
Abstract
β-amyloid peptide (Aβ) has been considered a critical factor that is associated with the development of oxidative stress and neuroinflammation in the pathogenesis of Alzheimer's disease. This study was performed to evaluate the effect of geraniin on Aβ 25–35-caused oxidative damage and neuroinflammatory response, and its underlying mechanism. Geraniin protected pheochromocytoma12 (PC12) cells from Aβ 25–35-mediated cell death by reducing oxidative stress and restoring cell cycle dysregulation. Moreover, geraniin markedly attenuated Aβ-triggered DNA injury that was partially associated with decreases in caspase-3 activity. Moreover, the compound significantly downregulated the release of neuroinflammatory factors. Upregulation of nuclear factor-κB activity was suppressed by geraniin, which was due to suppression of JNK, ERK1/2, and the p38 mitogen-activated protein kinase (MAPK) pathway. This was the first study to support further understanding of geraniin as a promising agent against neurotoxicity in the reduction of oxidative stress and neuroinflammation.
Introduction
Alzheimer's disease (AD) is an irreversible neurodegenerative disorder marked by memory and cognitive failure. The hallmark pathologies of AD include extracellular plaques composed of β-amyloid peptide (Aβ) and intracellular neurofibrillary tangles (NFTs) of the hyperphosphorylated tau proteins. 1 Recently, it has become more obvious that the deposition of Aβ is an important initiating event of neuronal damage and cell death in AD. 2 Further, exposure of both rat and human neurons to Aβ fibrils showed an elevated level of tau protein phosphorylation, suggesting that the fibril formation might alter the state of tau phosphorylation that accelerates NFT formation. 3
Even though the etiology of AD has remained obscure, many studies have demonstrated that Aβ deposition triggers excessive accumulation of reactive oxygen species (ROS) in neurons. 4 Aβ-induced oxidative imbalance has been confirmed to increase the levels of byproducts associated with oxidized protein, DNA, RNA, and lipid. In contrast, decreased levels of antioxidants or related antioxidant enzymes have been found in patients with AD. 5 –7 Further, Aβ accumulation has been observed to affect both respiratory function and membrane potential in mitochondria that impair calcium homeostasis, which, in turn, leads to the opening of permeability transition pores and apoptosis. 8,9
Another source of Aβ-induced ROS generation involves activated microglia during neuroinflammatory responses to the senile plaque deposition. 10 Aβ-activated microglia release pro-inflammatory factors such as nitric oxide (NO), prostaglandin E2 (PGE2), and tumor necrosis factor alpha (TNF-α). Excessive production of these cytokines contributes to amyloidogenic process as well as neuronal damage, and ultimately cellular death. 11
Nuclear factor-κB (NF-κB) modulates the neuroinflammation process by targeting downstream genes such as cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS). The activated NF-κB by Aβ deposition triggers the transcription of pro-inflammatory mediators, including COX-2, iNOS, TNF-α, and etc. 12 Moreover, the NF-κB activation is also modulated by the mitogen-activated protein kinase (MAPK) family, including JNK, ERK, and p38. Therefore, discovering a multi-potent compound that simultaneously inhibits AD pathogenic pathways might lead to preventive strategies for AD.
Geraniin is predominantly found in traditional herbal plant families, including Geranium, Euphorbiaceae, and Sapindaceae. 13 Geraniin is known for its broad spectrum of multi-biological properties, including hepatoprotective, antioxidative, antidiabetic, anti-inflammatory, anticancer, antihypertensive, and antitumor effects. 14 –20 In our previous study, geraniin, isolated from Geranium thunbergii, selectively and specifically inhibited BACE1 with an IC50 value of 2.8 μM. 21 Further, geraniin markedly attenuated prolyl endopeptidase (IC50, 0.07 μM), which is involved in the degradation of neuronal cell adhesion molecules. 22 However, limited study of the neuroprotective effects of geraniin and its molecular mechanism has been undertaken until now. Therefore, this study demonstrated the specific underlying mechanisms of neuroprotection by geraniin on oxidative damage and neuroinflammation in Aβ 25–35-induced rat pheochromocytoma12 (PC12) cells.
Materials and Methods
Cell culture and Aβ preparation
PC12 cells were cultured in RPMI1640 supplemented with 10% horse serum, 5% FBS, and penicillin (100 U/mL). Cells were preserved at 37°C in a CO2 incubator before the experiments.
Aβ 25–35 dissolved in dimethyl sulfoxide (DMSO) was re-suspended in phosphate buffered saline (PBS). Before use, the solution was incubated at 37°C for 2 days to form amyloid fibrils.
Cell viability evaluation
PC12 cells (5 × 104 cells/mL in a 96-well plate) were maintained for 24 h. The cells were pretreated with geraniin at different concentrations (0.1, 1, and 10 μM) for 1 h before Aβ 25–35 treatment for 24 h. MTT solution was added to the cells at 37°C. After 3 h of incubation, the medium was removed. The formazan crystals in each well were solubilized in DMSO, and the absorbance was read by using a microplate spectrophotometer at 570 nm (ELX808; BioTek, Winooski, VT, USA).
ROS levels analysis
Cells were incubated with CM-H2DCFDA in the dark at 37°C for 30 min and re-suspended in HBSS. The cells were placed on glass slides and stored overnight. Images of ROS were acquired by using a fluorescence reader at excitation 485/emission 528 nm (FLX800; Winooski, VT, USA).
Flow cytometric quantification of apoptosis and cell cycle
Cells (1 × 106 cells/mL in a 24-well plate) were washed and re-suspended in PBS. For apoptosis analysis, the cell suspension was stained with Annexin V and Dead Cell Reagent; the percentage of apoptotic cells was calculated by using the Muse™ Cell Analyzer (Millipore, Billerica, MA, USA). For cell cycle assay, cell pellets were incubated with Muse cell cycle reagent and assayed by using the Muse Cell Analyzer.
Detection of apoptotic nuclei
Harvested cells were fixed by 4% paraformaldehyde at RT for 20 min. The attached cells were incubated with Hoechst 33342 for 15 min at RT in the dark. The apoptotic morphology was detected by using a fluorescence microscope at a magnification of × 400 (Olympus Optical Co., Tokyo, Japan).
Determination of NO and PGE2 levels
Cell culture media, after incubating the cell with sample and/or Aβ 25–35, were used to determine NO and PGE2 production. Equal volumes of the media and Griess reagent were incubated for 10 min at RT. The levels of NO were analyzed by using a microplate reader (ELX808) at 570 nm.
The cell supernatants were collected and mixed with PGE2-specific antibody for 2 h. After incubation, the excess reagents were removed and stop solution was added. Absorbance wavelength was monitored at 450 nm through ELX808 reader.
Western blot analysis
Western blotting was conducted as previously described. 23 Protein samples were incubated with primary antibodies against TNF-α, COX-2, iNOS, p-65, p-IκB-, p-JNK, p-p38, p-ERK, and β-actin, and they were then incubated with secondary antibodies. Protein bands were detected by using Atto EZ-capture (Tokyo, Japan).
Statistical analysis
All data were expressed as the means ± standard deviation of the results obtained from three independent experiments. Statistical significance was determined by using the Student's t-test at P ≤ .05.
Results
Geraniin attenuates Aβ 25–35-triggered neurotoxicity in PC12 cells
Treatment with geraniin alone did not affect cell survival (data not shown). Aβ 25–35 treatment reduced cell viability by 74.5% ± 1.2% compared with the control (100% ± 1.95%, Fig. 1A). In the group pretreated with geraniin, cell viability was increased in a concentration-dependent manner. Particularly, geraniin at 10 μM exhibited a complete amelioration of Aβ 25–35-induced cell death.
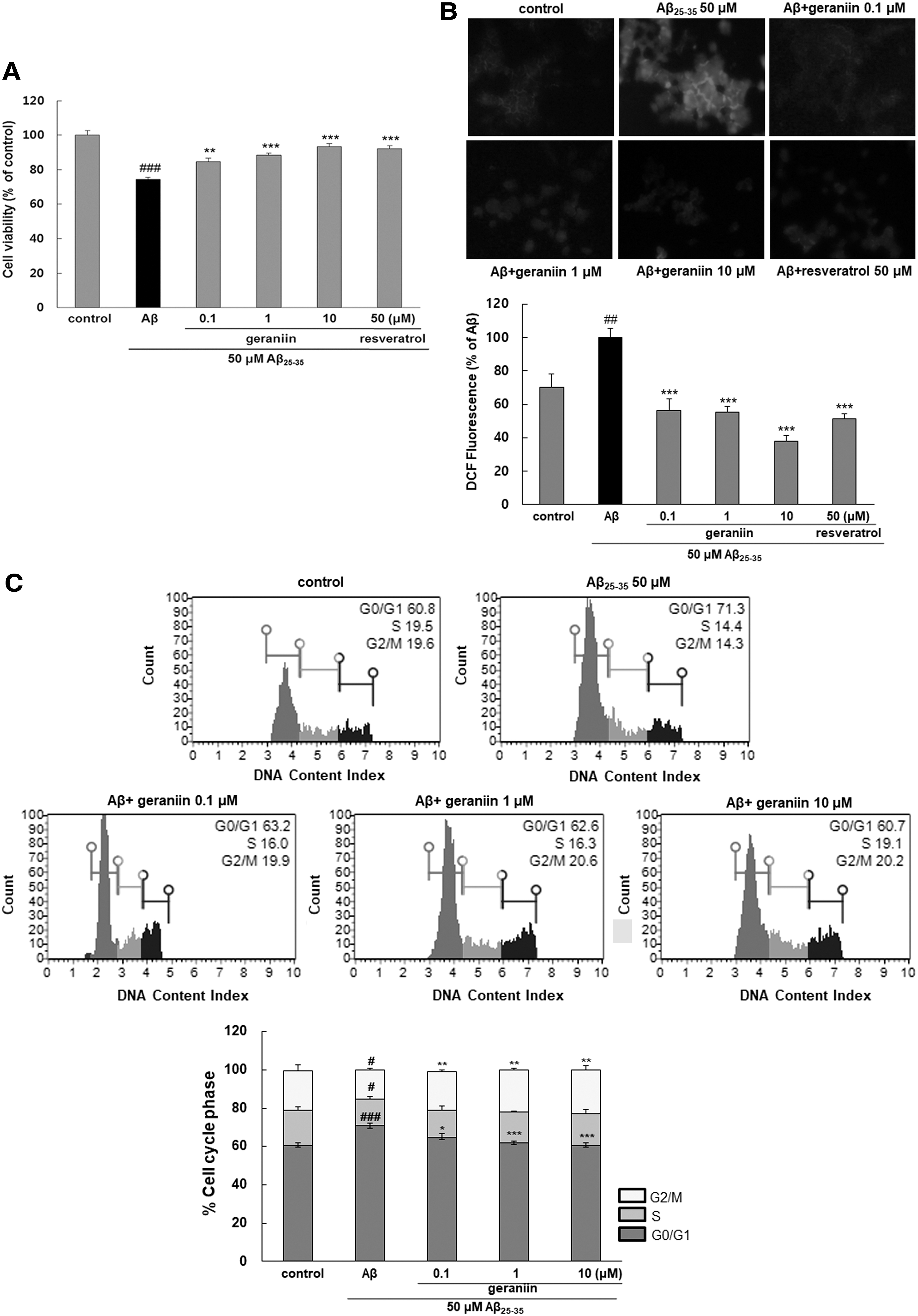
Neuroprotective effect of geraniin against Aβ
25–35-mediated cell death
Oxidative damage is a principal event during the development and progression of AD. 5 Aβ has been found to impair mitochondrial redox activity and induce ROS production in the early stage of AD, and this hyper-oxidative stress, consequently, led to senile plaque accumulation. A significant increase of ROS levels was observed when cells were treated with Aβ (Fig. 1B). Geraniin pretreatment significantly suppressed intracellular ROS levels compared with the control group.
To determine whether Aβ-stimulated neuronal damage was attributable to cell cycle arrest, the cells were analyzed by flow cytometry. Importantly, after Aβ 25–35 exposure, the proportion of cells in the G0/G1 phase increased from 58.7% ± 0.24% to 73.2% ± 0.42% and those in the S phase decreased from 15.75% ± 0.62% to 11.11% ± 0.99%, demonstrating that Aβ halted the cell cycle at the G0/G1 phase. However, the distorted cell cycle arrest was restored by pretreatment with geraniin in a dose-dependent manner (Fig. 1C).
Geraniin prevents cellular apoptosis induced by Aβ 25–35
Figure 2A showed that Aβ 25–35-triggered apoptosis marked by the characteristic morphological changes, including DNA fragmentation and chromatin condensation, was increased, whereas fewer apoptotic nuclei were observed in geraniin pretreated cells. For further quantitative evaluation of apoptotic cells, flow cytometry analysis was employed. Exposure to Aβ 25–35 significantly enhanced late apoptosis (22.75 ± 0.30, P < .001) in comparison with the control (7.75 ± 0.13), as shown in Figure 2B. However, geraniin markedly reduced the late stage of apoptosis in a dose-dependent mode.
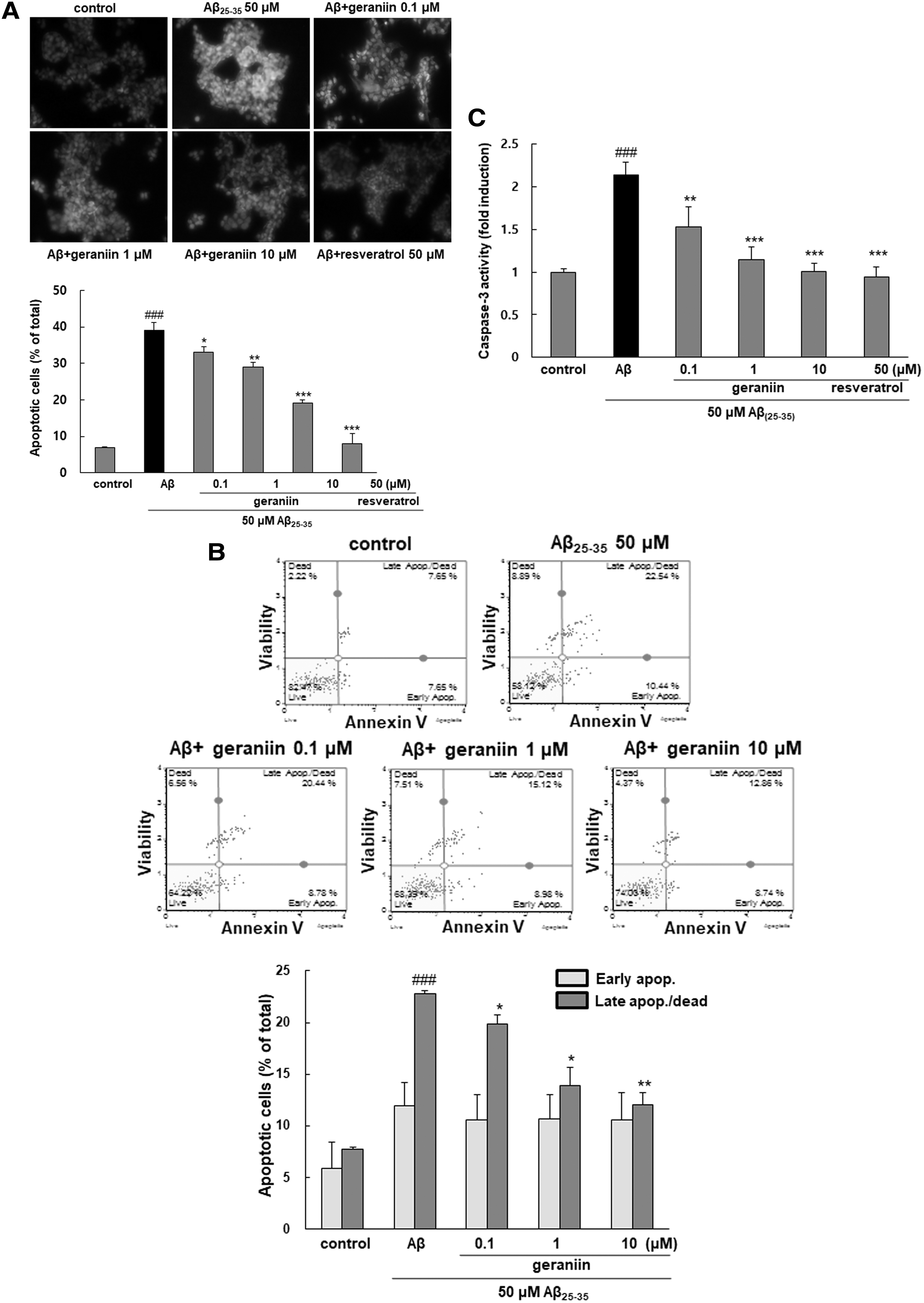
Protective effect of geraniin against Aβ
25–35-stimulated apoptosis and caspase-3 activation PC12 cells were pretreated with geraniin for 1 h before treatment with or without Aβ
25–35 (50 μM) for 24 h.
The activity of caspase-3, a terminal executor of apoptosis, was augmented by more than twofold after Aβ 25–35 incubation (P < .001, Fig. 2C). 24 However, when the cells were pre-incubated with geraniin, cell injury was noticeably ameliorated. Further, 10 μM geraniin exhibited a similar suppressive effect against apoptosis to that of resveratrol, a positive control.
Geraniin attenuates Aβ 25–35-induced expression of inflammatory mediators and their synthesis enzymes
As shown in Figure 3A–C, the production of NO, PGE2, and TNF-α was subsequently increased in response to Aβ 25–35 treatment. However, geraniin pretreatment significantly suppressed the expression of these inflammatory mediators. Particularly, geraniin at 10 μM completely blocked Aβ 25–35-stimulated NO production.
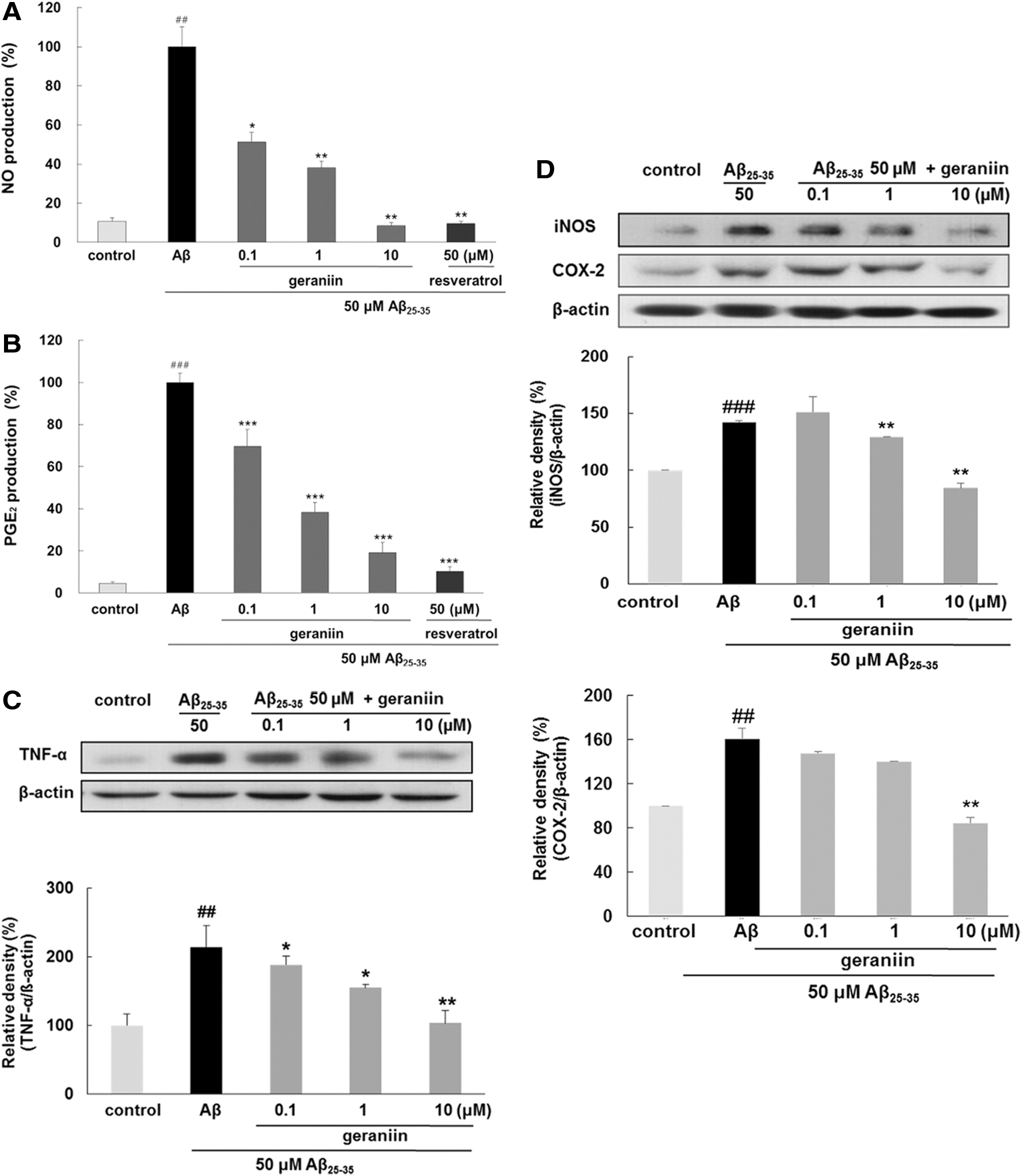
Inhibitory effects of geraniin on NO, PGE2, TNF-α, iNOS, and COX-2 expressions in Aβ
25–35-induced PC12 cells. The levels of
To determine the inhibitory effect of geraniin on regulating the expression of related synthesis enzymes, the levels of iNOS and COX-2 were evaluated (Fig. 3D). Consistent with its results on NO and PGE2 expression, geraniin suppressed the Aβ 25–35-induced protein expression of iNOS and COX-2 in a dose-dependent mode, implying that geraniin blocks the production of inflammatory mediators through decreasing the expression of iNOS and COX-2.
Geraniin suppresses the Aβ 25–35-stimulated NF-κB and MAPKs signaling pathway
NF-κB is an essential target for regulating inflammation, as it induces the transcription of many proinflammatory molecules. 11 As shown in Figure 4A, geraniin treatment inhibited phosphorylation of p65 subunit in Aβ 25–35-triggered PC12 cells in a dose-dependent mode. Interestingly, geraniin (0.1, 1 and 10 μM) treatment led to suppression of p65 phosphorylation to 87.69% ± 30.02% (P < .05), 71.09% ± 5.26% (P < .01), and 32.85% ± 8.67% (P < .01), respectively. An increased phospho-IκBα protein level by Aβ 25–35 was completely restored by geraniin even at 1 or 10 μM.
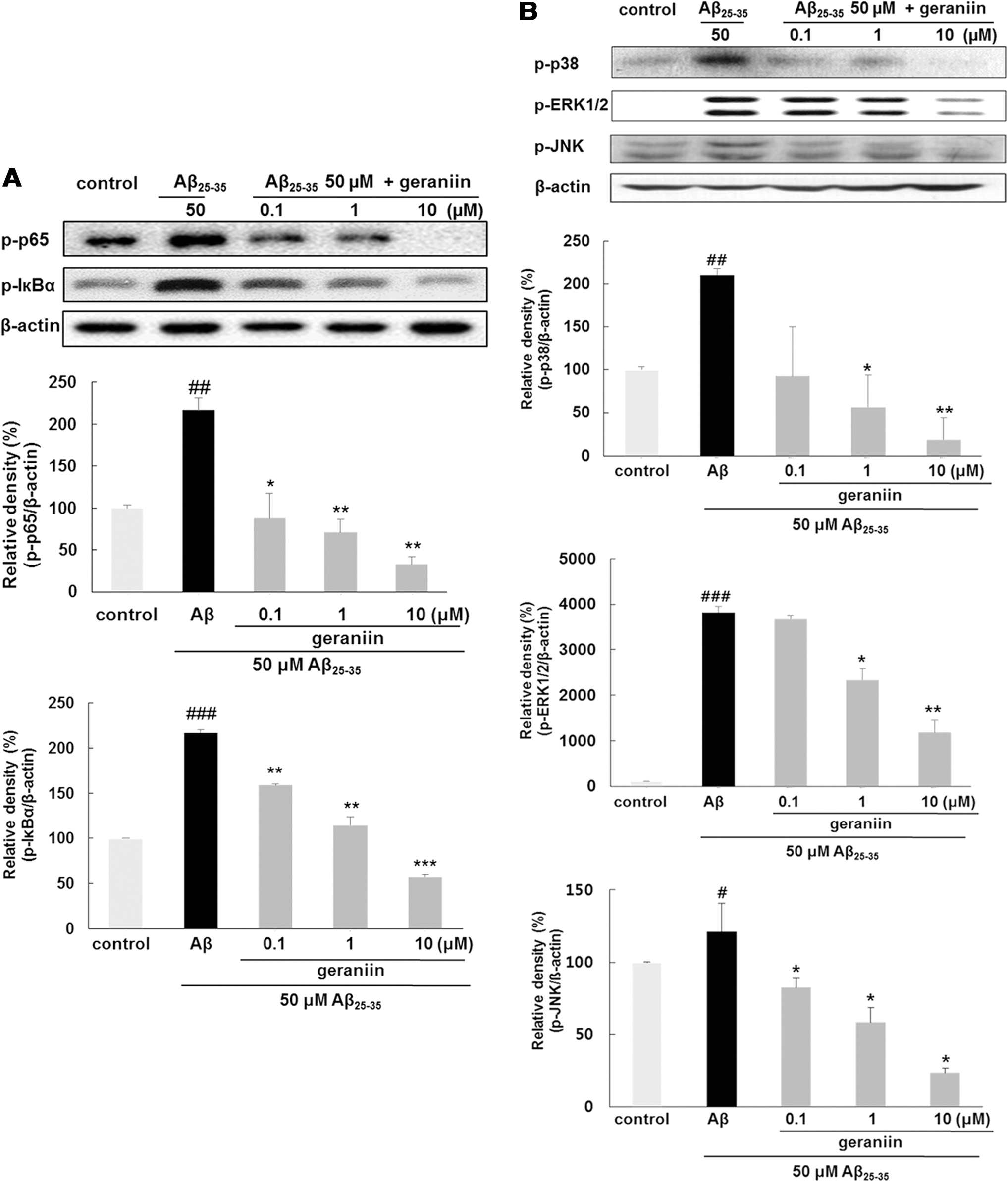
Inhibitory effects of geraniin on NF-κB and MAPKs activation in Aβ
25–35-stimulated PC12 cells
The levels of p38, ERK, and JNK MAPKs were elevated by Aβ 25–35 treatment, but they were markedly reduced by geraniin (Fig. 4B). In particular, the level of all MAPKs phosphorylation was statistically lower than that of the control at the highest concentration of geraniin.
Discussion
Many studies have suggested that Aβ mediates inflammatory responses, oxidative stress, mitochondrial membrane potential depolarization, tau phosphorylation, and Aβ overproduction. Consequently, given the complexity of the pathological pathways of AD, the therapeutic research should be focused on candidates that can modulate the multiple molecular targets involved in AD.
Since many studies have demonstrated that resveratrol possesses antioxidant, antiapoptotic, and anti-inflammatory effects in neuronal cell lines, 24 –27 the compound has been widely used as a positive control for in vitro neuronal damage studies. 28 –30 A growing body of evidence shows that resveratrol plays potential therapeutic roles in cancers, acute pancreatitis, respiratory diseases, and osteoarthritis by modulating NF-κB and MAPK signaling. 31 –34 In neurodegenerative disease, resveratrol suppressed oxidative stress and inflammation by Aβ through suppressing NF-κB activation. 35,36 Resveratrol, as an NF-κB signal transduction inhibitor, prevented the disruption of the blood brain barrier (BBB) and the overactivation of astrocyte and microglia. 37,38 The protective properties of resveratrol on the activation of NF-κB/MAPK pathway by Aβ were confirmed in our previous studies. 23,39
Geraniin has several hydroxyl groups, which provide antioxidant properties by scavenging free radicals and inactivating ROS. In this study, geraniin suppressed Aβ 25–35-dependent injury, to some extent, by decreasing oxidative stress as revealed by the suppressed levels of intracellular ROS, and it restored cell viability. In addition to eliminating oxidative stress directly, the antioxidant effect of geraniin may also be augmented via inducing signaling pathways that ultimately lead to the upregulation of antioxidant enzymes. 20,40 –42 A previous study has shown that geraniin reduced H2O2-caused oxidative injury in HepG2 cells via the translocation of nuclear factor-erythroid 2-related factor 2 (Nrf2), a crucial transcription factor that is responsible for regulating antioxidant and phase II detoxification genes. 40 The administration of geraniin (10 or 50 mg/kg b.w.) restored the levels of oxidative stress response factors, including serum antioxidants, glutathione (GSH) redox balance, and protein carbonyl contents in obese rats. 41 Further, the antioxidative ability of geraniin was demonstrated in tetrachloromethane-triggered acute hepatic dysfunction in mice and rats, as evidenced by a noticeable increase of GSH, superoxide dismutase, and catalase activity. 20,42
According to our results, Aβ 25–35 was markedly augmented late apoptosis. Aβ induced excessive ROS production, which accelerates mitochondrial dysfunction, release of cytochrome C (Cyt C), which initiates the activation of caspase-3 cascade leading to apoptosis. In addition, Aβ activates proapoptotic factors, including Bax and poly ADP ribose polymerase (PARP), and reduces antiapoptotic protein Bcl-2. 43 A previous study by Geske et al. demonstrated that the nuclear injury is too severe for reversibility and/or reparability in late apoptosis after caspase activation and DNA fragmentation has occurred. 44 However, the severity of late apoptosis was attenuated by the pretreatment of geraniin (Fig. 2B). Geraniin effectively inhibited apoptosis from being induced by proapoptotic stimuli. 45 –47 The compound protected pulmonary fibroblast cells against γ-radiation-evoked apoptosis and attenuated H2O2-induced apoptosis through decreasing the release of Cyt C, indicating recovery of mitochondrial function. 45,46 It also restored the levels of inactivated PARP and Bcl-2 in ethanol-induced apoptosis in mice liver cells. 47 Overall, this study confirmed that geraniin reduced excessive intracellular ROS levels and suppressed caspase-3 activation, thereby preventing Aβ 25–35-evoked apoptosis.
To delineate the mechanism associated with the anti-inflammatory activity of geraniin, its effect on MAPKs/NF-κB-mediated neuroinflammatory responses by Aβ 25–35 was studied. Our study demonstrated that geraniin blocked iNOS and COX-2 expression, which, in turn, led to the decreased levels of inflammatory mediators. Particularly, geraniin suppressed NO production more potently than TNF-α and PGE2. In addition, phosphorylation and degradation of IκBα required for NF-κB activation were inhibited by geraniin. Similar anti-inflammatory effects of geraniin on NF-κB activation have been reported in various cell types, including epithelial cells, macrophages, and RAW264.7 cells. 48,49 In addition, intraperitoneal injection of geraniin (20 mg/kg) reduced cisplatin-induced kidney injury via inhibiting NF-κB signaling pathways in mice. 50 Our results also revealed that geraniin dramatically ameliorated the level of MAPKs phosphorylation in Aβ 25–35-treated PC12 cells with a potent effect at 10 μM. Geraniin exhibited similar inhibitory activity on the MAPKs phosphorylation as several other existing MAPK inhibitors, such as LY 294002 (PI3K inhibitor), SB203580 (p38 inhibitor), and SP600125 (JNK inhibitor). 51 Taken together, our results show that geraniin has potential antioxidative, antiapoptotic, and anti-inflammatory properties in Aβ 25–35-triggered PC12 cells and may provide an attractive therapeutic and preventive strategy for AD.
Further, the safety of geraniin-enriched extract of Nephelium lappaceum (2000 mg/kg) was demonstrated in rats administered the extract for 4 weeks. There was no significant difference observed in mortality, side effects, and biochemical analysis of parameters, such as total protein, creatinine, serum urea, ALP, and AST. 52 Also, the supplementation of geraniin did not cause any toxicity in brain, kidney, and heart of the diet-induced obesity rat model. 41
Despite several studies showing various biological effects of geraniin, further and detailed studies are needed to investigate its bioavailability and permeability through the BBB. In addition, it is noteworthy that several hydrolyzable tannins, including corilagin and punicalagin with a similar chemical structure with geraniin, are absorbed and excreted in a small amount when orally consumed by rats. 53,54
In conclusion, our present study demonstrated the protective effect of geraniin against Aβ 25–35-stimulated PC12 cell damage for the first time. The neuroprotective activity of geraniin was induced by suppression of both oxidative stress and cellular apoptosis. In addition, geraniin exerted anti-inflammatory properties by upregulating ERK1/2, JNK, and P38 MAPKs, which resulted in the suppression of NF-κB and thus inhibited the expressions of inflammatory mediators. Therefore, the present results uncovered a basis for the neuroprotective property of geraniin and suggested the potential usage of geraniin in AD prevention.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by the National Research Foundation of Korea (NRF) grant funded by the Ministry of Education, Science, and Technology (NRF-2019R1A2C2006237).