
Abstract
Ethanol extract (RET) of Rosa multiflora Thunb flowers and its subfractions in ethylacetate (REA) or n-butanol subfractions (RBT) were reported to have potent antioxidative and anti-inflammatory activities. In this study, we investigated if those Rosa multiflora flower (RMF) extracts prevent ultraviolet (UV)-induced biochemical damages leading to photoaging. In keratinocyte or dermal fibroblasts, RET, REA, and RBT treatments with UV irradiation significantly decreased reactive oxygen species (ROS), interleukin (IL)-6, IL-8, and matrix metalloproteinase (MMP)-1 levels through suppression of nuclear factor kappa B and mitogen-activated protein kinases. In the animal experiment, mice were orally supplemented with RET (RET group) or REA and RBT mixture (RM group) for 10 weeks, concomitantly with UV exposure. Tumor necrosis factor alpha production and MMP-13 expression were reduced in the mouse skin of RET and RM groups compared with those in the UV control (UVC) group. UV-induced IL-6 production and epidermal thickening were reduced in RM group compared with those in UVC group. Eight phenolic compounds, including quercitrin (quercetin-3-O-rhamnoside), were identified in RMF extracts. Quercitrin treatment to dermal fibroblasts significantly attenuated an increase of MMP-1 expression and a decrease of type I procollagen expression caused by UV. Collectively, RMF extracts showed protective effects from UV-induced photoaging in the skin through suppression of ROS generation, proinflammatory cytokine production, and MMP expression. Quercitrin is suggested to be one of the effective compounds.
Introduction
It is known that ultraviolet (UV) radiation from sun light is a major environmental factor leading to skin aging. UV exposure induces the oxidative stress and proinflammatory response in skin, which are the main cause factors leading to burns, aging, and carcinogenesis. 1,2 UV-induced skin aging is characterized by wrinkles, thickness, roughness, dyspigmentation, and histological changes. 3 –5
Many researchers have investigated natural compounds in an attempt to develop an anti-UV product from plants and foods. 6 Since natural compounds have various biological activities such as antioxidative, anti-inflammatory, or immunosuppressive, many of them have been expected to have a photoprotective effect. It was reported that green tea polyphenols, berry anthocyanin, and grape resveratrol protected the skin against photoaging. 7
Reactive oxygen species (ROS) play a critical role in the skin aging process through induction of matrix metalloproteinases (MMPs) expression. 8 MMPs are endoproteinases that degrade macromolecules in extracellular matrix (ECM), such as collagen, elastin, proteoglycans, and fibronectin. 3,9 MMP-1, one of the interstitial collagenases, initiates the degradation of type I and III collagen. Because collagen is a major structural component of dermis, destruction of collagen is known to be mainly responsible for wrinkle formation. 10
Rosa multiflora Thunb is a species of rose. Recently, its draft genome sequence was determined by Nakamura et al. 11 The fruits and roots of R. multiflora Thunb were known to be used in China as a folk medicine for edema, osteoarthritis, and rheumatoid arthritis, 12,13 whereas the flowers have been consumed as an herbal tea in Korea.
There are a few reports that extracts of this plant showed antioxidant and anti-inflammatory activities, 12,14,15 ; however, there is no report on their biological functions in skin. Recently, we observed that some subfractions of the ethanol extract from R. multiflora flowers (RMFs) contained very high concentrations of polyphenols and flavonoids, and had a potent antioxidant and anti-inflammatory activities. 16 In this study, thus, we investigated if ethanol extract of RMF and two different subfractions protect against skin photoaging caused by UV using human skin cells and mice.
Materials and Methods
Sample extraction and phenolic compounds analysis
We purchased the dried RMF from the herb tea company (NANA and NAMU, Jeongseon, Korea) and freeze-dried it for 24 h followed by grinding in our laboratory. The voucher specimen (2007-0212) has been deposited at Korea University Herbarium. As shown in Figure 1, the dried sample was extracted in ethanol and further extracted with several solvents sequentially. Finally, we lyophilized the ethanol extract of RMF (RET) and ethylacetate (REA) and n-butanol subfractions (RBT). The yield of RET, REA, and RBT from dried RMF was 18.5%, 2.3%, and 3.2%, respectively. Major phenolic compounds existing in RET, REA, and RBT were analyzed using an Ultra Performance Liquid Chromatography system (Waters Co., Milford, MA, USA) as previously reported. 17
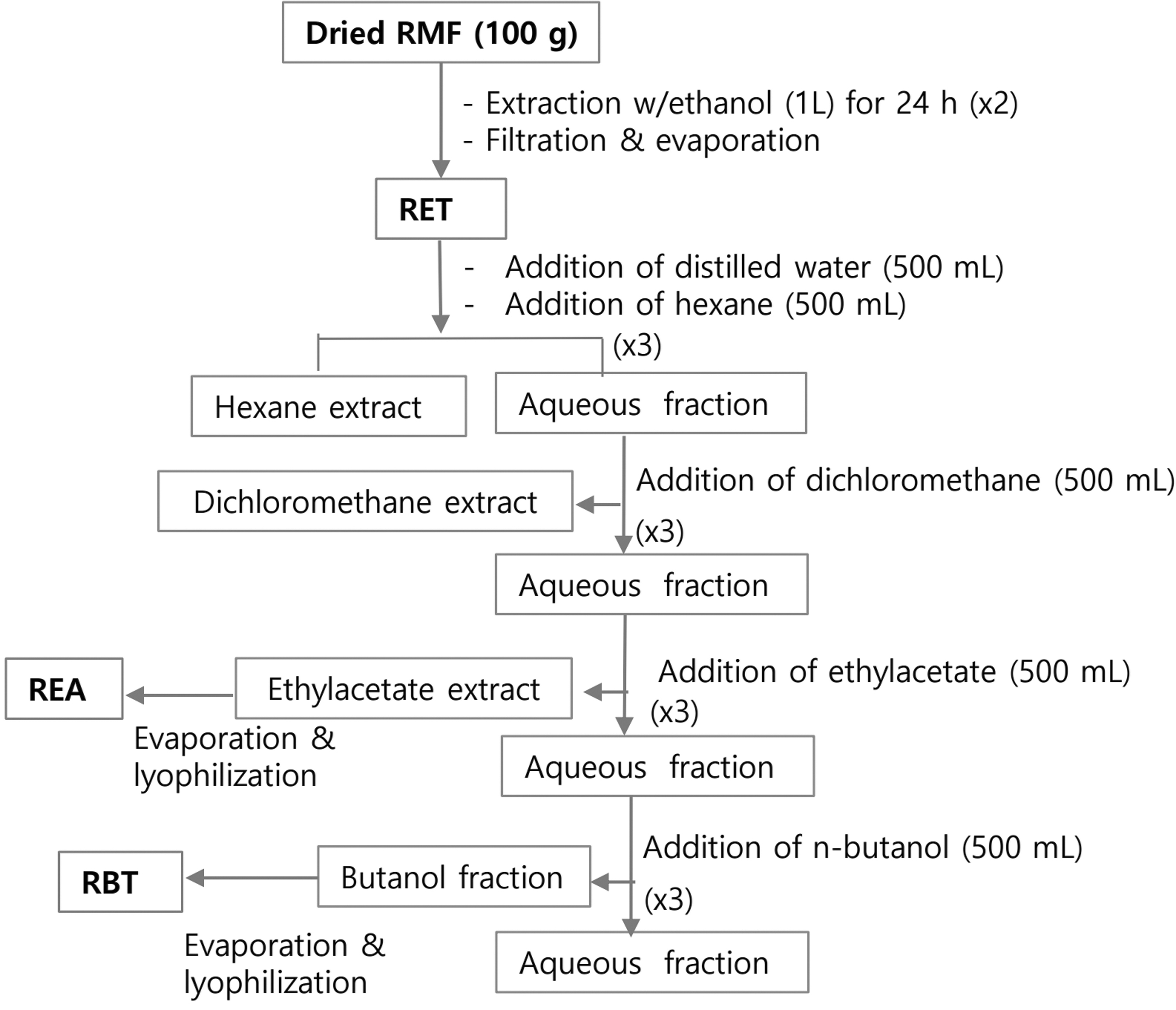
Procedure of sample extraction and fractionation from RMF. RMF, Rosa multiflora flower.
Cell culture, UV exposure, and sample treatment
Human keratinocyte (HaCaT) cells or human dermal fibroblasts (HDFs) were cultured in Dulbecco's modified Eagle's medium (Welgene, Gyeongsan, Korea) supplemented with 10% fetal bovine serum, 2 mM glutamine, 100 U/mL penicillin, and 50 μg/mL streptomycin (Welgene) in 5% CO2 at 37°C. Cells were irradiated with UV (275–380 nm, peak at 310–315 nm) using a fluorescent sun lamp (TL20W/12RS; Philips, Einthoven, Netherlands) with Kodacel filter (Kodak, Rochester, USA) blocking UVC. Sample extract was dissolved in dimethyl sulfoxide (DMSO), filtered, and treated. Cell viability was measured according to the Mosmann's methods. 18
Measurement of intracellular ROS production
HaCaT cells were treated with 2′-7′-dichlorofluorescein diacetate solution (40 μM) for 30 min at 37°C, exposed to UV, and then incubated for another 30 min. Fluorescence of supernatant was measured using a spectrophotometer microplate reader (TECAN, Zurich, Switzerland) at 485 nm of excitation and 530 nm of emission.
Animal treatments and epidermal thickness measurement
Female SKH-1 hairless mice (7-week-old) were purchased from Orient Bio Inc. (Seoul, Korea) and kept in a pathogen-free facility under the standard condition with regulated temperature and humidity, and 12-h light/dark cycle. Mice were allocated into four groups (n = 6/group) after an acclimation period of 1 week; (1) normal control (NC) group: sham-irradiated and vehicle (2.5% DMSO) treated, (2) UV control (UVC) group: UV irradiated and vehicle treated, (3) RET group: UV irradiated and RET treated, and (4) RM group: UV irradiated and mixture of REA and RBT (1:1.5) treated. Sample extracts were orally administered to the mice at 100 mg/kg body weight for 5 days/week for 10 weeks, whereas the dorsal skin was irradiated 3 times/week. UV strength was increased with time from 1 minimal erythema dose (MED, 100 mJ/cm2) to 2.5 MED as previously reported. 18
The animal experimental protocol was approved by the Institutional Animal Care and Use Committees at Seoul National University Hospital (IACUC No. 16-0037-S1A0). After sacrifice, mouse skin samples were hematoxylin and eosin (H&E) stained, and then epidermal thickness were measured as previously described. 17
Measurement of proinflammatory cytokine production
Concentrations of interleukin (IL)-β, IL-6, IL-8, and tumor necrosis factor alpha (TNF-α) in the conditioned cell medium or mouse skin sample were measured using commercial ELISA kits (R&D Systems, Minneapolis, USA).
Quantitative real-time polymerase chain reaction
Total RNA was isolated from the cells or mouse skin samples using Trizol reagent (Ambion-Thermo Fisher, MA, USA). cDNA preparation and quantitative real-time polymerase chain reaction were performed as previously reported. 17 The sequences of primers and a reference gene (36B4) are shown in Table 1. The relative level of mRNA expression was calculated using the 2−ΔΔCT method. 19
Primers for Polymerase Chain Reaction Experiment
F, forward; MMP, matrix metalloproteinase; R, reverse.
Western blotting
Protein was isolated from the cells or mouse skin samples as previously reported. 17 Equal amounts of protein were separated on 8% or 10% sodium dodecyl sulfate-polyacrylamide gels and transferred to PVDF membranes (GE Healthcare, Germany). After the incubation in a blocking solution for 1 h, the membranes were incubated overnight at 4°C with primary antibody against MMP-1 (Lab Frontier, Seoul, Korea), MMP-13 (Thermo Fisher Scientific), human type I procollagen (SP1. D8; Developmental Studies Hybridoma Bank, Iowa City, IA, USA), mouse Collagen-1 (Calbiochem; EMD Millipore Co., Temecula, CA, USA), p65, p-p65, JNK, p-JNK, ERK, p-ERK, p38, p-p38, α-tubulin or β-actin (Santa Cruz), and then incubated with a secondary antibody. The protein bands were visualized using the enhanced chemiluminescence solution (Tanslab, Daejeon, Korea).
Statistical analysis
Results are given as mean ± standard deviation. The differences between groups were determined by Wilcoxon rank-sum test using SAS version 9.4. It was considered statistically significant when P < .05.
Results
ROS and proinflammatory cytokine levels in HaCaT cells
Treating HaCaT cells with RET, REA, and RBT for 24 h did not affect the cell viability substantially (Fig. 2A). UV irradiation (50 mJ/cm2) significantly increased intercellular ROS levels to 193% of normal control (P < .01). However, 4 h pretreatment of RET, REA, and RBT significantly inhibited UV-induced ROS generation by 12.6% (P < .05), 15.5% (P < .01), and 8.5% (P < .05) at 50 μg/mL, respectively, and inhibited it by 27.0%, 22.6% (P < .01), and 11.8% (P < .05) at 100 μg/mL, respectively. Ascorbic acid treatment (100 μM), a positive control, inhibited it by 25.4% (P < .01) (Fig. 2B).
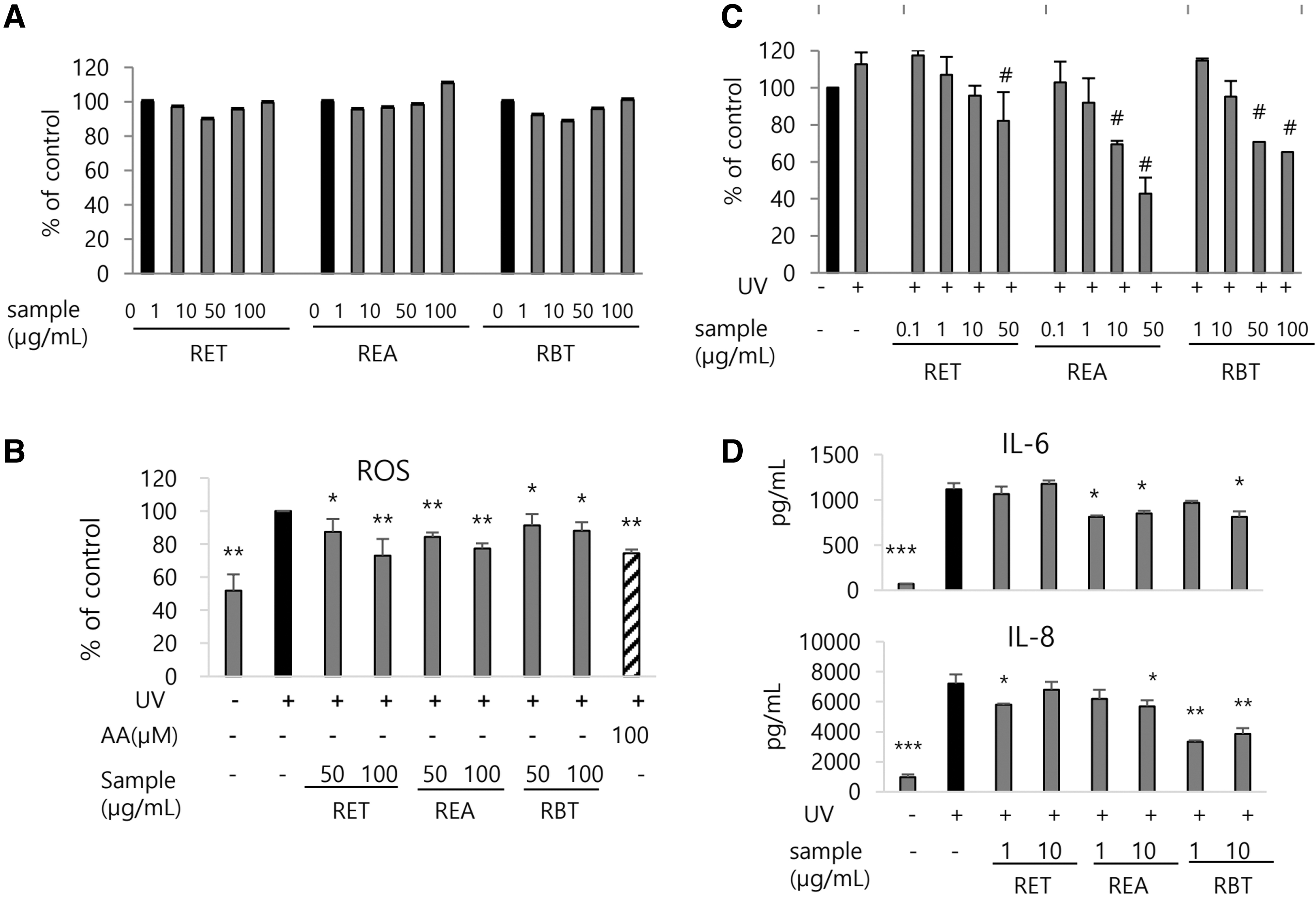
Inhibitory effects of RMF extracts on ROS and proinflammatory cytokine generation induced by UV irradiation in HaCaT cells.
The cell viability was maintained above 85% compared with that of normal control, when the cells were pretreated at lower concentration than 100 μg/mL of RET, 50 μg/mL of REA, and 100 μg/mL of RBT, respectively, before UV irradiation (Fig. 2C). Upon treatment of each sample, REA treatment (1 μg/mL) reduced the UV-induced IL-6 and IL-8 secretion by 23.9% and 21.1% (P < .05), respectively, whereas RBT treatment (10 μg/mL) significantly reduced it by 27.2% (P < .05) and 46.4% (P < .01), respectively. RET treatment (1 μg/mL) reduced only UV-induced IL-8 secretion by 19.7% (P < .05) (Fig. 3D).

Effects of RMF extracts on the expression of MMP-1 and type I procollagen in UV-irradiated HDF cells. Cells were treated with sample for 24 h and irradiated with UV (50 mJ/cm2).
MMP-1 and type I procollagen expressions in HFDs
After checking of the cytotoxicity caused by sample treatment or UV irradiation to HDFs (Fig. 3A), we examined the effects of RET, REA, and RBT on the expression of MMP-1 and type I procollagen. UV irradiation markedly upregulated MMP-1 expression (P < .001) (Fig. 3B, C), which was reduced by REA treatment by 28.7% at 1 μg/mL (P < .01) and 72.6% at 10 μg/mL (P < .001), and was reduced by RBT treatment by 29.6% at 1 μg/mL and 51.7% at 10 μg/mL (P < .001). RET treatment also reduced it by 27.0% at 10 μg/mL (P < .01) (Fig. 3B). Consistently, Western blot demonstrated that treatment with RET, REA, and RBT treatments at 10 μg/mL reduced UV-induced MMP-1 expression at protein level by 23.5% (P < .05), 41.4%, and 34.7% (P < .001), respectively (Fig. 3C).
Furthermore, UV irradiation significantly reduced type I procollagen expression at the mRNA and protein levels to 41.8% and 24.7% of normal control, respectively (P < .01). However, RET treatment restored it by 29.3% at 1 μg/mL and 53.9% at 10 μg/mL (P < .001), whereas REA treatment at 10 μg/mL or RBT treatment at 1 μg/mL recovered it by 35.7% (P < .01) and 20.8% (P < .05), respectively (Fig. 3B). Western blot demonstrated that treatment with RET, REA, or RBT at 10 μg/mL before UV irradiation significantly increased (P < .05) type I procollagen protein expression compared with vehicle treatment by 111.3% (P < .05), 348.6%, and 231.4% (P < .01), respectively (Fig. 3C).
Nuclear factor kappa B and mitogen-activated protein kinases signal pathways
The biochemical mechanisms related to MMP-1 downregulation by REA or RBT treatment in UV-irradiated HDFs were examined because REA and RBT suppressed MMP-1 expression more strongly than RET. As shown in Figure 4, UV irradiation increased phosphorylation of p65 and JNK, whereas it did not increase phosphorylation of ERK and p38. Treatment with REA or RBT at 10 μg/mL inhibited UV-induced p65 and JNK phosphorylation (P < .05). RBT treatment also reduced ERK phosphorylation compared with vehicle treatment (P < .05). On the contrary, REA and RBT treatments did not affect phosphorylation of p38.

Inhibitory effects of REA and RBT on NF-κB and MAPK signaling pathways activated by UV irradiation in HDFs. Cells were treated with sample for 24 h, irradiated with UV irradiation (50 mJ/cm2) and harvested after 30 min incubation.
Epidermal thickness and inflammatory response in the mouse skin
No significant differences were observed among groups in body weight gains of mice during the experimental periods (data not shown). Representative images captured from the H&E stained mouse skins in each group are shown in Figure 5A. The epidermal thickness in UVC group was increased to 297% of that in NC group (P < .01), which was reduced in RM group by 18% (P < .05) (Fig. 5B).

Inhibitory effects of oral treatment of RMF extracts on epidermal skin thickening and proinflammatory cytokines induced by UV irradiation in hairless mouse skin.
The concentrations of IL-1β, IL-6, and TNF-α in the mouse skin were increased in UVC group to 248%, 169% (P < .05), and 197% (P < .01) of that in NC group, respectively. However, the concentrations of IL-6 and TNF-α were reduced in the RM group compared with UVC group by 35.1% (P < .05) and 47.8% (P < .01), respectively, whereas TNF-α production was significantly reduced in the RET group by 48.6% (P < .01). IL-1β production was not affected by administration of any extracts (Fig. 5C).
MMP-13 and type I collagen expressions in the mouse skin
The mRNA expressions of MMP-13, functional substitute in the mouse skin for MMP-1 in human skin, were markedly elevated in UVC group to 222% (P < .01) and 125% (P < .01) of NC group, respectively. However, UV-induced MMP-13 expression was reduced in RET group by 66.5% (P < .01) at mRNA level and 10.7% (P < .05) at protein level, whereas it was reduced in RM group by 54.8% (P < .01) at mRNA level and 9.6% (P < .05) at protein level (Fig. 6A, B). Meanwhile, type I collagen mRNA expressions were not significantly changed by UV irradiation, but it was significantly higher in RET and RM groups than in UVC group by 223% and 230% (P < .05), respectively (Fig. 6A).
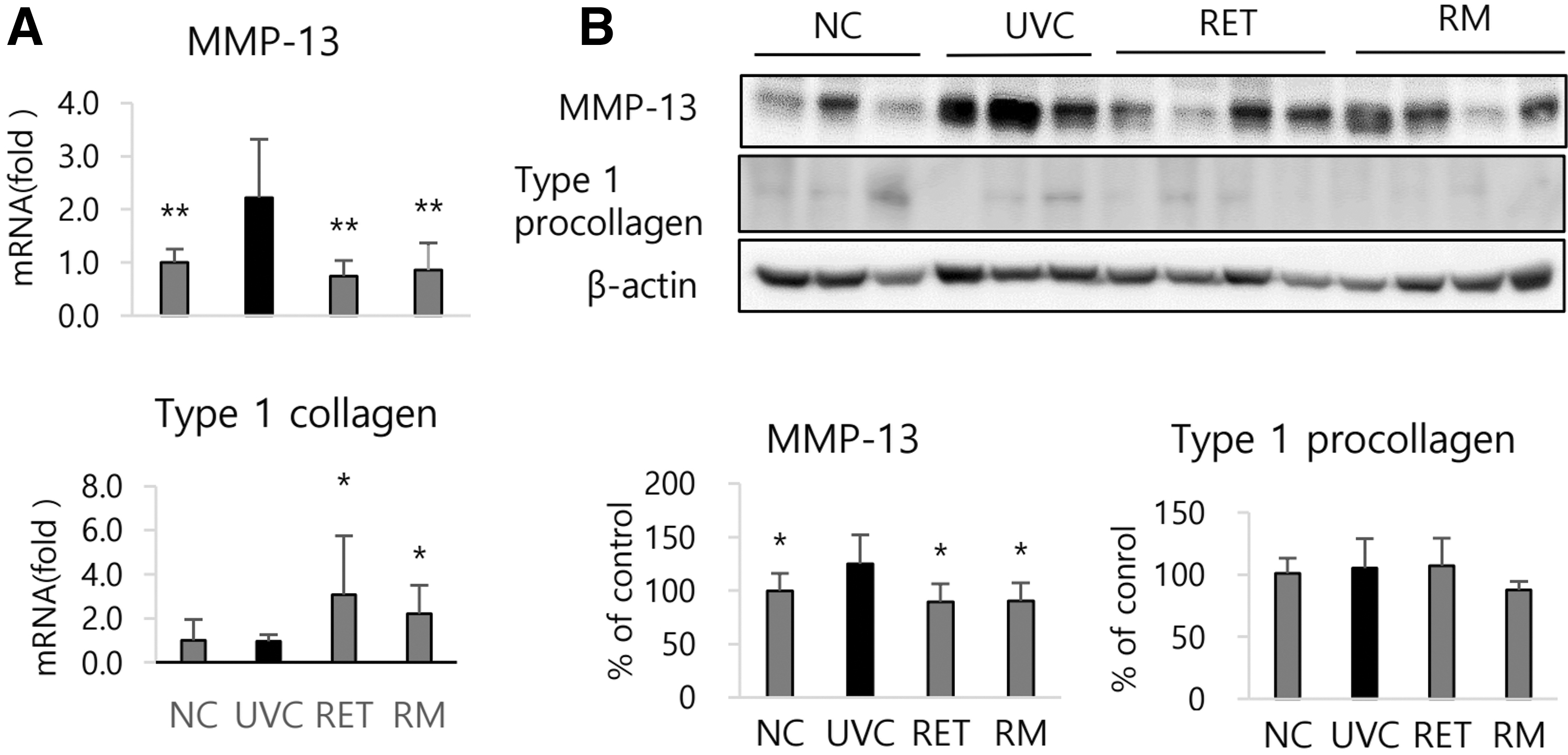
Effects of oral treatment of RMF extracts on MMP-13 and type I collagen expression modulated by UV irradiation in hairless mouse skin.
Phenolic compounds analysis
The HPLC chromatogram of each sample is shown in Figure 7. We identified eight compounds, including quercitrin (quercetin-3-O-rhamnoside, Fig. 8A), hyperin (quercetin-3-O-galactose), and isoquercetin (quercetin-3-O-glucoside) (Table 2). Two of them, quercitrin and hyperin, were common in RET, REA, and RBT, and five of them were common in REA and RBT.
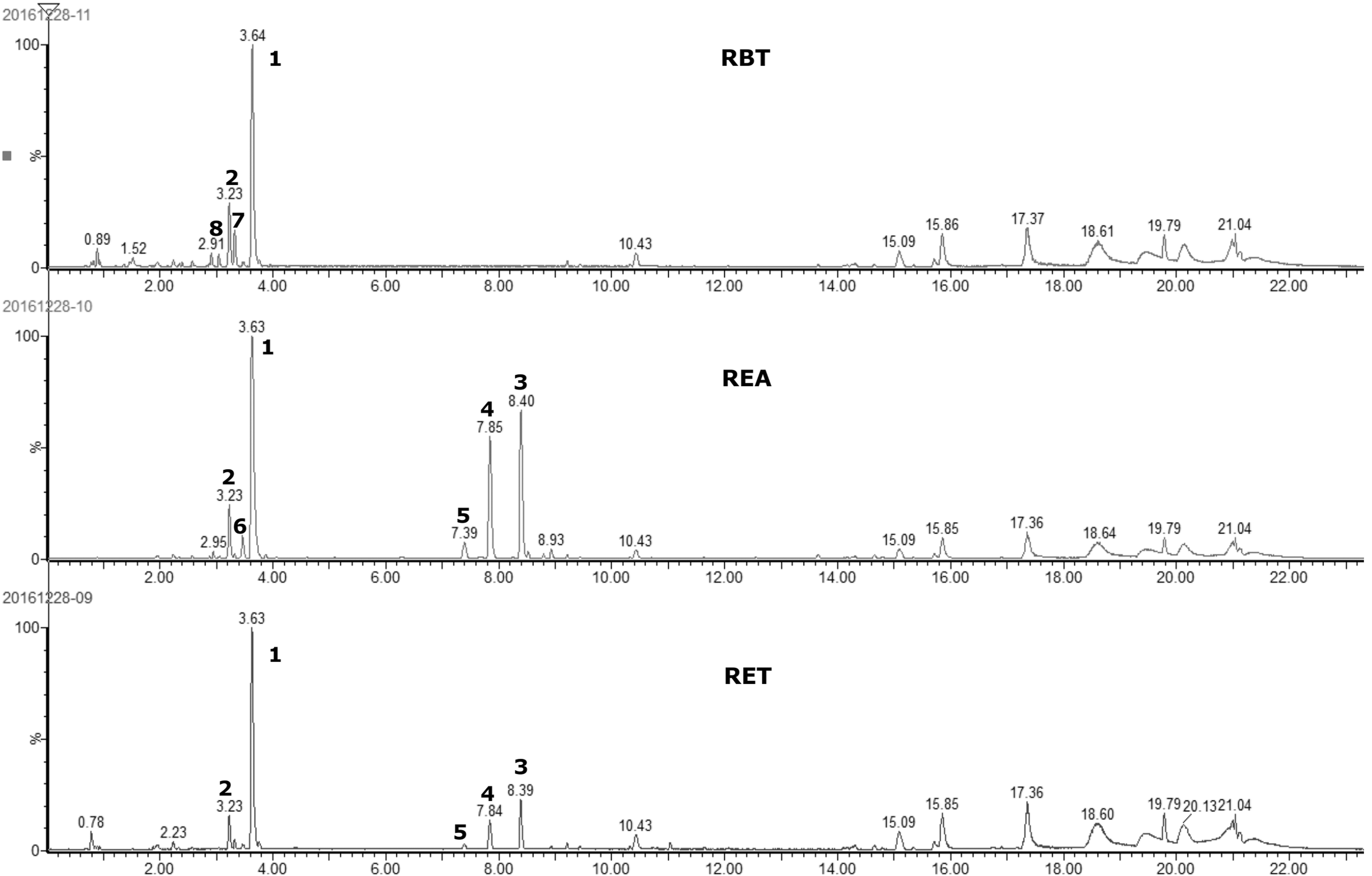
UHPLC chromatogram of main phenolic peaks in RMF extracts. UHPLC, ultra performance liquid chromatography; RBT, butanol fraction from RMF ethanol extract; REA, ethylacetate fraction from RMF ethanol extract; RET, RMF ethanol extract.

Effects of quercitrin on MMP-1 and type I procollagen expression modulated by UV irradiation in HDFs.
Main Phenolic Compounds Identified in Extract of Rosa multiflora Flowers
Furthermore, we examined if quercitrin could prevent UV-induced collagen degradation in HDFs, because quercitrin showed the highest peak in all the samples. Quercitrin treatment at 40 or 100 μM before UV irradiation significantly reduced MMP-1 protein levels in both cell lysate and medium (P < .001). Type I procollagen reduction caused by UV irradiation was also significantly attenuated by quercitrin treatment (P < .05) (Fig. 8B, C).
Discussion
Various phenolic compounds are known to protect cells from the oxidative damage caused by ROS. Epigallocatechin-3-gallate and chlorogenic acid from tea or coffee effectively suppressed production of ROS and proinflammatory cytokines and increased expression of MMPs caused by UV exposure in HDFs, whereas it increased synthesis of type I procollagen. 20,21 Patwardhen and Bhatt 22 reported that treatment with the ethylacetate subfraction from clove ethanol extract containing high level of flavonoids attenuated UVB-induced photodamage in HDFs and increased tolerance to oxidative stress. We reported recently that REA and RBT as well as RET contained high levels of phytochemicals and strong antioxidant and suppressed NO, IL-6, TNF-α, and PGE2 productions induced by lipopolysaccharide (LPS) treatment in macrophages. Consistently, treatment of HaCaT cells with REA and RBT in this study attenuated the increases of ROS, IL-6 and IL-8 caused by UV exposure. Since UV-induced oxidative stress is known to stimulate production of various proinflammatory mediators, 23 these inhibitory effects on IL-6 and IL-8 elevations are considered to be mediated by their antioxidant activity. Particularly, RBT treatment strongly reduced IL-8, which is known to attract neutrophils into the dermis in the acute inflammatory responses of UV-exposed skin. 24
Collagen, a main protein component of ECM in skin, provides resiliency and strength to skin. Accordingly, increased collagen destruction or/and decrease of collagen synthesis can lead to wrinkle formation. Collagen fibers in the human skin dermis are mainly composed with type I and III. 25 If collagens in ECM are degraded excessively by the increase of MMP-1 activity, it results in collagen loss and substantial inflammation. Thus, the balance between collagen degradation and synthesis is considered to be an important factor for maintaining the integrity of ECM in skin. 26 Many studies revealed that oxidative stress caused by UV exposure can mediate suppression of procollagen expression and induction of MMP expression. 27,28 Our results showing that treatment with RMF extracts attenuated upregulation of MMP expression and downregulation of type I procollagen expression caused by UV irradiation in both in vitro and in vivo models imply that these extracts can effectively inhibit UV-induced collagen degradation, and eventually protect skin from alteration of ECM integrity.
Regarding to the biochemical mechanisms of the photoaging process, UV-induced oxidative stress is known to modulate the activities of protein kinases through nuclear factor kappa B (NF-κB), mitogen-activated protein kinases (MAPK), and activated protein (AP)-1. 25,29,30 It was reported that antioxidant enzyme expression was reduced with the acceleration of oxidative damage and promoting of NF-kB and/or MAPK signaling in aged or photoaged skin cells. 31 Proanthocyanidins, a main phytochemical in grape seed, was reported to inhibit ROS generation and protein oxidation in UVB-exposed keratinocytes through suppression of phosphorylation of ERK, JNK, and p38, three major MAPK subfamilies in mammalian cells. 32,33 In UVB-exposed mouse skin, blackberry extract inhibited oxidative damage and inflammation through the modulation of NF-kB/p65 and MAPK pathways. 34 From the results in this study, we can suggest that REA and RBT reduced MMP expression in UV-irradiated skin cells through inactivation of NF-kB/p65, JNK and/or ERK, probably owing to the decrease in oxidative stress.
We intentionally exposed the cells with a mild dose of UV to avoid substantial cytotoxicity in this study. No obvious stimulation on ERK and p38 phosphorylation in UV-exposed cells might be because of the low UV strength. Afag et al. 32 reported that sublethal doses of UVB irradiation activated strongly JNK family of MAPK, but it activated ERK weakly. Farrukh et al. 35 demonstrated that UVB irradiation to human foreskin fibroblast cell line (Hs68) did not induce ERK phosphorylation. Son et al. 31 reported that increased ROS generation activated JNK pathway rather than ERK pathway.
In the animal experiment, the mixture of REA and RBT was prepared at a ratio of 1:1.5 corresponding to the extraction yield ratio because the yield of REA and RBT was so low and these two extracts showed similar potencies of antioxidative and anti-inflammatory activities in the in vitro study. 16 Repeated UV exposure to mice noticeably induced epidermal thickening, IL-6 and TNF-α productions, and MMP-13 expression in the skin; however, these changes were significantly attenuated in the RM group administered with REA and RBT mixture. It was reported that UV-induced proinflammatory cytokines downregulated procollagen expression and upregulated MMPs expression in skin, leading to degradation of dermal collagen and elastic fibers that induces wrinkle formation. 26,36
We first identified eight phenolic compounds, including quercetin glycosides, such as quercitrin hyperin isoquercetin, and multiflorin B in RMF extracts. Quercetin, one of the most common flavonoids in plant, is mainly present in the glycoside form, 37 and has excellent antioxidant, anti-inflammatory, and antiproliferative activities in vitro. 38 Lee et al. 39 reported that hyperin, isoquercitrin, and quercetin isolated from the root of Acanthopanax chiisanensis showed a potent anti-inflammatory activity through suppression of nitrite production in LPS-stimulated macrophages. Multiflorin B, one of the kaempferol glycoside derivatives, was reported to inhibit UVB-induced erythema formation. 40 A few studies revealed that quercitrin mitigated oxidative stress and inflammatory response in UV-irradiated skin cells 41 –43 ; however, there is no report on effects on UV-induced MMPs expression. We observed that quercitrin treatment significantly reduced MMP-1 levels and elevated type I procollagen levels in UV-irradiated HDFs in this study. Collectively, it could be suggested that quercitrin might be one of the effective components responsible for photopreventive activity of RMF extract. However, more studies are needed to ascertain those beneficial properties and underlying mechanisms and also test the toxicity of RMF extracts in various systems.
In conclusion, oral treatment with RET, REA, and RBT attenuated UV-induced skin photoaging in mice through suppression of ROS generation, proinflammatory cytokine production, and MMP expression. Quercitrin is suggested to be a potential effective compound to inhibit photoaging.
Footnotes
Acknowledgment
The authors thank the Ministry of Science, ICT and Future Planning, Republic of Korea.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This research was supported by the fund of this ministry through the National Research Foundation (NRF-2014R1A2A2A01-007435).