
Abstract
The aim of this study was to investigate the efficacy of an ethanol extract of Physalis alkekengi (PA) and its mechanistic pathway of action at the molecular level for its antiobesity properties. Four-week old male Institute of Cancer Research (ICR) mice were acclimatized for a week before starting the high-fat diet (HFD) for 2 weeks to induce obesity, followed by 8 more weeks of oral administration of 10 mg/kg orlistat and 300 mg/kg of PA extract, along with HFD. Body weights of the mice and feed and water intake were recorded weekly. After a total of 12 weeks, mice were euthanized, and blood, liver, and adipose tissues were harvested for further analysis. Administration of PA extract inhibited the progression of obesity by reducing weight gain, weight of adipose tissue, and normalizing serum triglyceride, glucose, total cholesterol, high-density lipoprotein, low-density lipoprotein, alanine aminotransferase, and aspartate aminotransferase. PA extract prevented the progression of nonalcoholic steatohepatitis induced by HFD and prevented the enlargement of liver. Phosphorylation of adenosine monophosphate-activated protein kinase α increased while phosphorylation of acetyl-CoA carboxylase was reduced. The browning gene uncoupling protein 1 expression was also increased by PA extract treatment. Our findings revealed that the antiobesity properties of PA extract may be mediated by browning of white adipose tissue.
Introduction
Obesity is known to be the cause of many diseases, including cardiovascular diseases, nonalcoholic fatty liver disease (NAFLD), and also inflammation-related diseases. In obesity, adipocyte hypertrophy occurs. To ensure enough blood flow to adipocytes, inflammation takes place. This in turn induces the release of proinflammatory cytokines, also known as adipokines, such as tumor necrosis factor-α (TNF-α) and interleukin (IL)-6, which induce the occurrence of other ailments when released into the bloodstream. 1 Therefore, obesity should not merely be viewed as a metabolic disorder but also as an inducer of many other ailments.
Physalis alkekengi (PA), also known as Chinese lantern, is normally found in Southern Europe and Asia. It has a bright orange reddish paper-like cape housing its fruit. PA has been reported for anti-inflammatory effects, 2 antihyperglycemic activity, 3 and even antimicrobial activity. 4 The fruits of PA have been widely used in traditional Chinese medicine for treating cough, pharyngitis, eczema, and even jaundice. 5 However, there are no reports on the antiobesity effects of PA. Orlistat, which is a known pharmacological drug for treating obesity, acts as an inhibitor of pancreatic lipases directly acting through the gut. However, side effects, including abdominal pain, soft stools, and increased defecation, were observed in patients. 6 Therefore, it would be desirable to find natural products with fewer side effects that that could replace pharmacological drugs.
Brown adipose tissue (BAT) is known to induce thermogenesis, which is crucial for heat production. It is found abundantly in small mammals and newborns, which converts lipids from white adipose tissue (WAT) for producing heat. Therefore, increasing BAT thermogenesis is one of the therapeutic ways for curbing obesity, as increased energy expenditure will use up lipids in WAT. 7 Thermogenesis is regulated by mitochondrial uncoupling protein 1 (UCP-1). Hence, the upregulation of UCP-1 in WAT may be therapeutic in curbing obesity. UCP-1 is also regulated by peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α). Therefore, in this study, we investigate whether PA is able to counter obesity, and whether it is acting through the browning of WAT to form beige adipose tissue.
Our study revealed that PA extract has antiobesity properties by upregulation of phosphorylated-adenosine monophosphate-activated protein kinase α (p-AMPKα) and downregulating phosphorylated-acetyl-CoA carboxylase (p-ACC). PA has also suppressed fat synthesis-related genes such as fatty acid synthase (FAS), acetyl-CoA carboxylase (ACC) and sterol regulatory element-binding protein 1 (SREBP-1), and upregulated UCP-1 in WAT, revealing the possible mechanistic pathway of the antiobesity effects of PA.
Materials and Methods
Reagents
TRIzol reagent used was acquired from Invitrogen (Carlsbad, CA, USA). Antibodies for p-AMPKα, AMPKα, p-ACC, and β-Actin were from Cell Signaling Technology (Danvers, MA, USA). Primers for FAS, ACC, SREBP-1, UCP-1, PGC-1α, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were purchased from Bioneer (Daejeon, Republic of Korea).
Preparation of PA extract
Dried PA fruits were ground and extracted with 70% ethanol for 2 h at 80°C. The extracts were condensed using a rotary evaporator, frozen, and lyophilized to obtain the powder form of the extract. The extract was weighed accordingly for the respective experiments.
Gas chromatography–mass spectrometry analysis of PA extract
An Agilent 7890A GC (Agilent Technologies, Santa Clara, CA, USA) with a DB-5MS column with an i.d. of 30 m × 0.25 mm, and an Agilent 5975C mass selective detector were used to separate and quantify the constituents of PA extract. Samples were injected in split mode at a temperature of 250°C. The temperature of the transfer line and the ion source were 280°C and 230°C, respectively. The column temperature was held at an initial temperature of 70°C for 1 min, then raised to a temperature of 300°C for 30 min. Helium was the carrier gas, with a constant flow rate of 1 mL/min. Mass spectrometry was acquired using the electron ionization and scan modes.
High-fat diet induced obesity in Institute of Cancer Research mice
Four-week-old male Institute of Cancer Research (ICR) mice were purchased from Orient Bio (Gyeonggi-do, Republic of Korea). All experiments were conducted with the approval of the Institutional Animal Care Committee of Kyungpook National University (approval no. KNU 2018-57). Animals were housed in a 12-h light/dark controlled room with a regulated temperature of 22 ± 2°C and a relative humidity of 50 ± 10%. Mice were given chow and water ad libitum. The mice were acclimatized for 1 week before conducting experiments. Mice were then separated into four treatment groups of six mice each: normal chow, high-fat diet (HFD), HFD with orlistat (10 mg/kg), and HFD with PA extract (300 mg/kg). After 1 week of acclimatization, other than the mice in normal chow group, mice were given HFD (D12492; Research Diets, New Brunswick, NJ, USA). After 2 weeks of HFD, samples of orlistat and PA extract were given orally daily for 8 more weeks. The weight of the mouse, along with feed and water intake were measured weekly. Twenty-four hours after the last dose of orlistat or PA extract, mice were euthanized by collecting blood through cardiac puncture. Serum was separated through centrifugation from the collected blood. The kidney, liver, and spleen, along with the retroperitoneal adipose tissue, subcutaneous adipose tissue, epididymal adipose tissue, and mesenteric adipose tissue were also collected, weighed, frozen, or fixed in neutral buffered formalin for further analysis. The feed efficiency ratio was calculated by the ratio of feed intake per mouse per day against the weight gain per day.
Real-time polymerase chain reaction of epididymal adipose tissue of mice
Epididymal adipose tissue harvested from mice was homogenized using a hand grinder with TRIzol solution to extract the RNA. BAT was harvested from mice in the normal group as a positive control. RNA was separated using chloroform, then purified using alcohol. Concentration of RNA was determined using a nanophotometer and was reverse transcribed and the resultant cDNA was used for real-time polymerase chain reaction (PCR) against related genes using a CFX96 Real-Time system (Bio-Rad, Hercules, CA, USA). The resultant product was loaded on to ethidium bromide-stained agarose gel for confirmation. The sequences of the primers used are shown in Table 1.
Primer Sequences for Real-Time Polymerase Chain Reaction
ACC, acetyl-CoA carboxylase; FAS, fatty acid synthase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; PGC-1α, proliferator-activated receptor gamma coactivator 1-alpha; SREBP-1, sterol regulatory element-binding protein 1; UCP-1, uncoupling protein 1.
Western blot analysis of liver tissue
Proteins from the liver tissue were extracted using Pro-Prep solution (Invitrogen, Daejeon, Republic of Korea). Concentrations of extracted proteins were measured using Pro-Measure solution (Invitrogen). Separation of proteins was carried out using 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis, followed by a transfer onto polyvinylidene fluoride membranes. Membranes were blocked with 5% skim milk for 1 h at room temperature, incubated with primary antibody at a concentration of 1:3000 overnight, followed by secondary antibody incubation (1:1000). The membranes were washed with 1% Tween-20 tris-buffered saline buffer before developing the membranes in an ECL chemiluminescence gel developer (General Electric, Boston, MA, USA). Western blotting was carried out in triplicate and quantified using ImageJ software (NIH, USA).
Biochemical markers in the serum of mice
Blood was collected through cardiac puncture and left to stand for 2 h in a vacutainer (BD, Plymouth, UK). Tubes were then centrifuged at 503 g for 15 min. Separated serum was collected and stored at −70°C until analysis. Serum triglyceride, glucose, total cholesterol, low-density lipoprotein (LDL), high-density lipoprotein (HDL), alanine aminotransferase (ALT), aspartate aminotransferase (AST), creatinine (CRE), and blood urea nitrogen (BUN) levels were analyzed using a blood analyzer.
Hematoxylin and eosin staining
Epididymal adipose tissue and liver tissue were harvested. After weighing, the tissues were directly fixed in neutral buffered formalin before dehydration and fixation in paraffin blocks. The fixed tissue was then sectioned into 5 μm sections using a microtome. Sections were then stained with hematoxylin and eosin (H&E) before visualization under a microscope. The number of adipocytes per frame was calculated using AdipoCount software. 8 Thirty frames were analyzed per group and averaged. The frames were observed at 100 × under a microscope.
Statistical analysis
GraphPad Prism version 7.00 (San Diego, CA, USA) was used to analyze the statistical significance using one-way analysis of variance (ANOVA) with Dunnett's posttest. Data are presented as mean ± standard deviation, and P < .05 was considered significant.
Results
PA extract reduces weight gain induced by HFD
HFD administration induced a significant change in weight compared with the control group at week 4. However, both orlistat and PA extract suppressed the weight gain in mice, which was significant from week 5 onward, confirming the antiobesity effects of PA extract (Fig. 1). There was also no significant difference in feed in water intakes among the groups. However, there was a significant increase in the food efficiency ratio (FER) in HFD-group, and the significant decrease in both orlistat and PA extract groups signify the ability of PA extract to reduce the efficiency of the conversion of feed to body weight. No significant changes in feed and water intake among the groups were observed (Table 3).
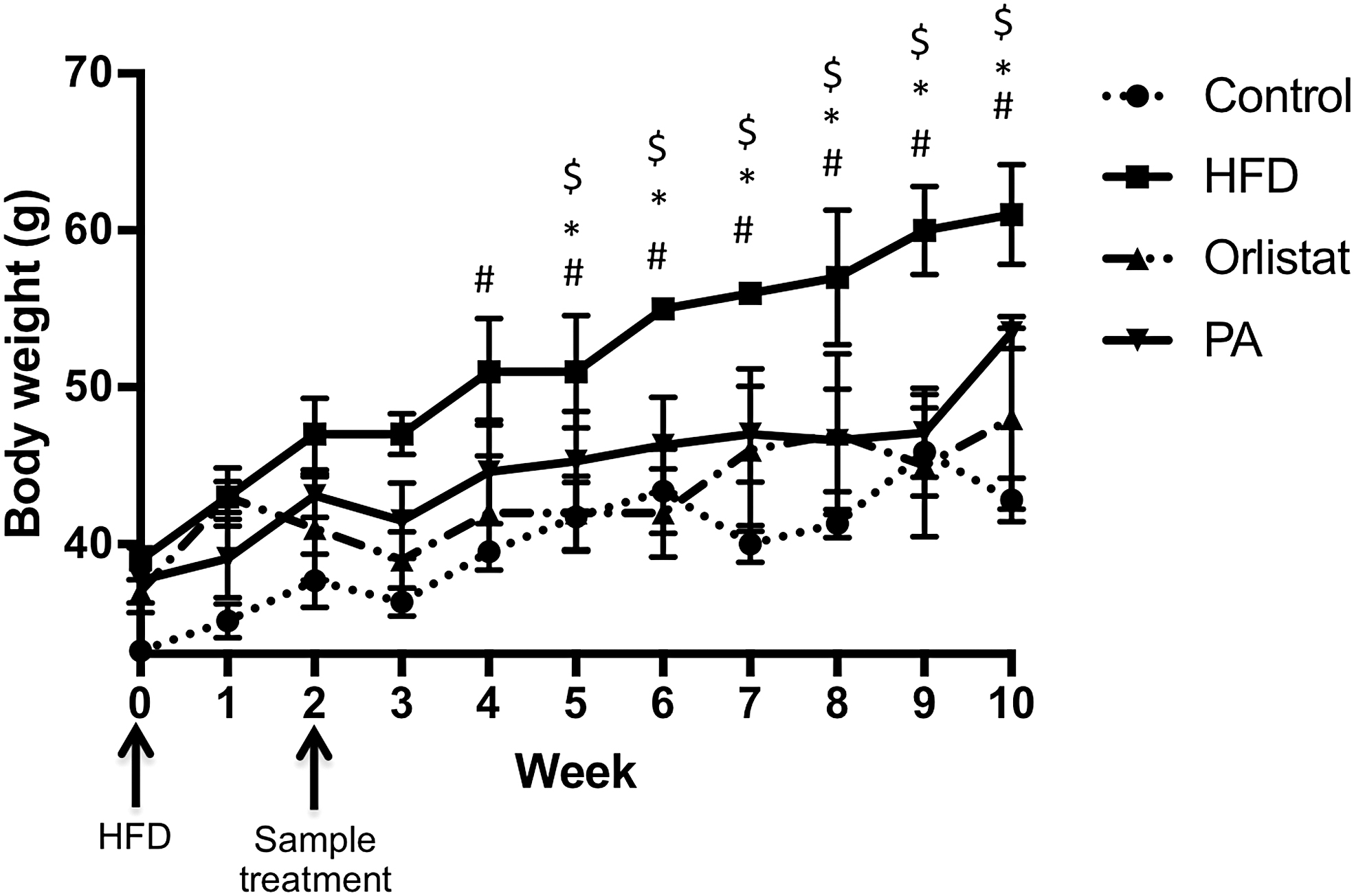
Mice were fed with or without HFD for 8 weeks with or without PA or orlistat treatment after treated with or without HFD 2 weeks prior. Weight of mice in control, HFD-, orlistat-, and PA-treated groups were measured weekly. #A significant difference of HFD-treated group at P < .05 compared with control group; *a significance for PA at P < .05 compared with HFD group; whereas $a significance for orlistat at P < .05 compared with HFD group. Data were analyzed using one-way ANOVA with Dunnett's posttest. Data are presented in mean ± SD (n = 6). ANOVA, analysis of variance; HFD, high-fat diet; PA, Physalis alkekengi; SD, standard deviation.
Feed Intake, Water Intake, and Feed Efficiency Ratio of Mice in Each Group
The average feed intake and water intake per mice per day, and the FER for control, HFD, orlistat and PA-treated groups. Data were analyzed using one-way ANOVA with Dunnett's posttest. Data were presented in mean ± SD (n = 6).
A significant difference at P < .05 compared with control group; *a significant difference at P < .05 compared with HFD group.
ANOVA, analysis of variance; FER, feed efficiency ratio; HFD, high-fat diet; PA, Physalis alkekengi; SD, standard deviation.
PA extract reduced weight of retroperitoneal, subcutaneous, and mesenteric adipose tissue in mice
Weights of different tissues were recorded, and from our findings, epididymal, retroperitoneal, subcutaneous, and mesenteric adipose tissues were significantly increased in HFD-group compared with the control group. The PA extract-treated group had significantly lower weights of retroperitoneal, subcutaneous, and mesenteric adipose tissue, whereas orlistat only reduced the weight of subcutaneous and mesenteric adipose tissue. The weight of spleen and kidney was not changed among the groups. However, the weight of the liver increased significantly with HFD treatment but was reduced significantly with treatment of orlistat and PA (Table 4).
Weight of the Respective Adipose Tissues and Organs in Mice
The weight of epididymal, retroperitoneal, subcutaneous, mesenteric adipose tissue, spleen kidney, and liver of mice from control, HFD, orlistat, and PA-treated groups were harvested and weighed. Data were analyzed using one-way ANOVA with Dunnett's posttest. Data are presented as mean ± SD (n = 6).
A significant difference at P < .05 compared with control group; *a significant difference at P < .05 compared with HFD group.
PA increased the expressions of UCP-1 and PGC-1α, improved metabolic markers p-AMPKα and p-ACC, and reduced fat synthesis-related gene expressions
The mRNA expression of browning markers, UCP-1 and PGC-1α, in the epididymal adipose tissue was investigated. The BAT in normal mice exhibited high expressions of both UCP-1 and PGC-1α. Orlistat and PA groups both showed an increased expression of UCP-1. However, it was not significant for orlistat. There was no change in expression of PGC-1α with the treatment of orlistat and PA extract (Fig. 2A). The quantified gel images were averaged as shown (Fig. 2B). Protein expression of p-AMPKα was decreased by HFD and increased significantly with orlistat and PA extract treatment. There was an increase in expression of p-ACC in the HFD group, and it was reduced by both orlistat and PA extract. This reveals that PA extract might be acting through p-AMPKα and p-ACC, with browning of WAT to curb obesity (Fig. 2C). The gel images were quantified using ImageJ and are presented in Figure 2D. Gene expressions of FAS, ACC, and SREBP-1 in the epididymal adipose tissue of mice were investigated using real-time PCR as shown (Fig. 2D–G). Our results reveal that HFD increased the expressions of all genes associated with the synthesis of fat, but their expressions were significantly suppressed with PA and orlistat treatments. This further confirms the efficacy of PA in inhibiting obesity.
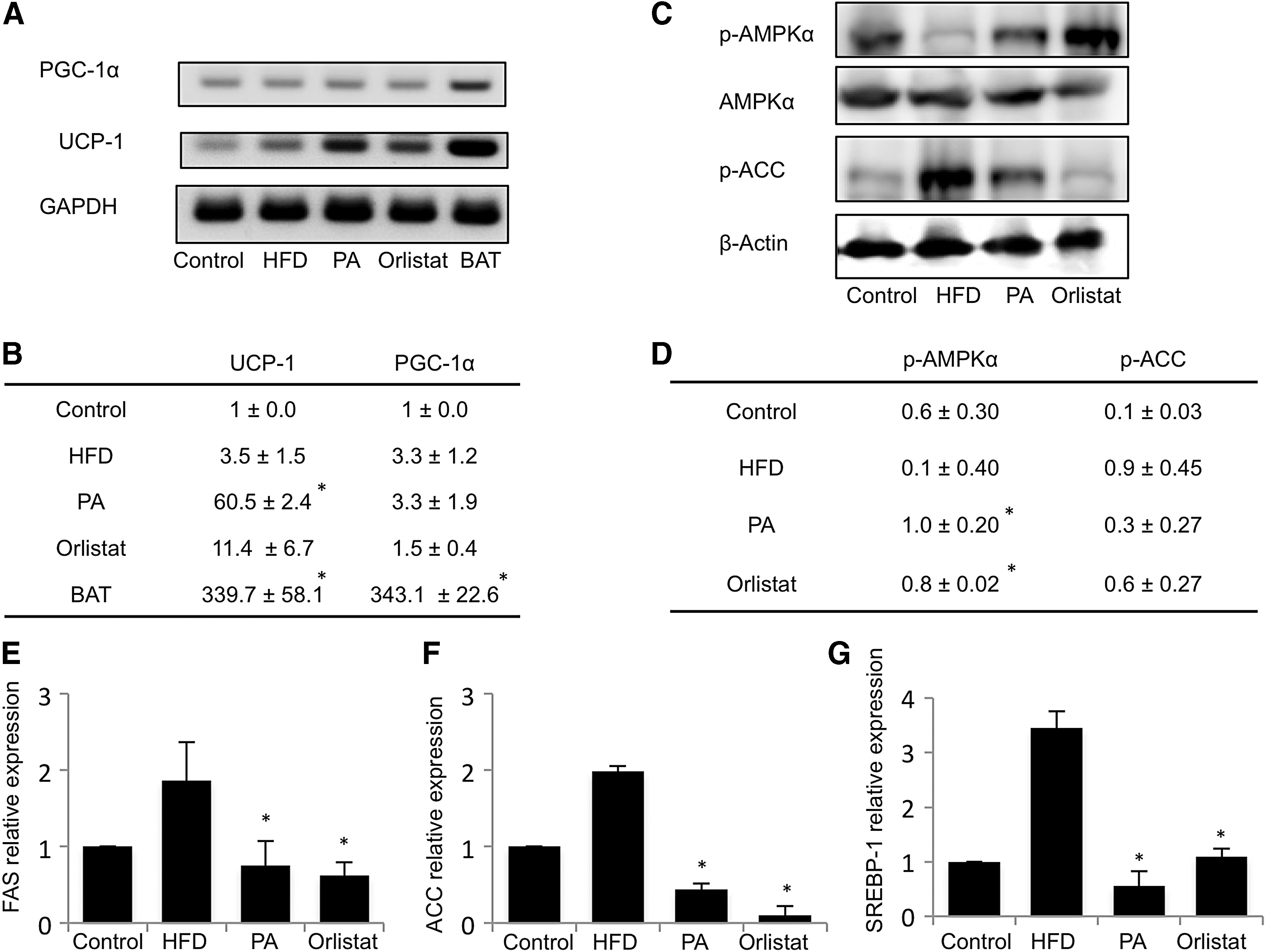
PA increased the expression of UCP-1 and regulated the expressions of p-AMPKα and p-ACC. RNA was extracted from epididymal adipose tissue and investigated for the expression of PGC-1α and UCP-1 using real-time PCR
PA extract improved serum triglyceride, glucose, total cholesterol, LDL, HDL, ALT, and AST levels in mice with HFD-induced obesity
The serum of mice were analyzed for triglyceride, glucose, total cholesterol, LDL, HDL, ALT, AST, CRE, and BUN levels. Our findings revealed that HFD significantly increased serum triglyceride, glucose, total cholesterol, HDL, and AST levels in mice. With treatment of orlistat, serum triglyceride, glucose, LDL, and AST levels were significantly decreased compared with the HFD-treated group, whereas treatment of PA extract has reduced serum triglyceride, glucose, total cholesterol, LDL, HDL, ALT, and AST levels compared with HFD-treated mice. This indicates that PA extract is more effective than orlistat for regulating HFD-induced obesity (Table 5).
Serum Biochemical Markers in the Serum of Mice
The serum biochemical markers in HFD-mice model. Data were analyzed using one-way ANOVA with Dunnett's posttest. Data are presented in mean ± SD (n = 6).
A significant difference at P < .05 compared with control group; *a significant difference at P < .05 compared with HFD group.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; BUN, blood urea nitrogen; CRE, creatinine; HDL, high-density lipoprotein; LDL, low-density lipoprotein.
PA extract improved histopathological appearance of liver and epididymal adipose tissue
Sizes of mice increased drastically after HFD treatment, but were reversed by PA extract and orlistat treatment (Fig. 3A). The coloration of the liver appeared paler, and there was a visible increase in size compared with the control group. This was reversed with treatment of PA extract and orlistat (Fig. 3B). The size of adipocytes were markedly increased in HFD-treated mice, compared to the mice in control group, and was reduced with treatment of PA extract and orlistat. The number of adipocytes per frame also showed a significant decrease in HFD-treated group, whereas the number had been significantly increased in both orlistat- and PA extract-treated mice, as quantified using AdipoCount software (Fig. 3C). The decreased number of adipocytes per frame indicates the increased size of adipocytes. The H&E staining of liver sections revealed the ballooning of hepatocytes (as shown with arrowheads) and the infiltration of inflammatory cells (shown with arrows). The ballooning of hepatocytes, revealing a pale cytoplasm, and its foamy characteristics are signs of steatohepatitis. However, this characteristic can still be observed slightly in orlistat-treated groups, and was improved in PA extract-treated mice (Fig. 3D). This indicates the potency of PA extract to inhibit the progression of HFD-induced steatohepatitis and adipocyte hypertrophy.

The representative images of mice and liver for each group were represented
Discussion
Naturally sourced antiobesity supplements should be developed as pharmacological drugs such as orlistat brings unwanted side effects to the patient. PA has been consumed for its many beneficial effects in traditional medicine and reportedly has many beneficial properties. Previous literature has reported the antifertility properties of PA, 9 anti-inflammatory properties, 10,11 and even anticancer properties. 12 Other than that, reports have also revealed the antihyperglycemic activity of PA. 3,13 Some studies have also reported its antidiabetic activities. 14,15 Gas chromatography–mass spectrometry analysis has identified the major compounds of PA extract to be octanoic acid, 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl-, n-hexadecanoic acid, 5-hydroxymethylfural, and oleic acid (Table 2).
Compounds of Physalis alkekengi Identified Using Gas Chromatography–Mass Spectrometry Analysis
4H-Pyran-4-one and 2,3-dihydro-3,5-dihydroxy-6-methyl- have proapoptotic effects in human colon cancer cells, 16 whereas oleic acid has been even reported to prevent cardiovascular diseases. 17 However, the major constituents of PA extract have not been reported for antiobesity effects, revealing the antiobesity properties of the major constituents of PA extract. In our study, we have selected orlistat as the positive control as it is a widely used drug to treat obesity by inhibiting the absorption of fat through the inhibition of gastrointestinal lipase. 6 Comparing the efficacy and mechanism of action of PA to a widely used drug would further confirm the potency of PA.
Intake of a diet high in fats causes NAFLD, which has also been observed in HFD-fed mice exhibiting enlarged size of the liver accompanied by pale coloration. Histological analysis of liver has also revealed the ballooning of hepatocytes with a clear cytoplasm, indicating mild nonalcoholic steatohepatitis 18 as shown in Figure 3. Treatment with PA extract significantly lowered ALT and AST levels compared with the HFD group, which is comparatively more potent than orlistat, which only lowered AST levels. As both ALT and AST are liver damage markers, our findings further confirms the efficacy of PA extract to reverse HFD-induced liver damage.
As adipocytes secrete adipokines such as IL-6 and TNF-α, it has been reported that both these adipokines promote liver inflammation and tumorigenesis, 19 revealing a possible mechanism of action of PA extract; PA reduces WAT hypertrophy, reducing the secretion of adipokines, and prevented HFD diet-induced liver damage. This sheds light on the possibility that PA may have also prevented the secretion of adipokines as part of its mechanism of action in preventing HFD-induced liver damage.
AMPK regulates cell lipid metabolism through the phosphorylation of ACC, which suppresses fatty acid synthesis and at the same time, promotes fatty acid oxidation via the suppression of carnitine palmitoyltransferase 1 (CPT1). 20 Our study showed that PA extract regulates the increase of p-AMPKα and inhibits the phosphorylation of ACC, revealing the mechanistic pathway of action of PA extract. The increase of UCP-1 induced by PA extract revealed that PA extract induced the browning of WAT, however, UCP-1 was not activated by PGC-1α as there was no change in its expression. Further studies should be conducted to further elucidate the other mechanisms of UCP-1 activation. The significant decrease in weight of most adipose tissues in PA extract-treated mice further confirms that the decrease in weight may be due to the increased energy expenditure by increased thermogenesis due to the activation of mitochondrial UCP-1, which uses and depletes lipids stored in adipose tissue. Therefore, our study sheds light on the mechanism of action of PA extract in curbing obesity by increasing energy expenditure and also preventing the secretion of adipokines in adipocytes that prevented liver inflammation.
Our study revealed that PA extract efficiently inhibited weight gain in mice fed with HFD. Moreover, no significant change in feed and water intake indicated that PA extract and orlistat both reduced the conversion of feed into body weight, which is confirmed by FER. PA extract has also effectively reduced serum triglyceride and glucose levels, and reduced liver injury markers, ALT and AST.
Further research should be conducted to further elucidate the activation pathway of UCP-1. PA extract increases UCP-1 expression in WAT, reducing the amount of lipids in WAT and prevented WAT hypertrophy. Hence, lesser amounts of adipokines were secreted by WAT and liver damage was prevented. PA also significantly reduced fat synthesis-associated genes FAS, ACC, and SREBP-1 mRNA expressions in epididymal WAT. Our study reveals the antiobesity properties of PA extract and its potential to be developed into a plant-based antiobesity supplement. Single compounds found in PA like 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl-, n-hexadecanoic acid, 5-hydroxymethylfural, and oleic acid should also be studied separately to identify the main compound exerting antiobesity properties.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the National Research Foundation funded by the Korean government (grant no. 2018R1D1A1A09083797).