curcumin (99% purity; Sigma-Aldrich, St. Louis, MO, USA) was mixed in a 5% gum Arabic suspension for intragastric administration. Cisplatin (Jiangsu Haosen Pharmaceutical, Liangyungang, China) was diluted with saline to a concentration of 0.25 mg/mL. All other reagents were purchased from Sinopharm Chemical Reagent Co. (Shanghai, China).
Animals and experimental strategy
Male Kunming mice weighing 19–21 g were supplied by the Laboratory Animal Center, Shandong Lukang Pharmaceutical Co. (Qingdao, China; SCXK Lu-20140007). The mice were maintained on a 12:12-h light/dark cycle with free access to food and water. The animal experiments were approved by the Animal Ethics Committee of Qingdao University. After adaptive feeding for 1 week, the mice were randomly divided into normal control (NC), cisplatin (CDDP), curcumin (Cur), and cisplatin+curcumin (CDDP+Cur). The CDDP+Cur group was further divided into 1, 5, 10, 15, 20, and 30 days according to the duration of curcumin treatment. Each group had 10 mice.
Curcumin (200 mg/kg/day) was intragastrically administered for the appropriate number of days, while mice during the days without curcumin treatment or in NC and CDDP groups received solvent solution. Cisplatin (2 mg/kg/day, 0.2 mL) was administered by intraperitoneal injection and started from the last 5th day, for a total of 5 days, while mice in NC and Cur group received saline (0.2 mL). At 24 h after the last intragastric administration, the gastric emptying rate was determined for each group.
Weight and health status of mice
Changes in body weight and general status of mice such as coat appearance, activity level, and food consumption were recorded 5 days before and after cisplatin or saline injection.
Gastric emptying rate
The mice were fasted for 24 h before the gastric emptying test. A semifluid solution was prepared by combing gum Arabic (40%) with blue dextrin 2000 (1%, as a nonabsorbable marker), and 0.8 mL was intragastrically administered to mice, which were sacrificed by cervical dislocation 20 min later. The stomach of each mouse was removed and cut open; the contents were removed and diluted in 2.0 mL of deionized water, then centrifuged (4000 g) for 5 min. The absorbance of the supernatant (Abs) was measured with a spectrophotometer at λ = 260 nm. The standard absorbance (SAbs) was also determined for 0.8 mL semifluid solution. The gastric emptying rate was calculated as Abs/SAbs × 100%.
ACh concentration and acetylcholinesterase activity measurement
Mouse stomach tissue homogenate (10%) was prepared at 4°C in lysis buffer using a commercial kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer's instructions. The supernatant obtained after 10 min of centrifugation at 2250–3250 g at 4°C was combined with the color development reaction solution (200 μL). The samples were transferred to a 96-well plate and the absorbance was measured with an automatic enzyme marker to obtain an optical density value that was used to calculate ACh content and acetylcholinesterase (AChE) activity, which are expressed as a value per gram of tissue.
Quantitative real-time polymerase chain reaction analysis
Total RNA was extracted from mouse gastric tissue using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Total RNA (1 μg) was reverse transcribed using the BcaBEST RNA PCR (polymerase chain reaction) kit (Takara Bio, Otsu, Japan). Quantitative real-time-PCR was performed with SYBR Premix Ex Taq (Takara Bio) on a 7500 Fast Sequence Detection System (Applied Biosystems, Foster City, CA, USA). The cycling conditions were as follows: 95°C for 3 min, followed by 40 cycles of 95°C for 10 sec, 60°C for 20 sec, and 72°C for 30 sec. Experiments were performed in triplicates. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) served as the internal control.
The primer sequences are shown below: For AChE, the forward primer was 5′-ACCTGCTTCTCCCACACCT-3′, and the reverse primer was 5′-GGTTCCCACTCGGTAGTTCA-3′. For AChR, the forward primer was 5′-CATCTTGCTGGCTTTCATCA-3′, and the reverse primer was 5′-CACAGCCAGTAGCCCAGATT-3′.
For c-kit, the forward primer was 5′-AGCGTGTGTAAATCGTGTTTG-3′, and the reverse primer was 5′-ACATTCAGCATTCCTCCCATA-3′. For SCF, the forward primer was 5′-ATAGACTCCAGCGTCTTCCG-3′, and the reverse primer was 5′-GGCCTGGATTTGCTCTTTGTT-3′. For Cx43, the forward primer was 5′-ACCCAACAGCAGCAGACTTTGA-3′, and the reverse primer was 5′-GCTTGGACCTTGTCCAGCAG-3′. For GAPDH, the forward primer was 5′-CGGAGTCAACGGATTTGGTCGTAT-3′, and the reverse primer was 5′-AGCCTTCTCCATGGTGGTGAAGAC-3′. Relative messenger RNA (mRNA) levels were calculated with the 2−ΔΔCT method and are presented as the n-fold difference of each sample.
Western blotting
Protein was extracted from stomach tissue by homogenization at 4°C in radio immunoprecipitation assay (RIPA) lysis buffer supplemented with phenylmethylsulfonyl fluoride (PMSF). The lysate was cleared by centrifugation at 10,000 rpm/min and 4°C for 5 min. The protein concentration in the supernatant was quantified and 10 μg of protein was loaded into each lane of a 5% concentrated gel and separated on an 8% gel by electrophoresis.
Proteins were electrotransferred to a polyvinylidene fluoride (PVDF) membrane that was probed with rabbit anti-SCF/c-kit and Cx43 antibodies (Bioss, Woburn, MA, USA) used at 1:500 dilution, followed by incubation with horseradish-peroxidase-conjugated goat anti-rabbit IgGs secondary antibody. The electrochemiluminescence (ECL) luminescent dye was dripped on the PVDF and developed a picture of protein bands in a developing camera obscura. The signal intensity was analyzed with gel imaging system.
Ultrastructural changes in ICC
Mice were sacrificed by cervical dislocation, and gastric tissue (0.3 × 0.3 mm) was removed and placed in a fixative solution composed of 3% glutaraldehyde and 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.2) for 2 h at 4°C. After rinsing the sample in the same buffer, specimens were postfixed in 1% osmium tetroxide for 2 h at 4°C, rinsed in distilled water, block-stained with saturated uranyl acetate solution for 3 h, dehydrated in a graded series of ethyl alcohol, and embedded in epoxy resin. Ultrathin sections were cut using a Reichert microtome (LabX, Midland, Canada) and double-stained with uranyl acetate and lead tartrate. The sections were observed with an electron microscope (JEM 1200EX II; Jeol, Tokyo, Japan). Interstitial cells connected to peripheral nerve fibers and smooth muscle cells were examined and imaged.
Immunofluorescence analysis
Mice were intraperitoneally anesthetized with thiotetrabarbital (100 mg/kg; Sigma-Aldrich) and the chest was cut open to expose the heart. The right atrium was incised and the saline (0.9%, 50 mL) was perfused rapidly from the left ventricle to flush the blood, then 4% paraformaldehyde (in 0.1 M sodium phosphate buffer, pH 7.4) was replaced to perfuse until the limbs of the mouse are stiff. The stomach was removed and a 2 × 2 cm piece of the corpus was dissected and postfixed in 4% paraformaldehyde for 4–6 h, then transferred to 30% sucrose solution for 24 h at 4°C for complete dehydration. The samples were embedded in optimal cutting temperature compound and frozen at −20°C. Serial coronal sections were cut at a thickness of 8 μm on a freezing microtome.
For immunofluorescence labeling, sections were incubated for 24 h at 4°C with rabbit anti-c-kit antibody (1:250; Bioss), washed, and incubated for 1 h at room temperature with fluorescein isothiocyanate-conjugated goat anti-rabbit IgG (1:500; Jackson ImmunoResearch, West Grove, PA, USA). Fluororescence was visualized and images were acquired with a fluorescence microscope (BX63F; Olympus, Tokyo, Japan) equipped with a digital camera (DP80; Olympus). Protein expression levels were quantified by determining the ratio of mean fluorescence intensity to total fluorescence from the images using Jieda image analysis software (Jieda, Beijing, China). The ratio of mean staining intensity and the proportion of positive staining were automatically detected and added together as relative index.
Statistical analysis
Data are expressed as mean ± standard deviation unless otherwise indicated. Differences between groups were evaluated by one-way analysis of variance, and were considered significant at P < .05.
Results
Changes in health status and body weight
Mice treated with cisplatin were thin, less active, and less aggressive than those in the NC group. The weight loss induced by cisplatin was inhibited by preadministration of curcumin for 10–30 days: Body weight was higher in the CDDP+Cur groups than in the CDDP group (P < .01), although no differences were observed after 1 or 5 days of treatment (Fig. 1A).

Changes in body weight and gastric emptying rate.
Changes in gastric emptying rate
Gastric emptying rate was reduced by cisplatin (P < .01). Pretreatment with curcumin for 10, 15, 20, or 30 days, inhibited the changes of the gastric emptying rate of mice reduced by CDDP (P < .05), although administration of curcumin for 1 or 5 days had no significant effect (P > .05). In a follow-up experiment, treatment with 200 mg/kg/day curcumin for 15 days did not alter gastric emptying rate in normal mice (P > .05) (Fig. 1B).
Changes in ACh indicators in mouse stomachs
After cisplatin injection, ACh concentration and AChE activity in the stomach were decreased (P < .05) (Fig. 2A, B) along with AChE and AChR mRNA levels (P < .05 and <.01, respectively) (Fig. 2C). Pretreatment with curcumin partly reversed these changes above (P < .05). In normal mice, neither cisplatin nor curcumin had any effect on these variables (P > .05) (Fig. 2).
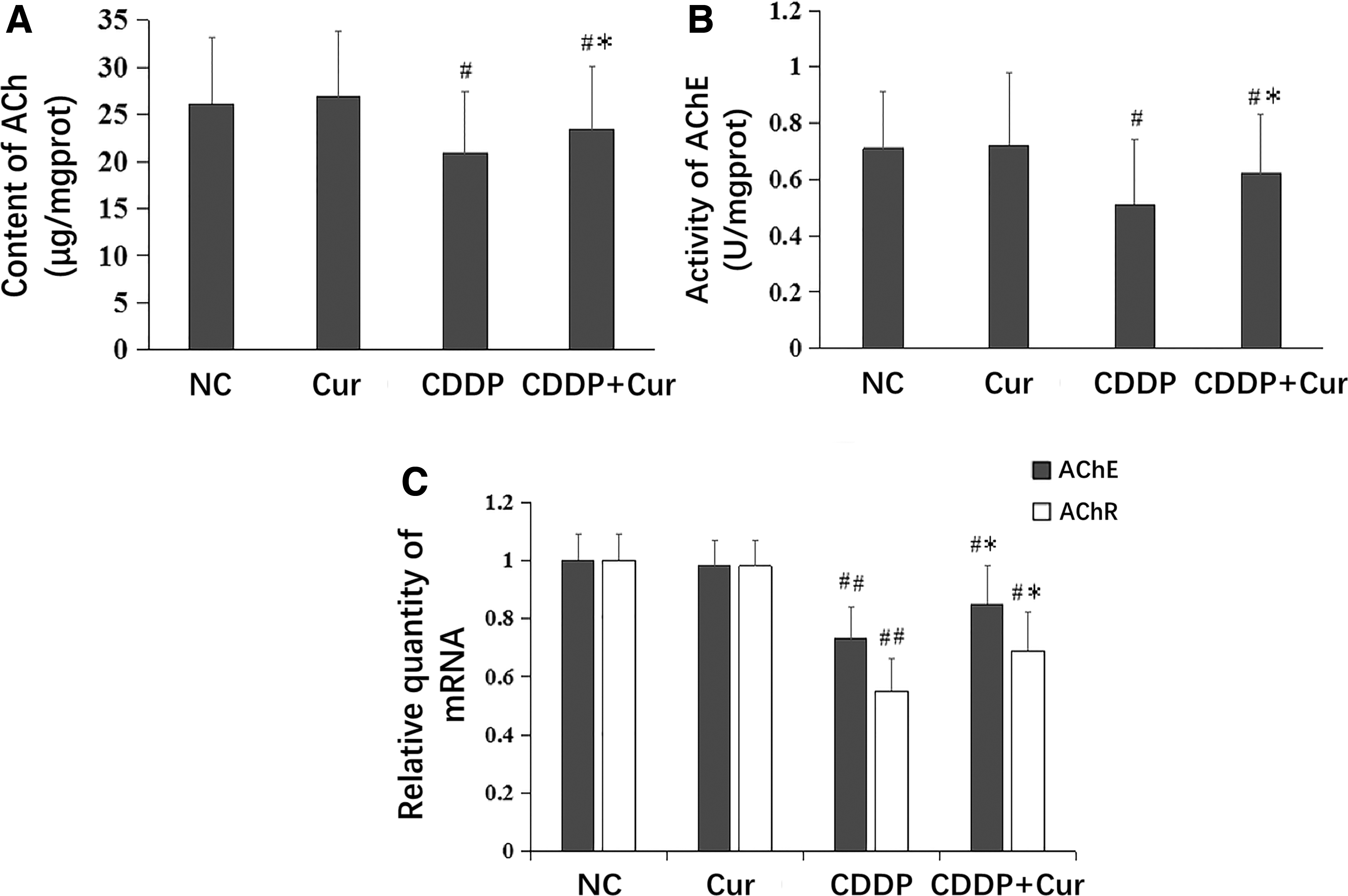
ACh, AChE, and AChR changes in mice stomachs. ACh concentration
Changes in ICC indicators in mouse stomach
Ultrastructure damage to ICC
Electron microscopy ( × 20,000) examination of ICC in mouse stomach revealed an electron-dense cytoplasm, caveolae, and gap junctions formed with smooth muscle cells, as well as close contact between ICC and nerve terminals. Cisplatin caused some ultrastructural damage to ICC, as evidenced by the shrunken and irregular morphology and enlargement of the space between the gap junctions connecting ICC and smooth muscle cells, or between ICC and nerve terminal. In addition, nuclear atrophy increased heterochromatin, and nuclear envelope breakdown were observed along with decreased number of mitochondria and cytoplasmic volume and an increased number of lysosomes. These changes were partly alleviated in the CDDP+Cur group (Fig. 3).
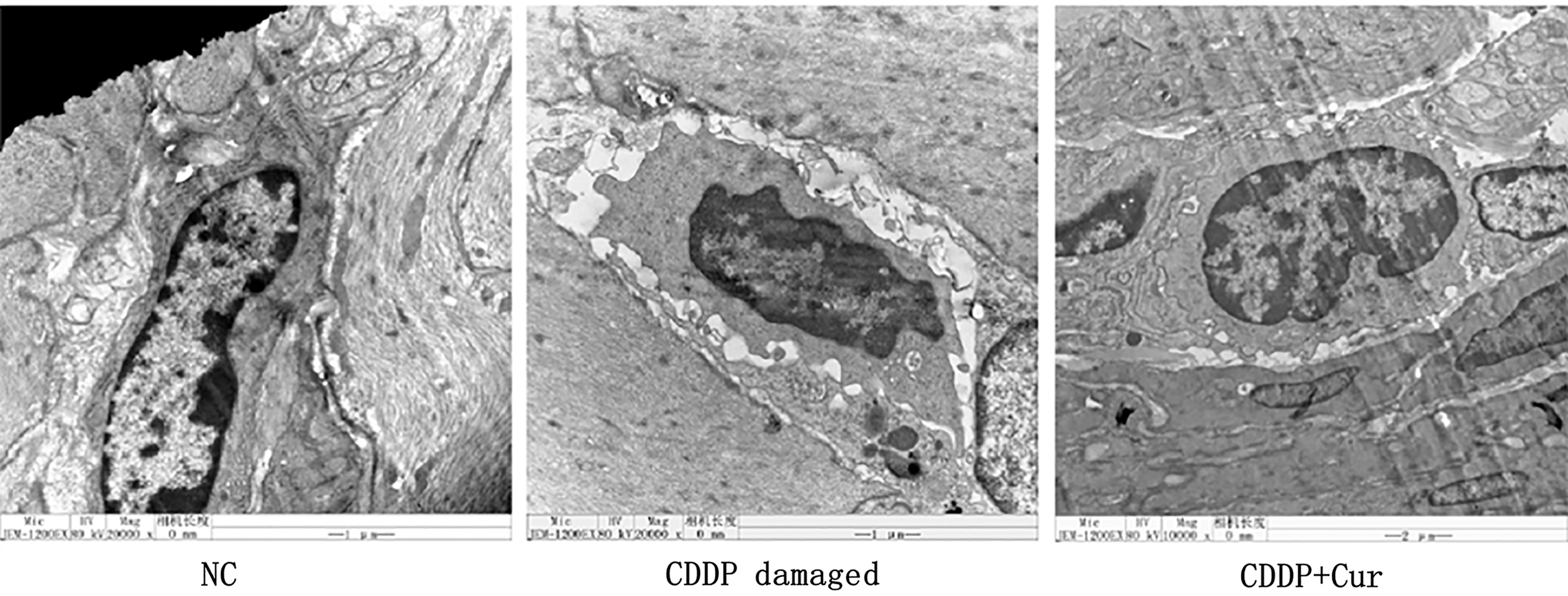
Ultrastructural changes in ICC. ICC were observed by electron microscopy (20,000 × ). Cells were characterized by an electron-dense cytoplasm, caveolae, gap junctions with a smooth muscle cell, and close contact with a nerve terminal. ICC were obviously damaged by CDDP, but this was partly alleviated in the CDDP+Cur group. No obvious changes in stomach smooth muscle cells surrounding ICC were observed in any of the mice. Cur, curcumin; ICC, interstitial cells of Cajal.
Changes in SCF/c-kit expression
The protein levels of SCF and c-kit were evaluated by western blotting. The results showed that c-kit expression was downregulated in gastric tissue of mice in the CDDP group (P < .05). Pretreatment with curcumin inhibited this decrease partly (P < .05). Similar trends were observed for SCF protein, and mRNA levels of SCF and c-kit were consistent with expression of the proteins in the four groups (Fig. 4A). Curcumin had no effect on the expression of SCF and c-kit in normal mice (P > .05) (Fig. 4B). Immunofluorescence labeling revealed a decrease in c-kit immunoreactivity between the longitudinal and circular muscles in the CDDP group (P < .05) that was ameliorated in the CDDP+Cur group (P < .05). These results were confirmed by western blotting (Fig. 4C, D).

Changes in SCF/c-kit expression.
Changes in the Cx43 expression
The relative quantity of Cx43 protein in gastric tissue of mice was decreased in the CDDP group (P < .01). Cx43 protein was partly restored in the CDDP+Cur group, although the level did not differ significantly from that in the CDDP group (P > .05). Curcumin had no effect on Cx43 expression in normal mice (P > .05) (Fig. 5). Similar trends were observed in the levels of Cx43 mRNA in all four groups (Fig. 5).

Changes of Cx43 expression. Protein and mRNA levels were determined by western blotting and qRT-PCR, respectively, relative to the reference gene GAPDH. Data are expressed as mean ± SD. # P < .05, ## P < .01 versus NC (one-way analysis of variance). GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Discussion
The results of this study demonstrate that preadministration of curcumin (200 mg/kg/day) for 10–30 days had a sustained ameliorative effect on gastric emptying rate in mice treated with cisplatin, and also improved overall health and body weight. This is consistent with our previous finding that curcumin stimulated gastrointestinal peristalsis that had been reduced by atropine in mice. 9
To determine whether curcumin directly induces the gastrointestinal peristalsis, we examined the effect of a single intragastric administration of curcumin, but found that this did not affect the delay in gastric emptying rate caused by cisplatin. Moreover, curcumin treatment for up to 15 days had no obvious effect on normal mice. This suggests that curcumin does not directly stimulate gastric emptying but instead functions by inhibiting the action of cisplatin; this is likely achieved through its metabolites, given that the effects were only observed after prolonged (>10 days) administration.
ACh indicators in the stomach of mice, including ACh concentration, AChR mRNA expression, and AChE activity, were reduced by cisplatin; pretreatment with curcumin decreased these changes, suggesting that it can improve gastric emptying disorder caused by cisplatin by modulating ACh transmission in mouse stomach. This is supported by previous reports that curcumin improved ACh neurotransmission and thereby alleviated brain dysfunction in rodent models, 23 –25 although the detailed mechanism remains unclear. In addition, further research is needed to determine whether curcumin regulates gastrointestinal motility through pathways other than ACh signaling.
As previously mentioned, ICC are necessary for normal gastrointestinal motility and are involved in ACh transmission in the gastrointestinal tract. Abnormalities in both the number and function of ICC in the stomach are associated with the gastric motility disorders. 26 Our study found that the ultrastructure of ICC was damaged by cisplatin, especially their connections to the smooth muscle cells or nerves. This was accompanied by a downregulation of SCF/c-kit in ICC. Pretreatment with curcumin restored ICC ultrastructure and SCF/c-kit levels. The expression of Cx43, a gap junction protein connecting ICC and smooth muscle cells, was also downregulated by cisplatin and increased following treatment with curcumin. Thus, the gastrointestinal side effects caused by cisplatin are partly related to ICC damage that is partly reversible by curcumin.
Clinically, CINV is a highly distressing syndrome for cancer patients undergoing cisplatin therapy; although the underlying mechanisms are not well understood, 27 impaired gastrointestinal peristalsis may be one contributing factor. Our results showed that curcumin improved the decreased rate of gastric emptying caused by cisplatin, although it is unknown whether it has an effect on nausea or vomiting. However, given that curcumin indirectly increases gastric emptying, it is reasonable to speculate that it can also alleviate nausea and vomiting caused by chemotherapy drugs by reducing food accumulation and enhancing motility in the stomach.
In the past decades, evidence from in vitro or in vivo studies has provided a basis for evaluating the therapeutic potential of curcumin in clinical trials. 28 –30 High doses of curcumin taken orally were shown to be well tolerated, 31 but the low bioavailability of curcumin when administered through this route limits its clinical applicability.
Recent preclinical studies reported very low concentrations of curcumin in the blood of female rats following oral administration, but all 3 oral curcumin formulations that were tested induced antioxidant genes. 31 The fact that curcumin metabolites are detected in plasma following oral administration 32 and that its degradation products are more potent free radical scavengers than the parent compound 32 strengthen our view that curcumin counters the effects of cisplatin through its metabolites. Recent studies have attempted to overcome the problem of low bioavailability of curcumin using self-assembled mixed micelles, 33 nanoparticles, 34 and solid dispersions 35 ; most of these increased not only the absorption but also the therapeutic efficacy of curcumin, which provide a very attractive prospect in the development and utilization of curcumin.
In conclusion, pretreatment with curcumin can alleviate the cisplatin-induced decrease in gastric emptying rate via modulation of ACh transmission and restoration of ICC structure and function. These findings provide early evidence supporting the physiologic benefits and clinical potential of curcumin in improving gastrointestinal functioning in cancer patients receiving cisplatin as chemotherapy.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Natural Science Foundation of Shandong Province, P.R. China (No. ZR2014HM033).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.